产纤维素酶细菌菌株的分离鉴定及产酶条件优化
2018-05-10毛丽春修立辉
毛丽春,修立辉,胡 刚
(广西师范学院 环境与生命科学学院,广西 南宁 530001)
人类对于能源的需求不断增加,化石能源作为人类最主要的消耗能源具有一次性、有限性等特点,其的燃烧对环境造成严重污染。这些因素导致一种可再生、可持续发展的能源物质将其取代。而将生物质通过酶转化为糖再发酵成为乙醇被认为是产生燃料和化学物质的可行方案[1]。纤维素是具有高度结晶结构的物质,由D-葡萄糖通过β-1,4键连接而成的线性聚合物。其被一层坚硬的木质素层所围绕,因此将纤维素水解成可利用的葡萄糖是非常困难的[2]。纤维素通过外切葡聚糖酶、内切葡聚糖酶和β-葡萄糖苷酶3种酶的协同作用水解成葡萄糖[3]。内切葡聚糖酶负责切割纤维素链内部β-糖苷键,外切葡聚糖酶负责切割纤维素链的末端产生纤维二糖,最后β-葡萄糖苷酶水解纤维二糖形成葡萄糖[4]。纤维素酶同大多数酶类一样,有自己特殊的活性区域[5]。其反应区域包含两个部分,即供底物结合的糖类结合域和分解底物的催化域,两个区域通过灵活的链接器连接[6-7]。目前发现能够降解纤维素酶的种类有Bacillus sp.、Pseudomonas sp.、Paenibacillus sp.、Lysinibacillus sp.、Micrococcus sp.、Xanthomonas sp.、Brucella sp.和Vibrio xiamenensis等[8]。
本研究经过初筛及液体发酵摇瓶培养得到高产纤维素酶菌株,对该菌株进行形态学分析和分子生物学鉴定,并对其产酶条件进行了优化,以期为进一步研究细菌纤维素酶生产菌株降解纤维素的机制提供实际应用价值。
1 材料与方法
1.1 材料与试剂
土壤样品(31份):采集自广西省富川县西岭山原始森林保护区;甘蔗渣:南宁市制糖厂;羧甲基纤维素钠(carboxy methyl cellulose sodium,CMC-Na)、微晶纤维素:美国Sigma公司;2×Easy Taq聚合酶链式反应(polymerase chain reaction,PCR)SuperMix(+dye)和DNA分子量Marker:北京全式金生物技术有限公司;脱氧核糖核酸(deoxyri bonucleic acid,DNA)回收纯化试剂盒:北京博迈德生物技术有限公司;四水合酒石酸钾钠、NaOH、Na2SO4、结晶酚和3,5-二硝基水杨酸(均为分析纯)等:国药集团化学试剂公司;琼脂粉:北京索莱宝科技有限公司;细菌16SrRNA基因通用引物F27(5′-AGAGTTTGATCCTGGCTCAG-3′)和 R1492(5′-TACGGTTACCTTGTTACGACTT-3′):生工生物工程(上海)股份有限公司合成。
初筛培养基:NaNO32 g/L,MgSO4·7H2O 0.5 g/L,NaCl 0.5g/L,FeSO4·7H2O 0.01g/L,KH2PO41.0g/L,酵母膏0.4g/L,甘蔗渣5 g/L,刚果红0.2 g/L,琼脂粉15.0 g/L,pH 5.0。
复筛培养基:KH2PO42.0 g/L,(NH4)2SO41.4 g/L,CaCl20.3 g/L,MgSO4·7H2O 0.3 g/L,酵母膏0.4 g/L,微量元素溶液1 mL,琼脂粉15 g/L,pH 5.0。
GBM培养基:牛肉膏2 g/L,酵母提取物2 g/L,蛋白胨5 g/L,蔗糖6 g/L,pH自然。另添加15 g琼脂粉则为GBM固体培养基。112℃灭菌20 min。
液体发酵培养基:KH2PO41.5 g/L,Na2HPO41.3 g/L,(NH4)2SO41.5 g/L,MgSO4·7H2O 0.7 g/L,CaCl20.1 g/L,微量元素溶液1 mL,CMC-Na10 g/L,pH7.0。
微量元素溶液组成:FeSO4·7H2O 0.005 g/L,MnSO40.001 6 g/L,ZnCl20.001 7 g/L,CoCl20.002 g/L。
1.2 仪器与设备
UV-5500(PC)紫外可见分光光度计:上海精密仪器仪表有限公司;DYY-6C型电泳仪:北京六一生物科技有限公司;德安特ES-320分析天平:天津德安特传感技术有限公司;Thermo ST8R台式冷冻离心机:赛默飞世尔科技(中国)有限公司;ZWY-211B细菌培养摇床:上海智城分析仪器制造有限公司;Gel K8160凝胶成像系统:北京科创锐新生物科技有限公司;DK-420S水浴锅:上海精宏实验设备有限公司。
1.3 方法
1.3.1 菌株分离和纯化
以广西省贺州市富川瑶族自治县西岭山原始森林保护区为采样地,选取该区域土壤腐殖质区域表层2~20 cm处土壤进行采集,每个土样点大约采集50 g土壤并编号同时记录周围植被类型情况。将采集土壤各称量5g分别加入盛有50 mL蒸馏水的三角瓶中(250 mL),加入适量的玻璃珠,180r/min、28℃振荡30min。将打散混匀的土壤梯度稀释至10-1、10-2、10-3,每个梯度取100 μL混合液涂布于以甘蔗渣为唯一碳源的刚果红培养基中培养。将所有能够生长的菌株都转移到以羧甲基纤维素钠为唯一碳源的培养基中进行第二次初筛培养。培养48h后,使用0.2%的刚果红进行染色,再用1 mol/L NaCl进行脱色。最后将所有能够在第二次筛选培养基上生长的菌株进行液体发酵摇瓶培养。
1.3.2 16SrRNA基因序列的聚合酶链式反应扩增[9]
采用菌落PCR方式扩展16SrRNA的基因序列。PCR反应体系(25μL):Template 1μL,F27/R1492引物各1μL,2×Taq PCR Mix 12.5μL,ddH2O 9.5μL。反应条件为:94℃预变性4 min,94℃变性30 s,55℃退火20 s,72℃延伸90 s,进行30次循环,反应完后72℃再延伸10 min,PCR产物8℃保温60 min。
用浓度为0.8%的琼脂糖凝胶验证PCR产物,将验证成功的PCR产物进行纯化并测序。利用Cluster W软件将测序拼接后的序列与GenBank中的相似序列进行多种比对,再通过MEGA6.0软件中基于Kimura-2的Neighbor-Joining(NJ)法(bootsrap=500次)构建具有产纤维素酶活的菌株以及相关相似菌株的系统进化树。
1.3.3 菌株的形态学观察和生理生化鉴定
形态学观察:将细菌接种于GBM培养基上,观察其菌落形态并对其进行革兰氏染色。
生理生化鉴定:参照《伯杰氏细菌手册》进行。
1.3.4 菌株产酶条件优化
分别考察培养基碳源(麦麸、1%CMC-Na、滤纸和微晶纤维素)、氮源(胰蛋白胨、(NH4)2SO4、NH4NO3、酵母粉、牛肉粉、NH4Cl、NaNO3、尿素)、初始pH值(5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5),接种量(2%、4%、6%、8%、10%(V/V))对菌株XI31-1产酶的影响。
1.3.5 菌株XI31-1的CMCase酶学特性分析
分别考察菌株XI31-1产CMCase的最适pH及温度。
1.3.6 测定方法
纤维素酶活的测定:按照参考文献[10-12]的方法分别检测内切葡聚糖酶、外切葡聚糖酶、滤纸酶的酶活力。
2 结果与分析
2.1 土壤中产纤维素酶细菌的分离及纯化
从西岭山原始森林土壤分离产纤维素酶活的菌株,经过初筛获得33株菌株,经过复筛仅有16株细菌能够生长。
通过菌落PCR的方法将第二次初筛得到的16株细菌的16SrRNA基因片段进行扩增,用琼脂糖凝胶电泳验证其PCR产物,结果见图1。
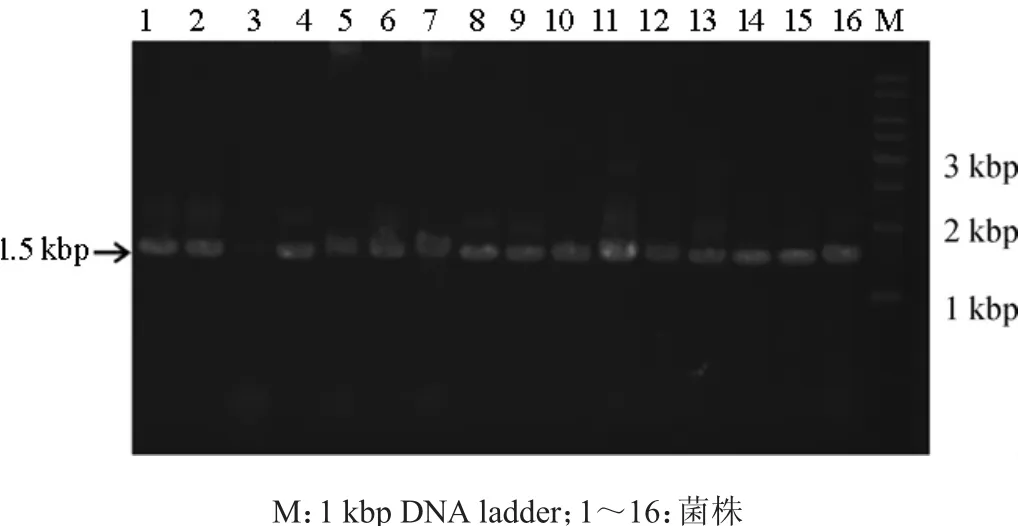
图1 细菌16S rRNA PCR产物凝胶电泳图Fig.1 Electrophoresis of PCR products of 16S rRNA of bacteria
由图1可知,表明所得片段大小约为1.5 kbp。将得到的PCR产物纯化后送武汉金开瑞生物工程有限公司测序。将测序得到的16S rRNA基因序列通过BLASTN分别在GenBank中进行比对,结果见表1,根据结果构建了系统进化树见图2。
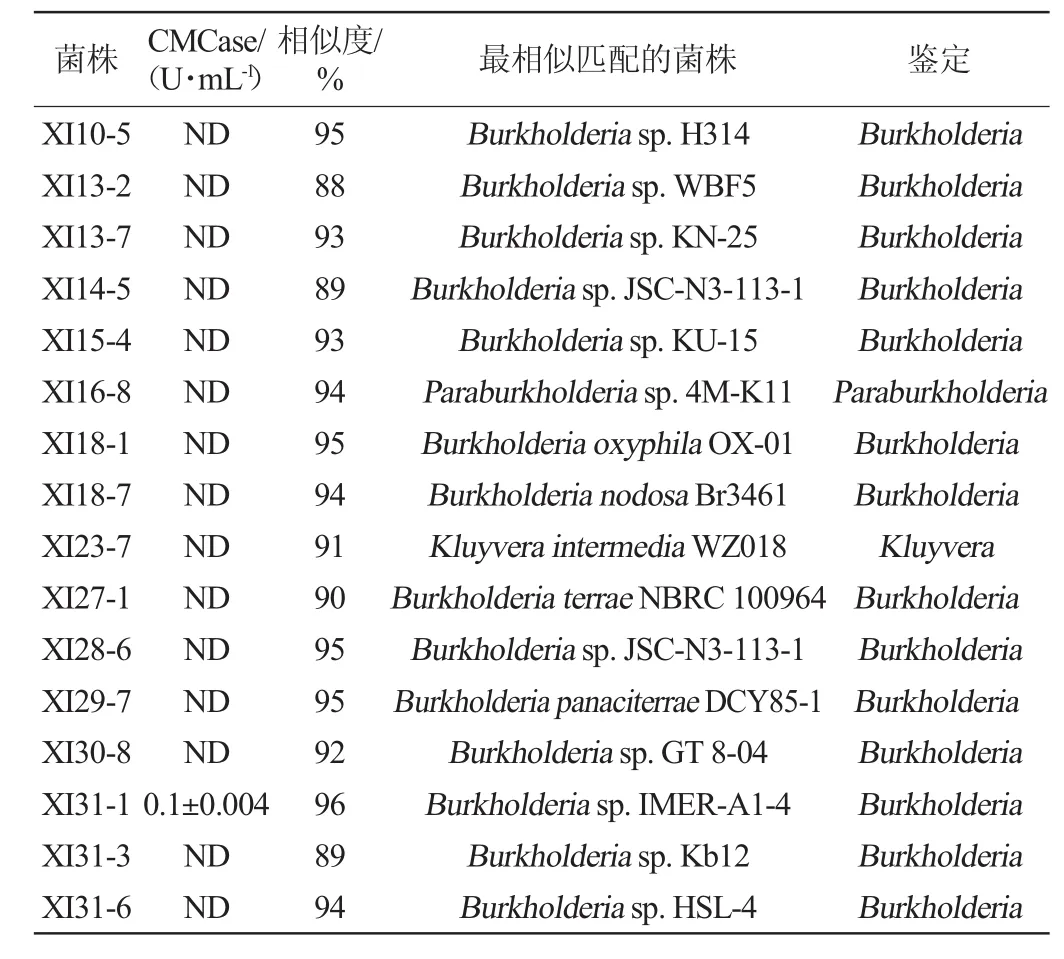
表1 16株细菌产纤维素酶活及鉴定结果Table 1 Cellulase activity and identification results of 16 bacteria strains
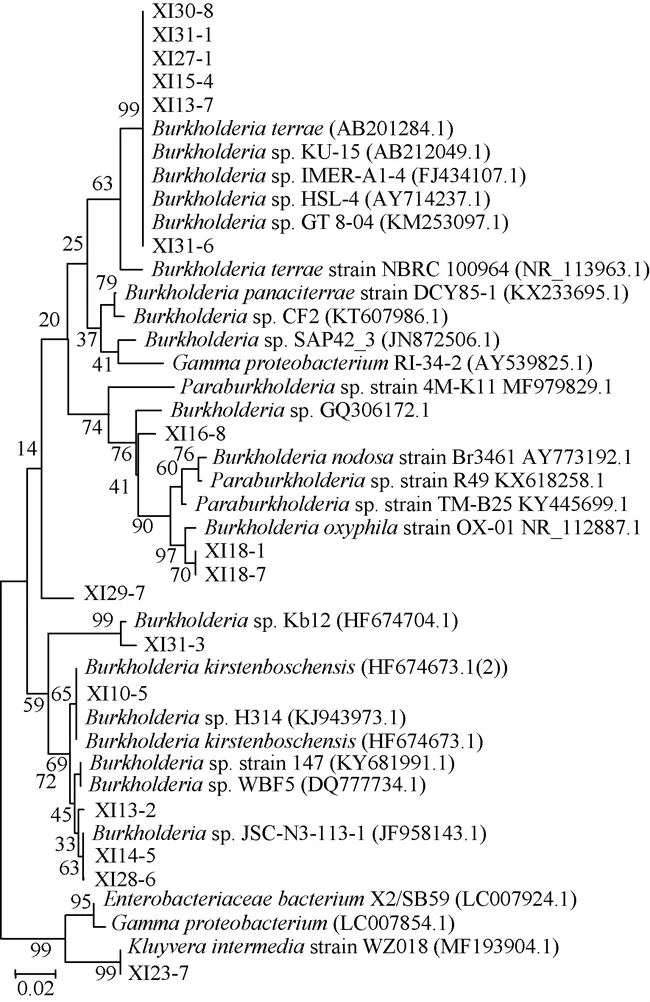
图2 16株细菌系统进化树Fig.2 Phylogenetic tree of 16 bacteria strains
由表1、图2可知,16株菌株分属于3个不同的属:伯克霍尔德菌属(Burkholderium 87.5%)、克吕沃氏菌属(Kluyvera 6.25%)、Paraburkholderia(6.25%),伯克霍尔德菌属菌株为西岭山中降解纤维素酶的优势菌群。16株细菌中仅菌株XI31-1具备产纤维素酶能力,故选择菌株XI31-1进行后续实验。
2.2 菌株的形态学观察和生理生化鉴定
2.2.1 形态学观察
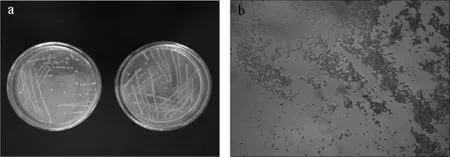
图3 菌株XI31-1的菌落形态(a)及菌株形态(b)Fig.3 Colonial(a)and morphology(b)of strain XI31-1
由图3可知,菌株XI31-11在GBM平板上呈乳白色,圆形,边缘完整,凸的,不透明,属革兰氏阴性菌。
2.2.2 生理生化鉴定实验
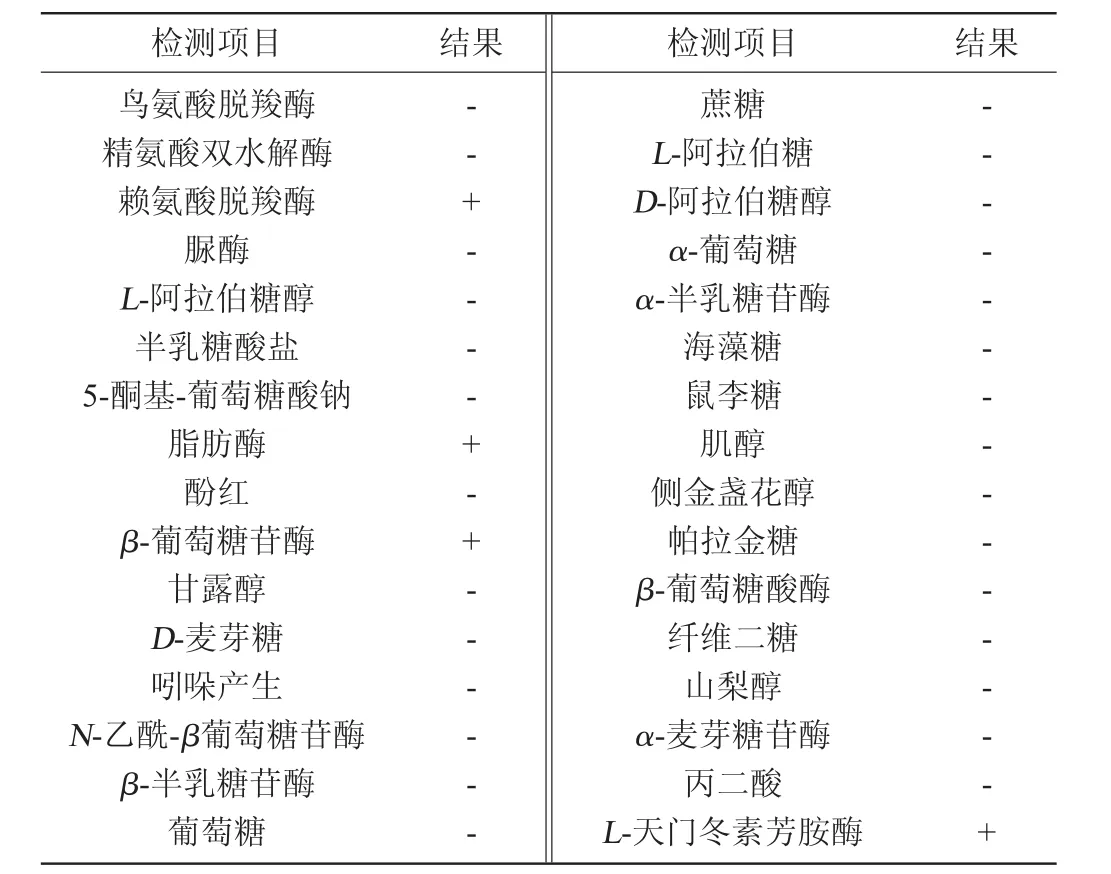
表2 菌株XI31-1的生理生化实验鉴定结果Table 2 Identification results of physiological and biochemical experiments of strain XI31-1
由表2可知,菌株XI31-1符合伯克霍尔德菌属的脂肪酶阳性和不利用麦芽糖等生化特征,为伯克霍尔德氏菌(Burkholderia cepacia)。
2.3 产酶条件优化
2.3.1 不同碳源对菌株XI31-1产酶的影响
在菌液接种量为4%,初始pH5.0,28℃摇床培养条件下,分别考察不同碳源对菌株XI31-1产酶的影响,结果见图4。
由图4可知,以麦麸和微晶纤维素为碳源时,其CMCase酶活力达到最高分别是2.5 U/mL和2.59 U/mL,滤纸次之,1%CMC-Na酶活力最低。鉴于麦麸和微晶纤维素为碳源时,其酶活力相差不大,但微晶纤维素价格昂贵,而麦麸廉价且环保,故选择麦麸作为该研究的碳源。
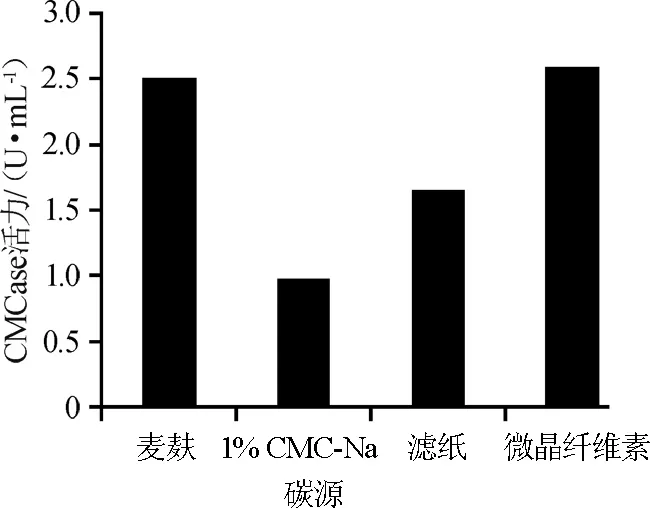
图4 不同碳源对菌株XI31-1产酶的影响Fig.4 Effects of different carbon sources on enzyme production by strain XI31-1
2.3.2 不同氮源对菌株XI31-1产酶的影响
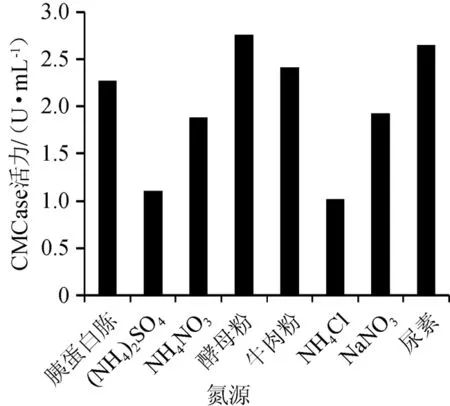
图5 不同氮源对菌株XI31-1产酶的影响Fig.5 Effects of different nitrogen sources on enzyme production by strain XI31-1
由图5可知,以酵母粉为氮源时其CMCase酶活力最高为2.76 U/mL。NH4Cl酶活力最低。因此选择酵母粉为培养基的最佳氮源。
2.3.3 不同初始pH值对菌株XI31-1产酶的影响
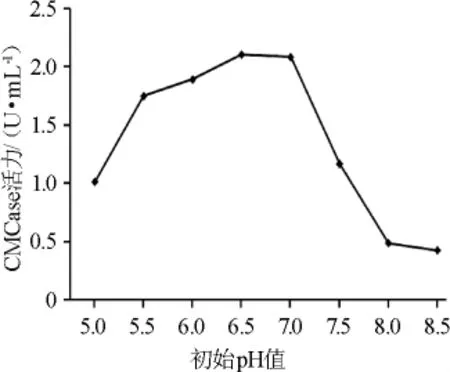
图6 初始pH值对菌株XI31-1产酶的影响Fig.6 Effect of initial pH on enzyme production by strain XI31-1
由图6可知,当初始pH值为7.0时,其CMCase酶活力最高为2.08 U/mL,继续升高初始pH值后其CMCase酶活力不断降低。因此选择初始pH值为7.0。
2.3.4 不同接种量对菌株XI31-1产酶的影响
在不同接种量对菌株产酶影响的预实验中,已证实当接种量为1%时,其CMCase酶活力低于接种量为2%的酶活力。由图7可知,随着接种量的不断升高,其CMCase酶活力有降低的趋势。因此选择最佳接种量为2%。
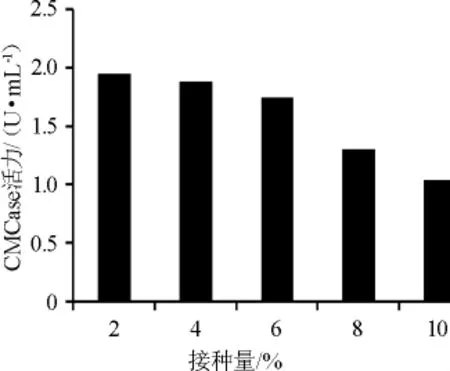
图7 不同接种量对菌株XI31-1产酶的影响Fig.7 Effects of different inoculum on enzyme production by strain XI31-1
2.4 菌株XI31-1的CMCase酶学特性分析
2.4.1 CMCase的最适pH值
在pH3.5、4.0、4.5、5.0、5.5、6.0、6.5条件下分别测定CMCase酶活。以最高酶活力为100%,计算各pH值条件下的CMCase相对酶活力,结果见图8。由图8可知,CMCase酶活在pH值为5.0时最高。

图8 不同pH值对CMCase酶活的影响Fig.8 Effects of different pH on CMCase activity
2.4.2 CMCase的最适温度
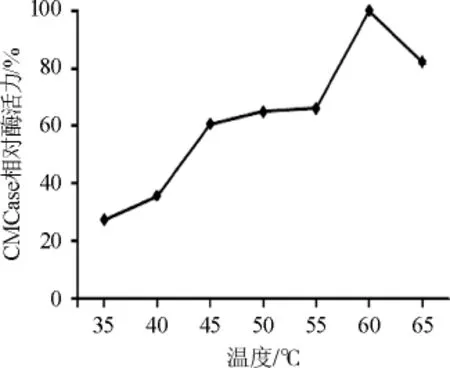
图9 不同温度对CMCase酶活的影响Fig.9 Effects of different temperature on CMCase activity
在35℃、40℃、45℃、50℃、55℃、60℃、65℃条件下分别测定CMCase酶活。以最高酶活力为100%计算各温度条件下的相对酶活力,结果见图9。由图9可知,CMCase酶活在温度为60℃时最高。
2.5 讨论
VARDAVKISE[13]研究认为纤维素酶的活力在春秋季节大于冬夏季节的活力,且深度为0~2 cm的森林土壤中有机物质的含量最高。有机物质和沙子内容物随着土壤深度的增加而减少,但淤泥和黏土内容物随着深度增加而增加。水解纤维素细菌的数量在0~2 cm的土壤中达到最大值,20~29 cm水解纤维素细菌的数量也处于最大值的状态。YANG JK等[14]的研究表明,落叶阔叶林的有机营养物高于常绿阔叶林,在落叶阔叶林中纤维素降解菌的种群密度和种群多样性高于常绿阔叶林,但常绿阔叶林更有可能含有高产纤维素酶的菌株。
对于产纤维素酶细菌菌株以及CMCase的研究中,AVELLANEDATORRESL M等[15]从哥伦比亚内瓦多斯国家自然公园筛选到了25株细菌,主要的产纤维素酶菌群是假单胞菌。SHAJAHAN S等[16]从印度中部马哈拉施特拉邦区域的拉贾布尔西部海岸热泉分离出的产纤维素酶菌为地衣芽孢杆菌(Bacillus licheniformis)。MAKIM L等[17]从造纸污泥和有机肥料中筛选到的主要纤维素酶菌群为乳酸菌和放线菌。HATEFIA等[18]从鞘翅目(Osphranteria coerulescens)昆虫的消化道中筛选一株高产纤维素酶菌枯草芽孢杆菌(Bacillussubtilis)。
3 结论
从西岭山原始森林保护区土壤中筛选到1株高产纤维素酶细菌菌株,鉴定其为伯克霍尔德氏菌。产酶条件优化结果表明其最佳碳源为麦麸,氮源为酵母粉,接种量为2%(V/V),初始pH 7.0,产酶时间为48 h,在最适条件下CMCase酶活最高可达2.76 U/mL。酶学特性研究显示,在pH5.0,温度60℃条件下CMCase酶活力最高。伯克霍尔德菌属菌株为西岭山中降解纤维素的优势菌群。菌株XI31-1为高产纤维素酶菌株,为丰富产纤维素酶菌株库提供价值。
参考文献:
[1]KUHAD RC,DESWAL D,SHARMA S,et al.Revisiting cellulase production and redefining current strategies based on major challenges[J].Renew Sust Energ Rev,2016,55:249-272.
[2]YANG W,MENGF,PENG J,et al.Isolation and identification of a cellulolytic bacterium from the Tibetan pig‘s intestine and investigation of itscellulaseproduction[J].Electron J Biotechnol,2014,17(6):262-267.
[3]LYNDL R,WEIMERPJ,VAN ZYL W H,et al.Microbial celluloseutilization:fundamentals and biotechnology[J].Microbiol Mol Biol Rev,2002,66(3):506-577.
[4]MOHAPATRA B R.Kinetic and thermodynamic properties of alginate lyase and cellulase co-produced by Exiguobacterium species Alg-S5[J].Int J Biol Macromol,2017,98(Supplement C):103-110.
[5]OBENGEM,BUDIMANC,ONGKUDONCM.Identifyingadditivesfor cellulaseenhancement-A systematic approach[J].Biocatal Agr Biotechnol,2017,11(Supplement C):67-74.
[6]HASHIMOTOH.Recent structural studiesof carbohydrate-binding modules[J].Cell Mol Life Sci,2006,63(24):2954-2967.
[7]ICHIKAWA S,YOSHIDA M,KARITA S,et al.Carbohydrate-binding modules influence substrate specificity of an endoglucanase from Clostridium thermocellum[J].Biosci Biotechnol Biochem,2015,80(1):188-192.
[8]BEHERA B C,SETHIB K,MISHRA RR,et al.Microbial cellulases-diversity&biotechnology with reference to mangrove environment:A review[J].J Genet Eng Biotechnol,2017,15(1):197-210.
[9]陈忠翔,房志家,陈 婷,等.一种简单高效的酵母单菌落PCR方法[J].生物技术通讯,2013(2):225-229.
[10]GHOSEK T.Measurement of cellulase activity[J].Pure Appl Chem,1987,59(2):257-268.
[11]RAWAT R T L.Purification and characterization of an acidothermophilic cellulaseenzymeproduced by Bacillussubtilis strain LFS3[J].Extremophiles,2012,4(16):637-644.
[12]FENG Y D C J,PANG H,ET AL.Cloning and identification of novel cellulase genes from uncultured microorganisms in rabbit cecum and characterization of theexpressed cellulases[J].Appl Microbiol Biotechnol,2007,2(75):319-328.
[13]VARDAVKIS E.Seasonal fluctuations of aerobic cellulolytic bacteria,and cellulase and respiratory activities in a soil profileunder a forest[J].Plant and Soil,1989,115:145-150.
[14]YANG JK,ZHANG JJ,YU H Y,et al.Community composition and cellulase activity of cellulolytic bacteria from forest soils planted with broad-leaved deciduousand evergreentrees[J].Appl Microbiol Biotechnol,2014,98(3):1449-1458.
[15]AVELLANEDATORRESL M,PULIDO CP,ROJASE T.Assessment of cellulolytic microorganisms in soils of Nevados park,Colombia[J].Brazi J Microbiol,2014,45(4):1211-1220.
[16]SHAJAHAN S,MOORTHY I G,SIVAKUMAR N,et al.Statistical modeling and optimization of cellulase production by Bacillus licheniformis NCIM 5556 isolated from the hot spring,Maharashtra,India[J].J King Saud U-Sci,2017,29(3):302-310.
[17]MAKIM L,BROEREM,LEUNG K T,et al.Characterization of some efficient cellulase producing bacteria isolated from paper mill sludges and organic fertilizers[J].Int J Biochem Mol Biol,2011,2(2):146-154.
[18]HATEFIA,MAKHDOUMIA,ASOODEH A,et al.Characterization of abi-functional cellulaseproduced by agut bacterial resident of Rosaceae branch borer beetle,Osphranteria coerulescens(Coleoptera:Cerambycidae)[J].Int J Biol Macromol,2017,103(Supplement C):158-164.
