不同供氮条件下加工番茄化感抑制龙葵的作用
2018-05-10李文杰崔家丽王巧丽王俊刚
李文杰, 刘 璐, 崔家丽, 钟 亮, 王巧丽, 王俊刚
(石河子大学农学院,新疆石河子 832003)
新疆地区具有良好的光热资源,是加工番茄的优秀生产基地,其种植与加工规模、年出口量居全国之首,全疆2014年出口的番茄酱量增质升。全疆加工番茄种植面积达7.5×103hm2,产量7.8×105t,出口报检量目前已近5×105t,10年来出口抽检合格率首次达到100%[1]。随着加工番茄的种植,杂草种类及其危害也随着变化,龙葵成为加工番茄的主要草害之一,其量大、危害性强,与番茄竞争阳光、水分、氮(N)、磷(P)等能量与营养元素,消耗了土壤中的大量营养物质,尤其是氮元素。在植物对营养物质、水、光热资源竞争的研究中,氮素营养、施用量得到了深入的研究,有报道指出,土壤中的氮素在作物产量上的贡献率约为48.6%~79.4%,但氮肥的利用率为28%~41%,相对较低[2]。同样,氮素对加工番茄的产量也有着不可忽视的作用[3-4]。关于是化感作用还是资源竞争引起了植物化感作用现已成为学者争论、研究的焦点,并希望能够找出明确的植物种间资源竞争的方法[5-12]。从Dilday等发明了田间抑制圈[8]开始,到现在的迟播、共培法,学者们正不断朝着克服资源竞争干扰、优化生物测试的方向努力。种间竞争和化感作用仍包含在田间抑制圈生物测试法所获得的结果中。Weidenhamer的竞争-化感分离法[9]对迟播共培法进行了弥补。新疆加工番茄在不同氮条件下的化感作用潜力变化目前尚缺乏相关研究,不同氮条件下的化感作用潜力变化对龙葵的竞争情况也不甚明了,因此分析引起化感作用潜力的原因,证实氮胁迫过程中的生理、生化机制等问题都亟待解决。本研究采用化感-竞争分离(ACS)生物测试法来研究龙葵在不同供氮水平下受加工番茄的化感作用,以期为生产实践提供一定的理论依据。
1 材料与方法
1.1 供试材料
加工番茄品种为硕源87-5,由北京硕源农业科技有限公司生产,购自石河子种子市场;龙葵采自石河子大学农学院试验站(2013年收获后保存于4 ℃冰箱)。
1.2 试验设计
1.2.1 试验方案1 将催好芽的加工番茄、龙葵分别播在已经覆土的秧盘里培养。挑选2叶1心期的2种植株移至盛有3 L完全相同培养液的塑料盆(45 cm×35 cm×15 cm)内培养,塑料盆内悬浮厚度为1.5 cm的泡沫板,上面均匀分布35个直径为2 cm小孔,待其恢复6 d,将加工番茄和龙葵分别移植于泡沫板上的孔中并用棉花固定(每孔1株,每盆30株加工番茄+5株龙葵,龙葵集中在中央1排),测试在共培条件下加工番茄对龙葵化感抑制作用潜力的变化。营养液组分为NH4NO3、K2SO4、KH2PO4,分3个氮水平(0.8、1.6、3.3 mg/L,分别计作N1、N2、N3或1/4N、1/2N、N),只含有氮、磷、钾(K)3种大量元素(磷、钾含量分别为12.41、0.73 mg/L);对照组不施营养液。每7 d对各处理的龙葵取样、烘干、称质量并更换1次营养液,试验重复3次,直到第21天。计算抑制作用率(inhibitory effect rate,简称IR):
IR=[(m处理-m对照)/m对照]×100%。
式中:IR>0表示有促进作用,其值为促进率;IR<0表示有抑制作用,取其绝对值,表示抑制率[13]。
1.2.2 试验方案2 将上述培养7 d后的培养液倒出,测定其中剩余的氮、磷、钾浓度,并将培养液补足到正常浓度时的氮、磷、钾水平,保证各组相对一致。此外用蒸馏水将NH4NO3、K2SO4和KH2PO4配制成正常营养水平的氮、磷、钾浓度作为另外一个对照(对照所用的培养液与加工番茄/龙葵培养液的区别仅在于,对照培养液中没有加工番茄与龙葵共培养时释放的物质,而加工番茄/龙葵培养液中含有加工番茄与龙葵共培7 d后释放的物质)。移植9株3叶1心期的龙葵到上述每种培养液中培养,每7 d分别取样、烘干、称质量并计算对龙葵的抑制率及培养液中磷、钾含量,直到第21天,每个处理3次重复。
1.3 数据处理与统计
采用SPSS软件进行数据统计分析。
2 结果与分析
2.1 不同共培条件对龙葵干质量及其抑制率的影响
由表1可知,第7、14天,随着原营养液氮处理水平的增加,龙葵干物质量增加,直到第21天开始降低;第7天与第21天,3种处理液中培养的龙葵干物质量间差异不显著,第14天3种处理间存在显著差异。表2表明:龙葵抑制率最高为73.00%,随着原营养液氮处理水平的增加,抑制率整体上持续降低,在第14天抑制率在处理2水平下(即N2水平)达到最高值73.04%。第14天与第21天3种处理液中部分龙葵干物质量间差异不显著,与第7天相比多数处理间差异显著,表明低氮条件下加工番茄对龙葵有较高的抑制率,但在中、高氮条件下抑制率相对较低。
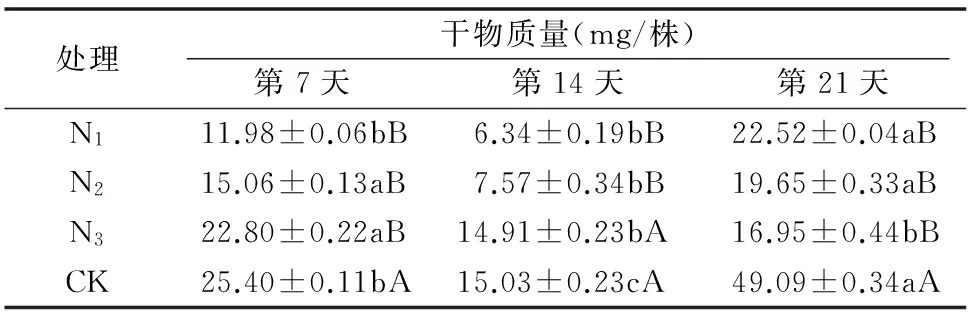
表1不同共培条件下加工番茄对龙葵干物质量的影响
注:同行数据后不同小写字母表示差异显著(P<0.05),同列数据后不同大写字母表示差异显著(P<0.05)。下表同。
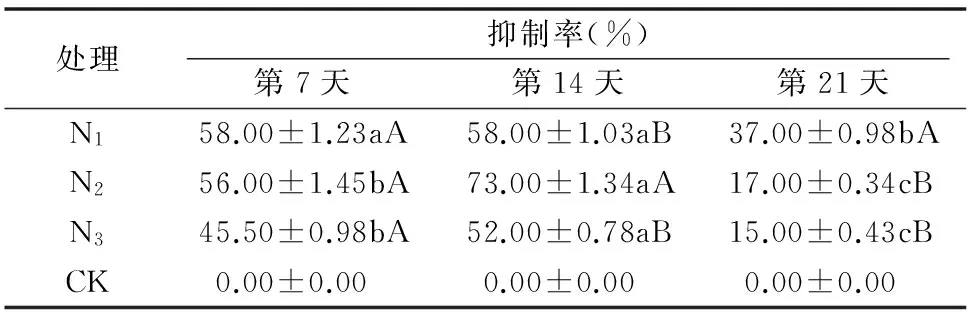
表2不同共培条件下加工番茄对龙葵抑制率的影响
2.2 加工番茄化感与龙葵种间竞争能力比较
2.2.1 加工番茄培养液对龙葵干物质量及其抑制率的影响 为了提高评价加工番茄化感抑制龙葵能力的准确性,降低各培养液之间的营养差异,对以上经过7 d共培的培养液进行浓度调节,直到各培养液氮、磷、钾浓度恢复到相对一致的水平。表3、表4表明,在经过营养调节后,各营养水平的差异得到消除,但仍表现出与表1、表2一致的趋势,即龙葵干物质量最低时,抑制率最高,并随着原营养液氮处理水平的增加,干物质量增加,而抑制率下降。
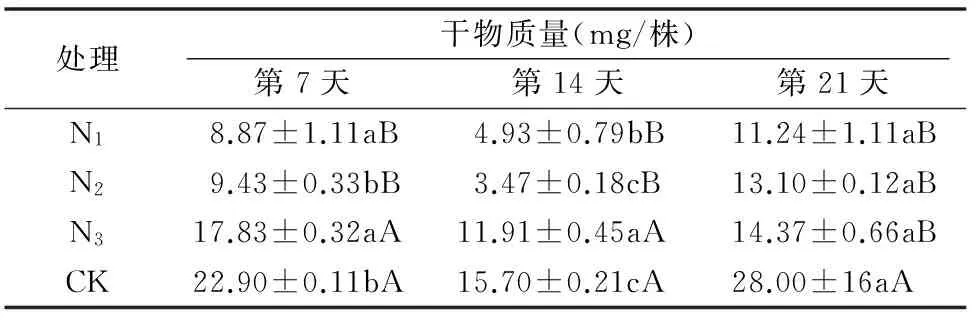
表3加工番茄培养液对龙葵干物质量的影响
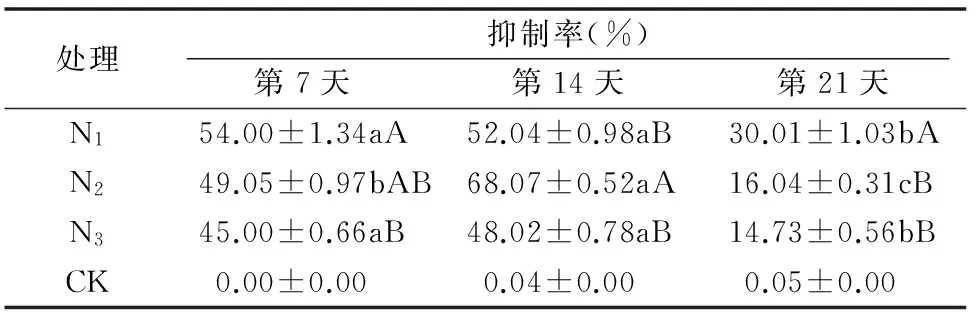
表4加工番茄培养液对龙葵抑制率的影响
2.2.2 加工番茄化感作用与种间竞争能力比较 图1、图2表明,加工番茄对龙葵具有化感抑制能力,在低氮胁迫条件下,其化感潜力高于氮素水平高的处理,并且有明显的增强趋势。如图3所示,在不同处理条件下,加工番茄对龙葵的化感作用变化情况(A)与生物干扰曲线(A+R)成功地分离、区别了植物化感作用与资源竞争的生物干扰现象。由此可以看出,因生物干扰(资源竞争+化感作用)在营养资源正常偏丰富的条件下对龙葵的相对抑制率为1%~17%和0.2%~4%,表现为化感作用大于资源竞争。在低氮胁迫下,因加工番茄资源竞争和化感作用导致对共生龙葵的抑制率为4%~7%。由此可见,加工番茄不但有资源竞争能力,而且具有化感抑制龙葵的能力。
2.3 加工番茄与龙葵对营养物质的竞争分析
对加工番茄与龙葵共培第7天培养液中磷、钾含量进行测定并比较分析。表5结果表明,培养液中钾含量最低为 0.09 μg/mL,在不同氮水平下,第7天与其他时间的钾含量差异显著;第14天N2与N3水平下钾含量差异不显著,与N1水平钾含量差异显著;第21天N1与N2水平下钾含量差异不显著,与N3水平钾含量差异显著。由此看出,在不同时间,培养液中钾含量均表现为显著差异。
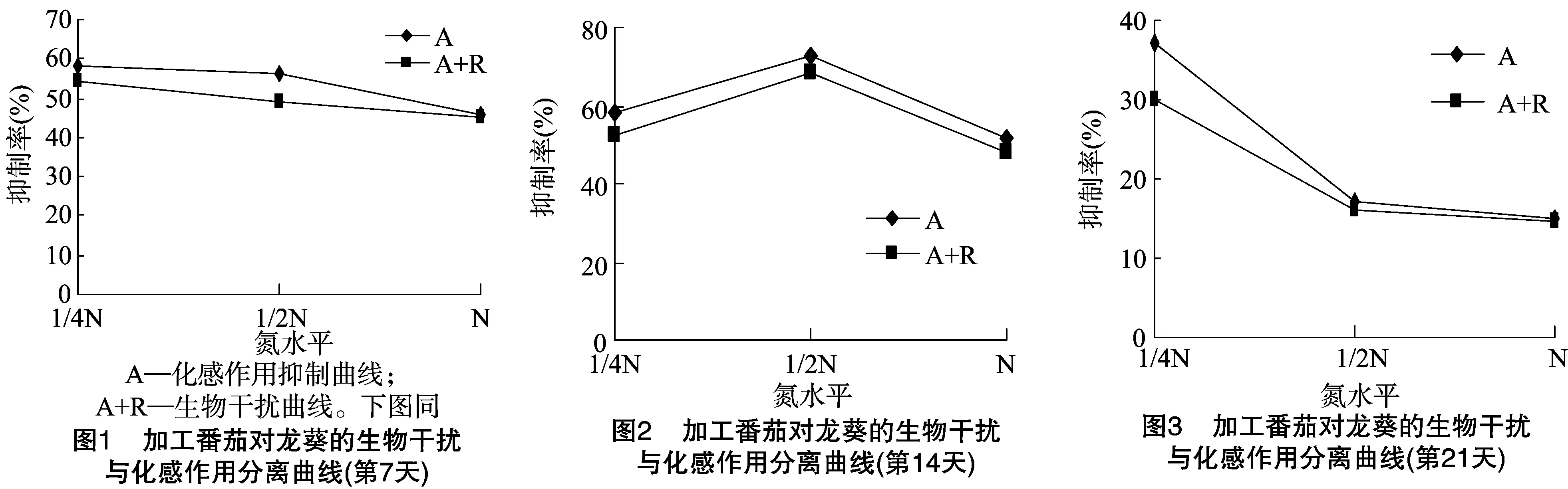
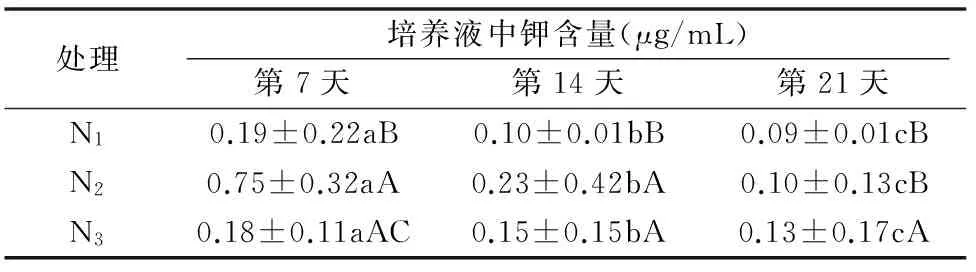
表5加工番茄对营养物质钾的竞争比较
表6结果表明,培养液中磷含量最低为10.45 μg/mL,在不同氮水平下,第7天与其他时间的磷含量差异显著;第14天,N1与N3水平下的磷含量差异不显著,与N2水平的磷含量差异基本表现为显著;第21天与其他时间的磷含量差异整体显著。在不同时间下,N1水平下的磷含量存在显著差异;N2水平下,第7天与第14、21天的磷含量差异显著;N3水平下的磷含量与其他水平差异显著。

表6不同共培条件下加工番茄对营养物质磷的竞争比较
3 结论
本试验结果表明,在加工番茄与龙葵共生系统中,加工番茄随氮营养水平的下降,在第14天的抑制率在N2水平下达到最高值73.00%,对龙葵施加的生物干扰逐渐增大,使龙葵生长受阻,从而导致抑制率也提高。生物干扰(资源竞争+化感作用)在营养资源正常偏丰富的条件下导致对龙葵的抑制率为1%~17%和0.2%~4%,表现为化感作用大于资源竞争。在低氮胁迫下,因加工番茄资源竞争和化感作用导致对共生龙葵的抑制率为4%~7%。在加工番茄与龙葵共生系统中,加工番茄在氮胁迫(资源短缺)下,主要通过提高化感作用[14-16]来抑制龙葵生长。当资源短缺(如N1条件)时加剧了生态位竞争,表明加工番茄对其伴生杂草抑制作用增大,相应的抑制率提高。对于加工番茄而言,它具有化感作用,但竞争行为有所不同,无论外在环境资源丰欠与否,均表现出生物干扰能力。
4 讨论
氮作为参与植物体内各种代谢活动的生命元素,对加工番茄的生长、发育及产量的影响深刻而复杂。根据供试土壤的养分状况,加工番茄干物质及氮、磷、钾养分的积累动态均呈“S”形增长,可以通过增加施氮量来促进氮、磷、钾的吸收,各施氮处理比不施氮处理多吸收75.8%~189.2%氮,49.4%~162.7%磷,60.7%~176.8%钾。齐红岩等的研究表明,土壤中氮素对作物产量作出48.6%~79.4%的贡献,加工番茄与龙葵共生系统中,随着氮营养水平的下降,加工番茄对龙葵生物干扰也逐渐加大,使龙葵生长受阻,进而对龙葵抑制率提高[2]。是否因为氮素的变化激发了番茄对龙葵抑制的信号,以及氮素的改变对番茄抑制的分子机制尚不可知。
目前通过合理密植作物,增加田间杂草与作物共生系统的竞争,从而抑制杂草的生长,这种新型的杂草生物防治方式正在摸索中,使得新型的杂草防治模式成为可能。众所周知,龙葵不仅是杂草更是一味中药材,在杂草与作物的共生系统中,龙葵是否分泌了特殊的物质从而刺激了番茄的化感作用,从而引起在低氮胁迫下番茄对龙葵的抑制能力加强,还有待继续探究。
参考文献:
[1]齐士发,石 强,王新燕,等. 新疆加工番茄产业原料生产中存在的问题与对策[J]. 农业科技通讯,2018(11):81-85.
[2]齐红岩,李天来,富宏丹,等. 不同氮钾施用水平对番茄营养吸收和土壤养分变化的影响[J]. 土壤通报,2006,37(2):268-272.
[3]汤明尧,张 炎,胡 伟,等. 不同施氮水平对加工番茄养分吸收、分配及产量的影响[J]. 植物营养与肥料学报,2010,16(5):1238-1245.
[4]王 新,马富裕,刁 明,等. 不同施氮水平下加工番茄植株生长和氮素积累与利用率的动态模拟[J]. 应用生态学报,2014,25(4):1043-1050.
[5]徐 涛,周 强,夏 嫱,等. 水稻挥发物对褐飞虱行为的影响[J]. 科学通报,2002,47(11):849-853.
[6]熊 君,林文雄,周军建,等. 不同供氮条件下水稻的化感抑草用与资源竞争分析[J]. 应用生态学报,2005,16(5):885-889.
[7]何华勤,董章杭,梁义元,等. 水稻化感作用研究的新进展[J]. 农业现代化研究,2002,23(2):140-143.
[8]Dilday R H,Mattice J D,Moldemhauer K R.An overview of rice allelopathy in the USA[M]//Kim K U,Shin D H.Rice allelopathy.Korea:Chan Suk Park Publish,2000:15-26.
[9]Weidenhamer J D.Distinguishing resource competition and chemical interference:overcoming the methodological impasse[J]. Agron J,1996,88(6):866-875.
[10]黄云霄,齐 勇,杨晓娜,等. 龙葵提取物对不同作物幼苗的化感效应[J]. 杂草学报,2017,35(2):40-45.
[11]McKey D,Waterman P G,Mbi C N,et al.Phenolic content of vegetation in two African rain forests:ecological applications[J]. Science,1978,202(4363):61-64.
[12]朱 峰,何永福,叶照春. 大叶芥菜对眼子菜化感作用潜力的初步评价[J]. 杂草学报,2016,34(2):49-52.
[13]Gershenzon J. Changes in the levels of plant secondary metabolites under water and nutrient stress[J]. Rec Adv Phytochem,1984,18:273-321.
[14]Granéli E,Johansson N.Increase in the production of allelopathic substances byPrymnesiumparvumcells grown under N- or P-deficient conditions[J]. Harmful Algae,2003,2(2):135-145.
[15]Einhellig F A. Allelopathy:current status and future goals[J]. American Chemical Society Symposium Series,1995,58:1-24.
[16]Olofsdotter M. Getting closer to breeding for competitive ability and the role of allelopathy-an example from rice(Oryzasativa)[J]. Weed Technol,2001,15(4):798-806.
