内隐多效性选择的ERP证据*
2018-05-11蔡荣华钟毅平张文洁
蔡荣华,钟毅平,范 伟,2,张文洁
(1.湖南师范大学认知与人类行为湖南省重点实验室,长沙 410082;2.中国科学院心理研究所,脑与认知科学国家重点实验室,北京 100101)
1 引言
有研究表明人们所作出的选择可能被两种性质的目标驱动,一种是有意识的外显目标,另一种则是不被意识到的内隐目标(Bagozzi & Dholakia,1999)。而在日常生活中,人们的选择常常是被自己能意识到的外显目标所引导(Bagozzi & Dholakia,1999;Simonson,2005)。也有研究表明,多种类型的目标构造了认知结构,这些认知结构受一些敏感的环境刺激影响,并在个体没有意识到的情况下,发挥它们的影响(Dijksterhuis,Smith,Van Baaren,& Wigboldus,2005;Fishbach,Friedman,& Kruglanski,2003;Morsella,Bargh,& Gollwitzer,2008;Shah & Kruglanski,2002,2003)。无意识目标的激活已被证明影响了个体的认知、行为和情感(Fitzsimons & Bargh,2003;Kruglanski et al.,2002)。还有学者的研究直接表明个体的选择可能是外显目标和内隐目标共同作用,两种目标都可能影响个体的选择,即使在个体存在明确的外显目标的情况下,内隐目标同样可能影响个体的选择(Chun & Kruglanski,2005;Simonson,2005),这就是内隐多效性选择。具体来说,内隐多效性选择是指选择的目标可以带来多种效果,能同时满足意识到的外显需要和未意识到的内隐需要(蔡荣华,钟毅平,2014)。个体的外显目标和内隐的目标会被同时激活,在其选择过程中,特定的目标结构会影响着这一激活过程(蔡荣华,钟毅平,2014)。
而选择行为被认为是决策的一个部分。决策的基本结构根据时间和功能的不同,可粗略划分为三个阶段。一是评估选项的效用并做出选择。 二是执行上一阶段的选择,在该过程中,需要完成一系列与执行有关的加工过程,如选择不同行动的先后顺序、禁止其他的竞争行为及选取适宜的行动时间。三是体验决策的结果,体验主要源自预期和实际结果之间的差异,这种差异对调整下轮决策选项的赋值具有重要意义(方平,陈满琪,姜媛,2009)。
前人的研究表明,与内隐多效性选择有关系的ERPs成分包括N2和P3等。回顾前人的研究发现,P3主要与背景更新假说,刺激的评定假说和反应的选择、执行有关(Gray,Ambady,Lowenthal,& Deldin,2004;张凤华,王金娥,张庆林,2007)。例如,有研究表明在作为情境下,“好”决策诱发了更大的P3正波,而在不作为情境下,并没有显示出这种涨与跌的差异(周海波,钟毅平,2011)。Torres等人的研究表明,病态赌博成瘾者在决策任务中比控制组被试诱发了更小的N2,而海洛因成瘾者在决策任务重诱发的N2波幅与病态赌博成瘾者、控制组都没有差异(Torres et al.,2013)。Filevich等人的研究表明,相比外部命令,被试面对瘙痒刺激时,主动决策不收回手时所诱发的晚期正成分(P3)更大一些(Filevich,Kühn,& Haggard,2012)。Cui等人的研究表明,在爱荷华赌博任务中,被试做盈利选择时在左脑会诱发更大的P3,被试做损失选择时在右脑会诱发更大的P3(Cui et al.,2013)。Zhang等人基于PCA的ERP朔源分析结果提供了关于当前结果对随后决策影响的神经基础的很有价值的信息。最为重要的是,该分析揭示了在PCA-P3时间窗口内右侧额极皮层(布鲁德曼10区)上的显著激活,它表明该区域很有可能是P3波形产生的生理机制(Zhang et al.,2013)。研究者预测符合多效性原则的选择行为应该是一个好的决策行为,它会比其他选择行为诱发更大差异的N2和P3波幅。
因此,基于以上的证据和理由,未来可以考虑从认知神经层面上展开内隐多效性选择的研究,进一步揭示内隐多效性选择的本质以及神经基础。
2 方法
2.1 实验目的与实验假设
实验目的:采用颜色选择实验,探讨内隐多效性选择及其神经机制。
实验假设:实验假设对被试实施积极的内隐启动后,被试的选择都遵循多效性效应。对比控制刺激,被试对积极启动刺激做多效性选择时会诱发更大的P2和P3波幅和更小的N2波幅。
2.2 被试
筛选70名大学生参加对颜色与国家相关程度的排序评定,评定的任务是对“红、橙、黄、绿、青、蓝、紫”七种颜色进行排序,与国家相关程度最高的排在最左边,与国家相关程度最低的排在最右边,从左至右相关程度依次递减。结果表明有84.29%的被试把红色排在了最左边,认为红色与国家的相关程度最高。31位将红色排在最左边的被试被邀请参加脑电实验。
实验收录31名在校大学生的脑电数据,这31位为将红色排在最左边的被试,均为右利手,身体健康无神经系统疾病,没有脑部损伤史,视力正常或校正后正常。实验完成后付给一定报酬。
2.3 实验材料
通过Photoshop软件制作“红、橙、黄、绿、青、蓝、紫”七种颜色的图片,图片大小和明暗度、饱和度控制为相同。进行实验时,红色图片与其他图片配对呈现作为目标刺激,共计6种呈现方式,在实验中每种显现方式重复15次,平衡左右位置后,如“红、橙”,“橙、红”,共计呈现180次。除红色以外的其他六种颜色自由组合配对呈现作为掩饰刺激,共计15种呈现方式,在实验中每种显现方式重复6次,平衡左右位置,如“蓝、橙”,“橙、蓝”等,共计呈现180次。
2.4 实验设计
实验为单因素两水平(图片类型:积极启动刺激、对照刺激)的被试内设计,因变量为行为数据(反应时)和ERP数据(N2、P2、P3波幅)。
2.5 实验程序
实验开始前先对被试进行积极认同启动,启动方法与以前的研究相同(蔡荣华,钟毅平,2014)。要求被试参加一个对国家认同度的调查,然后进行9点量表评分。完成调查后,要求被试参加正式实验。
正式实验时,首先在屏幕中央呈现一个300ms的十字架,紧接着随机呈现800~1200ms的黑屏,之后呈现500ms的配对纯色图片,最后再呈现1000ms的黑屏。在呈现配对的纯色图片刺激时,被试的任务是认真看刺激并进行分类按键反应,且要求又快又准。如果被试认为左边的图片颜色更纯一些,则按“1”键;如果被试认为右边的图片颜色更纯一些,则按“2”键;练习实验结束后,进行360trails的正式实验。

图1 实验2中一个试次的流程图
2.6 脑电记录
使用NeuroScan ERP记录与分析系统,按国际10~20系统扩展的64导电极帽记录EEG。在线纪录时以左侧乳突连线为参考电极,离线后转为双侧乳突为参考电极,离线双眼外侧安置电极记录水平眼电(HEOG),左眼上下安置电极记录垂直眼电(VEOG)。滤波带通为0.05~40Hz,采样频率为500Hz/导,头皮阻抗<5kΩ。
2.7 ERP数据处理与统计
完成连续记录EEG后,离线(off-line)处理数据,用NeuroScan软件校正VEOG,并充分排除其他伪迹。对不同相关程度的自我参照刺激呈现后的脑电数据进行分析与EEG叠加。波幅大于±80μV 者视为伪迹而自动剔除。分析时程(epoch)为刺激后1000ms,基线为刺激呈现前200ms。根据已有研究结果与本研究目的,对ERP数据(P2、N2、P3、LPP平均波幅)进行统计分析。
实验在枕叶区位置的电极(P1,PZ,P2,PO3,POZ,PO4)发现了明显的N1(120~200ms)、P2(200~280ms)、N2(280~360ms)、和P3(360~460ms)波,因此进行两因素重复测量方差分析,2(图片类型:积极启动刺激、对照刺激)×3脑区(左:P1,PO3;中:PZ,POZ;右:P2,PO4)。方差分析的P值采用 Greenhouse-Geisser法校正。将不同分类的总平均ERPs导入具有标准MRI头像的Curry6.0系统,用sLORETA方法在三壳球模型中重建各任务在不同时段的头皮活动源。
3 结果
3.1 行为结果
在积极认同评定中五个问题的平均得分分别为8.69、8.23、8.34、8.34、8.31,结果表明积极认同启动效果较好。在反应时上,重复测量方差分析表明图片类型存在着一个显著的主效应,F(1,30)=38.05,p<0.001,被试对积极启动刺激的反应时显著的快于对照刺激。
由于实验任务的设置,没有涉及正确率的考察。
3.2 ERPs结果
见图2,在两类刺激中都发现了N1,P2,N2和P3成分。重复测量方差分析表明,N1(120~200ms)在平均波幅上都没有发现显著的差异。
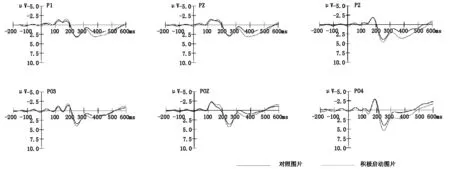
图2 两种图片在P1、PZ、P2、PO3、POZ和PO4电极点上诱发的ERPs总平均图
3.2.1 P2(200~280ms)

3.2.2 N2(280~360ms)

3.2.3 P3(360~460ms)
3.3 溯源分析结果
将不同分类的总平均ERPs导入具有标准MRI头像的Curry6.0系统,用sLORETA方法在三壳球模型中重建各任务在不同时段的头皮活动源。根据实验2的ERPs成分,将积极启动刺激减去对照刺激所得差异波取P300(360~460ms)成分的数据进行定位分析。结果发现积极启动刺激减去对照刺激所得差异波P300成分显著激活了颞中回(见图3)。
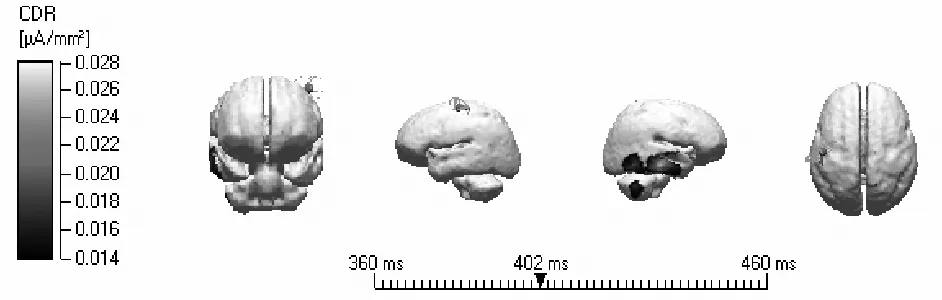
图3 积极启动刺激减去对照刺激所得差异波P300的溯源图
4 讨论
在当前的研究中,研究结果发现两类刺激在N1的波幅上均没有发现显著的差异,这可能是由于对刺激类型的尺寸大小、明暗度和复杂性等物理属性上做到了均衡处理。在这一时间阶段积极启动也没有影响到被试对图片刺激的加工,估计这种影响可能会出现在稍晚一点的阶段。
在刺激呈现后,研究结果发现在枕叶区电极观察到了明显的P2成分,对积极启动刺激的加工诱发了更大的P2波幅。有研究表明,P2成分可能是对刺激特征的快速侦查,易敏感的刺激会得更多的关注(Karayanidis & Michie,1996;Thorpe,Fize,& Marlot,1996)。此外,还有研究表明与情绪有关的刺激可能比中性刺激诱发更大的P2波幅,生物重要性更强的刺激容易得到注意(Carretié,Mercado,Tapia,& Hinojosa,2001)。但是这种注意缺乏高级的认知加工和控制资源的分配(Del Cul,Baillet,& Dehaene,2007;Hu,Wu,& Fu,2011)。因此,积极启动刺激加工得到的P2波幅更大,可能是由于对被试积极启动后,积极启动刺激的呈现会诱发被试的正性情绪,从而吸引了大量的注意资源。而人脑对控制图片的加工消耗的认知资源较小,所以诱发的P2波幅也更小。这一阶段发现差异可能与多效性选择行为无关,这是因为人脑对多效性刺激的选择与决策是晚期加工,可能发生在稍晚的阶段。
在枕叶区电极还发现了比较明显的N2成分,积极启动刺激比控制刺激诱发了更小的N2波幅。N2通常被认为是一种非特殊的成分,它与注意的转换机制相联系,随后一般会有P3成分出现(Kiehl,Laurens,Duty,Forster,& Liddle,2001)。在被动接受刺激的情况下,只有刺激足够突出或者发生了注意的转换,N2b/P3a都会被观察到(Näätänen,Simpson,& Loveless,1982)。近期,还有研究发现,N2成分可能与自我加工有关(Fan et al.,2013;Fan et al.,2011;钟毅平等,2014)。因此,在当前的研究中,积极启动刺激为红色图片,由于其突出的生物和社会意义上的重要性很快抓住了大脑的注意力。另外一种解释是,积极启动刺激为红色图片,而红色图片与国旗、国徽的主体颜色相同,有可能在某种程度上会诱发被试的集体自我或者心理自我,激发被试的归属感和依恋之情,进而诱发了更小的N2波幅。另外,还有研究表明,病态赌博成瘾者在决策任务中比控制组被试诱发了更小的N2,而海洛因成瘾者在决策任务中诱发的N2波幅与病态赌博成瘾者、控制组都没有差异(Torres et al.,2013)。因此,这一阶段发现差异也可能与多效性选择行为有关,人脑对多效性刺激的选择与决策有可能在这一阶段就开始了。
两种图片都能观察到具有显著差异的P3成分。P3通常被认为是人脑在晚期阶段对中枢控制加工的朝向反应(Campanella et al.,2002;Carretié,Hinojosa,Martín-Loeches,Mercado,& Tapia,2004;Yuan,Yang,Meng,Yu,& Li,2008)。而且前人的研究也发现,P3成分主要与背景更新假说,刺激的评定假说和反应的选择、执行有关(张凤华等,2007)。例如,有研究表明在作为情境下,“好”决策诱发了更大的P3正波,而在不作为情境下,并没有显示出这种涨与跌的差异(周海波,钟毅平,2011)。Filevich等人的研究表明,相比外部命令,被试面对瘙痒刺激时,主动决策不收回手时所诱发的晚期正成分(LPP)更大一些(Filevich et al.,2012)。Cui等人的研究表明,在爱荷华赌博任务中,被试做盈利选择时在左脑会诱发更大的P3,被试做损失选择时在右脑会诱发更大的P3(Cui et al.,2013)。对比控制刺激,人脑在加工积极启动刺激时会投入更多的认知和控制加工资源。因此,在实验中积极启动刺激比控制刺激诱发了更大的P3波幅。这些结果可能表明了积极启动刺激成功诱发了多效性选择行为,对积极启动刺激多效性选择的执行诱发了更大的P3波幅。这些结果还表明P3成分,可能不同于P2和N2成分,它可能反应了内隐多效性选择的精细加工过程,能够作为内隐多效性选择的有效的ERPs指标。
有趣的是,在P2和P3时间段,实验结果都发现了明显的内隐多效性选择效应,积极启动刺激比控制刺激诱发了更大的P2和P3波幅,而这一效应在右后部脑区最为明显(right sites)。而且偶极子溯源结果表明积极启动刺激减去对照刺激所得差异波P300成分显著激活了颞中回。这些实验结果与前人的研究相似,前人的研究发现ACC,VMPFC,眶额皮层(OFC)等都可能是个体决策的神经机制(Bechara,Damasio,& Damasio,2003;Kenning & Plassmann,2005)。这种右后部脑区的优势效应可能表明了右后部脑区是加工多效性刺激的核心和关键区域。这也为进一步的研究提供了一个方向,未来的研究可以通过具有精细定位功能的fMRI技术来探讨这一部分脑区是不是内隐多效性选择加工的关键的神经机制。
总的来说,当前的研究借助高时间进程的精密仪器ERPs,不但重复了内隐多效性选择,而且拓展了前人的研究,揭示了多效性选择行为的神经机制。研究结果表明P3成分,可能不同于P2和N2成分,它可能反应了内隐多效性选择的精细加工过程,能够作为内隐多效性选择的有效的ERPs指标。而且右后部脑区的优势效应可能表明了右后部脑区是加工多效性刺激的核心和关键区域。
蔡荣华,钟毅平.(2014).外显与内隐选择的多效性原则.心理科学,37(1),66-72.
方平,陈满琪,姜媛.(2009).决策的脑认知神经机制.心理科学,32(3),640-642.
张凤华,王金娥,张庆林.(2007).事件相关电位 P300 与蓝斑-去甲肾上腺素交感神经系统的研究与进展.中国组织工程研究与临床康复,11(5),965-967.
钟毅平,范伟,蔡荣华,谭千保,肖丽辉,占友龙,秦敏辉.(2014).正性情绪诱导下的自我参照加工:来自ERPS的证据.心理学报,46(3),341-352.
周海波,钟毅平.(2011).行为决策中的 “作为” 与 “不作为”:事件相关电位研究.中国临床心理学杂志,19(3),296-298.
Bagozzi,R.P.,& Dholakia,U.(1999).Goal setting and goal striving in consumer behavior.TheJournalofMarketing,63(1),19-32.
Bechara,A.,Damasio,H.,& Damasio,A.R.(2003).Role of the Amygdala in Decision‐Making.AnnalsoftheNewYorkAcademyofSciences,985(1),356-369.
Campanella,S.,Gaspard,C.,Debatisse,D.,Bruyer,R.,Crommelinck,M.,& Guerit,J.(2002).Discrimination of emotional facial expressions in a visual oddball task:An ERP study.BiologicalPsychology,59(3),171-186.
Carretié,L.,Hinojosa,J.A.,Martín-Loeches,M.,Mercado,F.,& Tapia,M.(2004).Automatic attention to emotional stimuli:Neural correlates.HumanBrainMapping,22(4),290-299.
Carretié,L.,Mercado,F.,Tapia,M.,& Hinojosa,J.A.(2001).Emotion,attention and the “negativity bias”,studied through event-related potentials.InternationalJournalofPsychophysiology,41(57),75-85.
Chun,W.Y.,& Kruglanski,A.W.(2005).Consumption as a multiple-goal pursuit without awareness.ApplyingSocialCognitiontoConsumer-focusedStrategy,201-217.
Cui,Q.,Vanman,E.J.,Wei,D.,Yang,W.,Jia,L.,& Zhang,Q.(2013).Detection of deception based on fMRI activation patterns underlying the production of a deceptive response and receiving feedback about the success of the deception after a mock murder crime.SocCognAffectNeurosci,9(10),1472-1480.doi:10.1093/scan/nst134
Del Cul,A.,Baillet,S.,& Dehaene,S.(2007).Brain dynamics underlying the nonlinear threshold for access to consciousness.PLoSBiology,5(10),2408-2423.
Dijksterhuis,A.,Smith,P.K.,Van Baaren,R.B.,& Wigboldus,D.H.J.(2005).The unconscious consumer:Effects of environment on consumer behavior.JournalofConsumerPsychology,15(3),193-202.
Fan,W.,Chen,J.,Wang,X.,Cai,R.,Tan,Q.,Chen,Y.,et al.(2013).Electrophysiological Correlation of the Degree of Self-Reference Effect.PLoSONE,8(12),e80289.
Fan,W.,Zhang,Y.,Wang,X.,Wang,X.,Zhang,X.,& Zhong,Y.(2011).The temporal features of self-referential processing evoked by national flag.NeurosciLett,505(3),233-237.
Filevich,E.,Kühn,S.,& Haggard,P.(2012).Intentional inhibition in human action:The power of ‘no’.Neuroscience&BiobehavioralReviews,36(4),1107-1118.
Fishbach,A.,Friedman,R.S.,& Kruglanski,A.W.(2003).Leading us not into temptation:Momentary allurements elicit overriding goal activation.JournalofPersonalityandSocialPsychology,84(2),296.
Fitzsimons,G.M.,& Bargh,J.A.(2003).Thinking of you:Nonconscious pursuit of interpersonal goals associated with relationship partners.JournalofPersonalityandSocialPsychology,84(1),148.
Gray,H.,Ambady,N.,Lowenthal,W.,& Deldin,P.(2004).P300 as an index of attention to self-relevant stimuli.JournalofExperimentalSocialPsychology,40(2),216-224.
Hu,X.,Wu,H.,& Fu,G.(2011).Temporal course of executive control when lying about self-and other-referential information:An ERP study.BrainRes,1369,149-157.
Karayanidis,F.,& Michie,P.(1996).Frontal processing negativity in a visual selective attention task.ElectroencephalographyandClinicalNeurophysiology,99(1),38-56.
Kenning,P.,& Plassmann,H.(2005).Neuro Economics:An overview from an economic perspective.BrainResearchBulletin,67(5),343-354.
Kiehl,K.A.,Laurens,K.R.,Duty,T.L.,Forster,B.B.,& Liddle,P.F.(2001).Neural sources involved in auditory target detection and novelty processing:An event-related fMRI study.Psychophysiology,38(01),133-142.
Kruglanski,A.W.,Shah,J.Y.,Fishbach,A.,Friedman,R.,Chun,W.Y.,& Sleeth-Keppler,D.(2002).A theory of goal systems.AdvancesinExperimentalSocialPsychology,34,331-378.
Morsella,E.,Bargh,J.A.,& Gollwitzer,P.M.(2008).Oxfordhandbookofhumanaction(Vol.2).USA.Oxford University Press.
Näätänen,R.,Simpson,M.,& Loveless,N.E.(1982).Stimulus deviance and evoked potentials.BiologicalPsychology,14(1-2),53-98.
Shah,J.Y.,& Kruglanski,A.W.(2002).Priming against your will:How accessible alternatives affect goal pursuit.JournalofExperimentalSocialPsychology,38(4),368-383.
Shah,J.Y.,& Kruglanski,A.W.(2003).When opportunity knocks:Bottom-up priming of goals by means and its effects on self-regulation.JournalofPersonalityandSocialPsychology,84(6),1109.
Simonson,I.(2005).In defense of consciousness:The role of conscious and unconscious inputs in consumer choice.JournalofConsumerPsychology,15(2),211-217.
Thorpe,S.,Fize,D.,& Marlot,C.(1996).Speed of processing in the human visual system.Nature,381(6582),520-522.
Torres,A.,Catena,A.,Megías,A.,Maldonado,A.,Cándido,A.,Verdejo-García,A.,& Perales,J.C.(2013).Emotional and non-emotional pathways to impulsive behavior and addiction.FrontHumNeurosci,7,43.doi:10.3389/fnhum.2013.00043.
Yuan,J.,Yang,J.,Meng,X.,Yu,F.,& Li,H.(2008).The valence strength of negative stimuli modulates visual novelty processing:Electrophysiological evidence from an event-related potential study.Neuroscience,157(3),524-531.
Zhang,D.,Gu,R.,Wu,T.,Broster,L.S.,Luo,Y.,Jiang,Y.,& Luo,Y.J.(2013).An electrophysiological index of changes in risk decision-making strategies.Neuropsychologia,51(8),1397-1407.
