微小隐孢子虫黏蛋白CGD5_2060原核表达及其黏附功能
2018-05-11阳毅敏潘灵韬庄浩瀚孙洪超杨怡陈学秋杜爱芳
阳毅敏,潘灵韬,庄浩瀚,孙洪超,杨怡,陈学秋,杜爱芳
(浙江大学动物科学学院动物预防医学研究所,杭州310058)
隐孢子虫是一种可以引起严重腹泻疾病的顶复门寄生虫,在世界范围内分布广泛[1]。隐孢子虫病流行于许多资源匮乏的国家,并且可以在工业化国家娱乐用水中爆发[2]。该疾病自限于免疫健全的宿主,但可使免疫力低下的个体虚弱,甚至致命,尤其是在资源有限地区未经治疗的艾滋病患者[3]和营养不良的儿童[1]。隐孢子虫是亚洲和非洲幼儿大多数中到重度腹泻病的4种病原体之一,是造成这些儿童腹泻和死亡的第2大病因[4]。尽管如此,目前仍缺乏有效的治疗措施[5],因此迫切需要鉴定出能够开发新型干预措施的分子靶点。
参与介导隐孢子虫和宿主细胞相互作用的蛋白质是干预隐孢子虫病的潜在靶标,但是已被表征的很少。隐孢子虫在体外难以繁殖和进行遗传操作,阻碍了新分子靶标的发现和验证[6-7]。尽管如此,许多研究已经证明,黏蛋白样糖蛋白和凝集素在介导体外和体内隐孢子虫感染中发挥了重要的作用[8-9]。隐孢子虫黏附和入侵过程的特征表明,这种寄生虫可能遵循所发现的其他顶复门寄生虫黏附和入侵的机制[10]。在与宿主细胞接触后,特有的顶端复合体细胞器(棒状体、致密颗粒和微线体)释放参与宿主细胞识别和黏附的抗原,入侵并形成纳虫空泡[11]。与其他顶复门原虫不同的是,黏蛋白样糖蛋白可能参与了隐孢子虫的这些过程。黏蛋白是由20%~55%丝氨酸、苏氨酸和脯氨酸残基组成的糖蛋白,由于丝氨酸和苏氨酸残基的这种广泛的O端糖基化,蛋白分子质量的40%~80%由O端相连的糖类组成[12]。本试验通过对黏蛋白cgd5_2060基因克隆表达后,进行细胞黏附功能的初步研究,为进一步揭示其参与微小隐孢子虫(Cryptosporidium parvum)入侵宿主细胞的作用机制提供理论依据。
1 材料与方法
1.1 主要材料与试剂
6~8周雌性ICR小鼠购自浙江省医学科学院。微小隐孢子虫卵囊由河南农业大学张龙现教授惠赠,由本实验室保存。克隆载体pMD18-T购自宝生物工程(大连)有限公司,表达载体pET-32a、大肠埃希菌Escherichia coli TOP10、E.coli Rosetta、Caco2细胞等均由本实验室保存。镍琼脂糖凝胶FF购自北京韦氏博慧色谱科技有限公司;蛋白质标志物购自美国Thermo Fisher科技公司;聚偏二氟乙烯(PVDF)膜购自美国Millipore公司;抗His标签鼠源单克隆抗体、HRP标记的山羊抗小鼠IgG抗体购自美国Sigma公司。
1.2 基因扩增和克隆
根据微小隐孢子虫cgd5_2060的基因序列(GenBank:XM_626146.1),用Primer Premier 5.0软件设计引物,并在上下游引物的5'端分别引入Bam HⅠ和SalⅠ的酶切位点,序列如下。F1:5′-CG ggatccATGAAGTTCATAAGATTTGGTTT-3′;R1:5′-GCgtcgacTTATTCAGAATCTTCATCCTGG-3′。设计完成后由生工生物工程(上海)股份有限公司合成。用TRIzol试剂(购自美国Invitrogen公司)提取微小隐孢子虫卵囊总RNA后,按照PrimeScript®RT试剂盒(购自日本TaKaRa公司)说明书反转录合成cDNA,用合成的引物进行聚合酶链式反应(polymerase chain reaction,PCR)扩增。PCR反应程序为:98℃预变性3 min;94℃变性15 s,58℃退火15 s,72℃延伸2 min,35个循环;72℃延伸10 min。产物经1%琼脂糖凝胶电泳检测后,按照凝胶回收试剂盒(购自美国Axygen公司)说明书进行回收,回收产物与pMD18-T载体连接并转化至大肠埃希菌TOP10中,菌液经PCR鉴定后,阳性样品由铂尚生物技术(上海)有限公司进行测序分析。
1.3 基因的序列比对及生物信息学分析
用MegAlign软件将测序反馈的结果与在GenBank中已登录的隐孢子虫cgd5_2060基因序列进行比对分析。使用SignalP4.1软件预测CGD5_2060蛋白是否具有信号肽,是否为分泌型蛋白。使用PROSITE数据库对CGD5_2060蛋白的功能性基序进行预测,并用软件DOG 1.0[13]绘制其结构示意图。
1.4 原核表达重组质粒的构建
对蛋白CGD5_2060进行序列分析的结果表明,其前19个氨基酸为信号肽区域,为分泌型蛋白。信号肽对其在进行原核表达时会有一定的影响,据此设计去信号肽引物,并在上下游引物的5'端分别引入Bam HⅠ和SalⅠ的酶切位点,序列如下。F2:5′-CGggatccaatggggaaaacAGTATTTCTTT-3′;R2:5′-GCgtcgacTTATTCAGAATCTTCATCCT-3′。设计完成后由生工生物工程(上海)股份有限公司合成。用上述引物通过PCR扩增测序正确的质粒,电泳后回收PCR产物并连接至pMD18-T载体上,随后转化到大肠埃希菌TOP10内,挑取单菌落进行PCR鉴定和测序分析。用限制性内切酶Bam HⅠ和SalⅠ对测序正确的质粒pMD18TS-cgd5_2060和表达载体pET-32a进行双酶切,电泳后分别回收双酶切产物,用T4 DNA连接酶连接2种产物后转化入E.coli Rosetta中,挑取单菌落进行PCR鉴定,提取阳性菌的质粒进行酶切鉴定,测序正确后进行保种,将菌命名为pET32a-cgd5_2060-Rosetta。同时,将pET32a空载体质粒转化至E.coli Rosetta中,挑取单菌落,命名为pET32a-Rosetta。
1.5 重组蛋白的表达和纯化
将菌液pET32a-cgd5_2060-Rosetta和pET32a-Rosetta按照1∶100接种于5 mL含氨苄青霉素的LB液体培养基中,37℃、190 r/min培养过夜,然后按照1∶50接种至新鲜的5 mL氨苄抗性LB液体培养基中,37℃振荡培养至λ(600 nm)=0.6(约2.5 h),加入终浓度为0.5 mmol/L的异丙基硫代半乳糖苷(isopropylβ-D-thiogalactoside,IPTG),在37 ℃下进行诱导,在不同时间点取样,收集菌体后,进行十二烷基硫酸钠-聚丙烯酰氨凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)分析。将蛋白表达成功的菌液过夜培养后按照1∶50接种至新鲜的300 mL含氨苄抗性的LB液体培养基中,37℃、190 r/min摇床培养至λ(600 nm)=0.6(约2.5 h),添加终浓度为0.1 mmol/L的IPTG,16℃低温诱导12 h,菌液经8 000 r/min离心5 min,按照镍琼脂糖凝胶FF说明书对表达的重组蛋白和载体蛋白进行纯化。
1.6 多克隆抗体的制备
采用SDS-PAGE方法对纯化的重组蛋白CGD5_2060进行鉴定,用考马斯亮蓝法(Bradford法)测定重组蛋白的浓度。将纯化的重组蛋白CGD5_2060作为抗原免疫6~8周雌性ICR小鼠。首次免疫采用弗氏完全佐剂乳化(50μg/只),之后的免疫均用弗氏不完全佐剂乳化(25μg/只)。免疫前对每只小鼠采血作为阴性对照,采取皮下多点注射法免疫,首次免疫后14 d进行第二次免疫,二免后10 d进行第三次免疫,三免后7 d采小鼠尾静脉血,用间接酶联免疫吸附测定(enzyme-linked immuno-sorbent assay,ELISA)法检测其血清抗体的效价,达到目标后即进行眼眶后静脉丛采血,收集血清。
1.7 重组蛋白的蛋白质印迹法鉴定
通过蛋白质印迹法对纯化的重组蛋白CGD5_2060进行分析,一抗为抗His标签鼠源单克隆抗体,稀释倍数为1∶1 000,二抗为HRP-羊抗小鼠IgG,稀释倍数为1∶5 000,用增强化学发光法(enhanced chemiluminescence,ECL)进行显色。同时,对制备的鼠抗CGD5_2060血清进行蛋白质印迹法鉴定,一抗为1.6节中收集的血清,稀释倍数为1∶2 000。
1.8 重组蛋白与Caco2细胞黏附特性的检测
1.8.1 蛋白质印迹法检测
参照文献[14]中的细胞结合实验进行蛋白质印迹法检测。即先将Caco2细胞经胰酶消化后吹打分散成单个细胞,铺至6孔细胞培养板中,置于37℃、5%CO2培养箱中培养,密度达80%~90%后,加入预热至37℃的含1 mmol/L CaCl2、0.5 mmol/L MgCl2和2%胎羊血清的1640,于37 ℃、5%CO2培养箱中封闭30 min。用含1 mmol/L CaCl2、0.5 mmol/L MgCl2的1640洗涤2次后,将1 mL稀释至约200μg/mL的重组蛋白CGD5_2060和载体对照蛋白加入相应的孔中,孵育2 h。孵育后用含1 mmol/L CaCl2、0.5 mmol/L MgCl2的1640洗去未结合的蛋白质。收集细胞制成蛋白样品,用抗His标签鼠源单克隆抗体作为一抗,碱性磷酸酶标记的山羊抗小鼠IgG作为二抗进行蛋白质印迹法检测,观察蛋白与细胞的黏附情况。
1.8.2 间接ELISA检测
参照文献[15]中的方法进行间接ELISA检测。即先将Caco2细胞经胰酶消化后吹打分散成单个细胞,铺至96孔细胞培养板中,置于37℃、5%CO2培养箱中培养至单层后进行后续试验。用含1%戊二醛的磷酸盐缓冲液(phosphate buffer saline,PBS),每孔50μL,室温固定30 min。用含5%胎羊血清的PBS,每孔150μL,室温封闭4 h。将用含1 mmol/L CaCl2、0.5 mmol/L MgCl2和0.1%胎羊血清的PBS稀释成不同浓度的重组蛋白CGD5_2060,加入至细胞培养板中,每孔100μL,每个稀释倍数3个重复,载体对照蛋白作相同处理。抗His标签鼠源单克隆抗体作为一抗,碱性磷酸酶标记的山羊抗小鼠IgG作为二抗,进行间接ELISA测定,检测蛋白与细胞的黏附情况。
2 结果与分析
2.1 目的基因的扩增与克隆
以微小隐孢子虫cDNA为模板,使用设计的引物进行PCR扩增,获得的特异性片段长度为1 854 bp(图略)。
2.2 序列分析
用MegAlign软件将测序所得序列与在GenBank中登录的隐孢子虫cgd5_2060序列进行比对分析。结果显示,基因核苷酸序列同源性为99.9%,氨基酸序列同源性为100%。使用SignalP 4.1软件分析的结果表明,CGD5_2060的1~19个氨基酸是其信号肽,说明该蛋白是分泌型蛋白。使用PROSITE数据库分析发现:CGD5_2060含有1个富含苏氨酸的区域TTTTTTKSTTTTTTAVTT,位于第510~527个氨基酸处;此外,还有1个与细胞黏附有关的序列RGD,位于第69~71个氨基酸处,用DOG 1.0软件绘制的蛋白质结构域示意图如图1所示。

图1 CGD5_2060蛋白结构示意图Fig.1 Structural schematic diagram of CGD5_2060 protein
2.3 原核表达重组质粒的构建分析
对重组克隆质粒pMD18TS-cgd5_2060进行Bam HⅠ和SalⅠ双酶切,可见预期大小的载体条带和目的基因条带(图2A)。将pMD18TS-cgd5_2060双酶切回收产物与pET32a表达载体双酶切回收产物连接转化后,挑取单克隆菌落进行PCR鉴定(图2B),重组质粒pET32a-cgd5_2060经Bam HⅠ和SalⅠ双酶切后,可见大小约为1 788 bp的目的条带及5 900 bp左右的载体条带(图2C),与预期一致。测序后与GenBank公布的序列进行比对,氨基酸同源性为100%,表明cgd5_2060基因已成功插入pET32a中,重组原核表达质粒构建正确。
2.4 重组蛋白的诱导表达、纯化及蛋白质印迹法分析
2.4.1 重组质粒pET32a-cgd5_2060的诱导表达
将含有质粒pET32a-cgd5_2060和pET32a的Rosetta大肠埃希菌经IPTG诱导表达后,进行SDSPAGE电泳分析。结果显示,在约87和22 kDa处有表达目的蛋白的趋势,与理论推算的蛋白产物相对分子质量相符(图3)。随着诱导时间的延长,蛋白表达量升高,且在6 h达到较高水平。
2.4.2 蛋白纯化
蛋白经大量诱导表达后,于8 000 r/min、4℃下离心5 min,向菌体沉淀中加入50 mmol/L PBS(Na2HPO4·12H2O 1.45 g,NaCl 2.93 g,NaH2PO4·2H2O 0.148 g,加双蒸水至100 mL,调节pH至7.4)充分溶解,经超声波裂解、离心后,按照镍琼脂糖凝胶柱说明书进行蛋白纯化,并对纯化后的处理样品进行SDS-PAGE电泳分析。结果(图4)显示,2种蛋白均可由含250 mmol/L咪唑的PBS洗脱,效果很好。纯化后的蛋白经透析、超滤浓缩后,测定其重组蛋白CGD5_2060和载体对照蛋白的质量浓度分别为1.5和2.1 mg/mL。

图2 重组质粒的鉴定Fig.2 Identification of recombinant plasmid
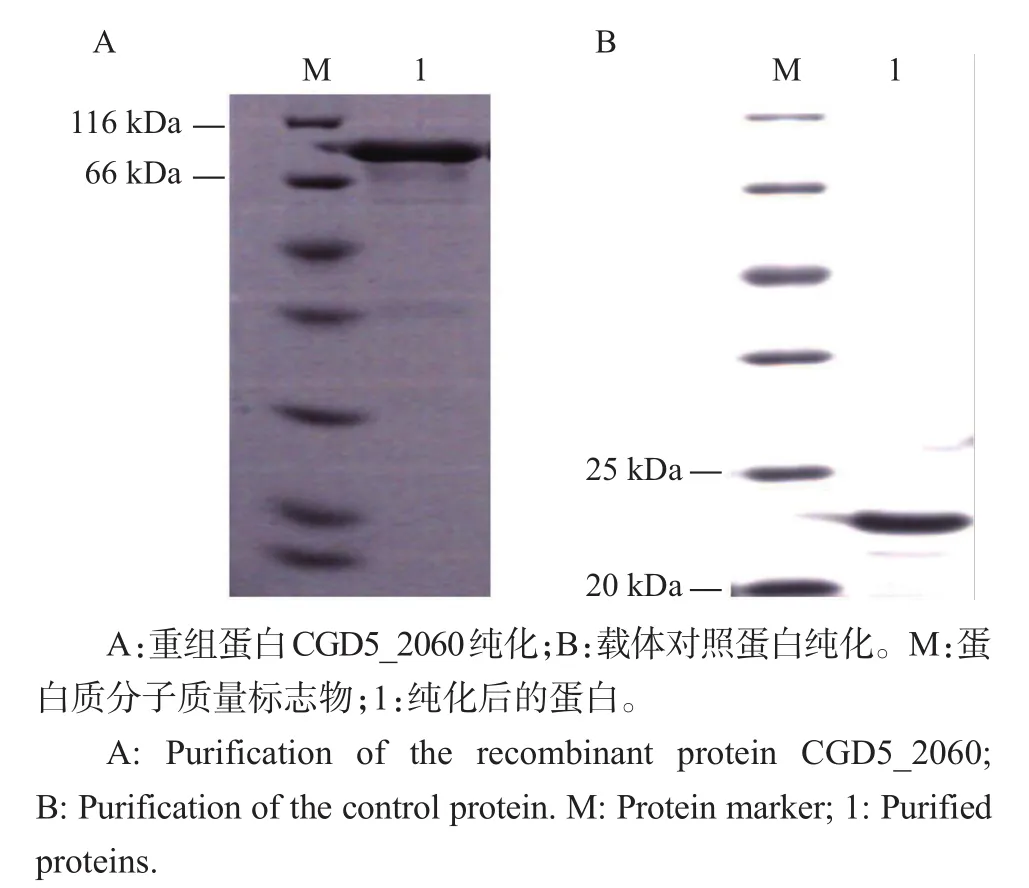
图4 蛋白的纯化Fig.4 Purification of proteins
2.4.3 蛋白质印迹法检测结果
以抗His标签鼠源单克隆抗体为一抗进行蛋白质印迹法检测。结果(图5)显示,重组蛋白CGD5_2060和载体对照蛋白均可与His标签鼠源单克隆抗体特异性结合,且大小与预期相符,说明得到的蛋白正确。
2.5 抗体效价的间接ELISA检测及蛋白质印迹法分析
以重组蛋白CGD5_2060作为抗原免疫6~8周雌性ICR小鼠3次后,收集血清,进行倍比稀释,从1∶2 000至1∶2 048 000,并设置阴性对照和空白对照。间接ELISA检测结果显示,最后所得的血清抗体效价为1∶512 000~1∶1 024 000。将所得的血清与重组蛋白进行蛋白质印迹法分析,结果(图6)显示,制备的多克隆抗体可以与重组蛋白特异性结合。

图5 纯化后蛋白的蛋白质印迹法分析Fig.5 Western blotting analysisof purified proteins

图6 小鼠多抗与重组蛋白的蛋白质印迹法分析Fig.6 Western blotting analysis of mouse polyclonal antibody against purified proteins
2.6 重组蛋白与Caco2细胞黏附的检测结果
对照蛋白和重组蛋白CGD5_2060与Caco2细胞孵育后,蛋白质印迹法分析结果(图7)显示:最后一次洗涤两者相对应的位置均没有出现条带,说明最后一次洗涤已经将所有未结合的蛋白全部洗净;而在最后收集的细胞裂解液中,重组蛋白CGD5_2060在其相应位置出现条带,且对照蛋白相应位置没有条带,说明CGD5_2060重组蛋白能黏附Caco2细胞。将不同浓度稀释的2种蛋白与Caco2细胞孵育后进行间接ELISA检测,结果(图8)显示,对照蛋白对Caco2细胞没有黏附作用,而重组蛋白可以与Caco2细胞黏附,其结合量在一定范围内随着蛋白浓度的增加而增加,随后基本趋于稳定。

图7 蛋白质印迹法检测重组蛋白与Caco2细胞黏附特性的结果Fig.7 Western blot ting analysis of the recombinant protein CGD5_2060 adhering to Caco2 cells

图8 间接ELISA检测重组蛋白与Caco2细胞黏附特性的结果Fig.8 Indirect ELISA analysis of the recombinant protein CGD5_2060 adhering to Caco2 cells
3 讨论
尽管隐孢子虫具有重要的公共卫生学意义,但是很少有探究这种病原体与宿主的相互作用,并且仅很少的寄生虫抗原被表征。然而,已经被充分研究涉及宿主细胞侵入过程的抗原,均显示或被预测具有黏蛋白型O-糖基化。隐孢子虫基因组含有31个编码推测为黏蛋白抗原的开放性阅读框,这进一步突出了这类抗原在隐孢子虫与宿主相互作用中的重要性。迄今为止,被确定在隐孢子虫黏附和入侵过程中不可或缺的4种抗原都是糖蛋白,包 括 CSL[16]、GP900[17]、GP40/15[18]和 P23[19]。GP900和GP40/15都是黏蛋白样糖蛋白,而P23预测含有黏蛋白型O-糖基化位点。
本研究从微小隐孢子虫cDNA中克隆获得了1个黏蛋白基因cgd5_2060,并对其蛋白功能相关基序进行了预测。结果表明,该蛋白的1~19个氨基酸可能为信号肽,属于分泌型蛋白。在PROSITE数据库中进行比对分析发现:CGD5_2060蛋白含有1个富含苏氨酸的区域TTTTTTKSTTTTTTAVTT,位于510~527个氨基酸处,属于黏蛋白样结构域;此外,还有1个与细胞黏附有关的氨基酸序列,由3个氨基酸组成,分别为精氨酸-甘氨酸-天冬氨酸(RGD),位于69~71个氨基酸处。对CGD5_2060蛋白功能相关基序的预测结果表明,cgd5_2060基因可能与微小隐孢子虫黏附宿主细胞相关。
本研究成功构建了原核表达载体pET32acgd5_2060,并在大肠埃希菌Rosetta中成功表达了该重组蛋白,位于菌体超裂后的上清液中。通过镍柱亲和层析法获得了较高质量的重组蛋白CGD5_2060,免疫ICR小鼠获得了高效价的多克隆抗体,为进一步研究CGD5_2060蛋白在虫体不同阶段中的定位,以及该多抗对虫体黏附、入侵和增殖的影响提供了材料。
微小隐孢子虫依赖黏蛋白抗原黏附和入侵宿主细胞,而Caco2细胞系是隐孢子虫研究中常用的细胞系。本研究将纯化的重组蛋白与Caco2细胞共孵育,通过蛋白质印迹法和间接ELISA法检测了其与宿主细胞的黏附能力。蛋白质印迹法分析结果表明,CGD5_2060重组蛋白可以黏附Caco2细胞。间接ELISA检测表明,不同浓度稀释的重组蛋白与Caco2细胞孵育后,其结合量在一定范围内随着蛋白浓度的增加而增加,随后基本趋于稳定。
本研究初步验证了cgd5_2060是隐孢子虫感染时重要的黏蛋白基因。后续利用抗体体外阻断实验和建立微小隐孢子虫cgd5_2060基因敲除株,对于研究该基因在虫体黏附、入侵过程中发挥的作用及其介导黏附的机制,理解隐孢子虫的生物学特性及开发疫苗具有重要意义。
参考文献(References):
[1]CHECKLEY W,WHITE JR A C,JAGANATH D,et al.A review of the global burden,novel diagnostics,therapeutics,and vaccine targets for Cryptosporidium.The Lancet Infectious Diseases,2015,15(1):85-94.
[2]HLAVSA M C,ROBERTS V A,KAHLER A M,et al.Recreational water-associated disease outbreaks:United States,2009—2010.Morbidity and Mortality Weekly Report,2014,63(1):6-10.
托架沿Z向运动,通过激光跟踪仪测量定位器1~4的z向移动副上的任一定点的空间位置变化,可求解矩阵T中第8、14、17、20列对应的结构误差;托架绕S1S4旋转,测量定位器2的x向移动副上任一定点的空间位置变化,可以计算矩阵T中第9列对应的结构误差。
[3]O’CONNOR R M,SHAFFIE R,KANG G,et al.Cryptosporidiosisin patientswith HIV/AIDS.AIDS,2011,25(5):549-560.
[4]KOTLOFF K L,NATARO J P,BLACKWELDER W C,et al.Burden and aetiology of diarrhoeal disease in infants and young children in developing countries(the Global Enteric Multicenter Study,GEMS):A prospective,case-control study.The Lancet,2013,382(9888):209-222.
[5]CABADA M M,WHITE JR A C.Treatment of cryptosporidiosis:Do we know what we think we know?Current Opinion in Infectious Diseases,2010,23(5):494-499.
[6]MORADA M,LEE S,GUNTHER-CUMMINS L,et al.Continuous culture of Cryptosporidium parvum using hollow fiber technology.International Journal for Parasitology,2016,46(1):21-29.
[7]VINAYAK S,PAWLOWICM C,SATERIALEA,et al.Genetic modification of the diarrhoeal pathogen Cryptosporidium parvum.Nature,2015,523(7561):477-480.
[8] LENDNERM,DAUGSCHIESA.Cryptosporidium infections:Molecular advances.Parasitology,2014,141(11):1511-1532.
[9] WANYIRIJ,WARD H.Molecular basisof Cryptosporidiumhost cell interactions:Recent advances and future prospects.Future Microbiology,2006,1(2):201-208.
[10]WETZEL D M,SCHMIDT J,KUHLENSCHMIDT M S,et al.Gliding motility leads to active cellular invasion by Cryptosporidium parvum sporozoites.Infection and Immunity,2005,73(9):5379-5387.
[11]SHARMA P,CHITNIS C E.Key molecular events during host cell invasion by Apicomplexan pathogens.Current Opinion in Microbiology,2013,16(4):432-437.
[12]VAN KLINKEN B J,DEKKER J,BULLER H A,et al.Mucin gene structure and expression:Protection vs.adhesion.American Journal of Physiology:Gastrointestinal and Liver Physiology,1995,269(5):G613-G627.
[13]REN J,WEN L P,GAO X J,et al.DOG 1.0:Illustrator of protein domain structures.Cell Research,2009,19(2):271-273.
[14]SAOUROSS,EDWARD-JONESB,REISSM,et al.A novel galectin-like domain from Toxoplasma gondii micronemal protein 1 assiststhefolding,assembly,and transport of acell adhesion complex.Journal of Biological Chemistry,2005,280(46):38583-38591.
[15]O’CONNOR R M,BURNS P B,HA-NGOC T,et al.Polymorphic mucin antigens CpMuc4 and CpMuc5 are integral to Cryptosporidium parvum infection in vitro.Eukaryotic Cell,2009,8(4):461-469.
[16]LANGER R C,RIGGS M W.Cryptosporidium parvum apical complex glycoprotein CSL contains a sporozoite ligand for intestinal epithelial cells.Infection and Immunity,1999,67(10):5282-8291.
[17]PETERSEN C,GUT J,DOYLE PS,et al.Characterization of a>900 000-M(r)Cryptosporidium parvum sporozoite glycoprotein recognized by protective hyperimmune bovine colostral immunoglobulin.Infection and Immunity,1992,60(12):5132-5138.
[18]STRONG W B,GUT J,NELSON R G.Cloning and sequence analysis of a highly polymorphic Cryptosporidium parvum gene encoding a 60-kilodalton glycoprotein and characterization of its 15-and 45-kilodalton zoite surface antigen products.Infection and Immunity,2000,68(7):4117-4134.
[19]PERRYMANL E,JASMERDP,RIGGSM W,et al.Acloned gene of Cryptosporidium parvum encodes neutralizationsensitive epitopes.Molecular and Biochemical Parasitology,1996,80(2):137-147.
