西伯利亚杏种质资源花表型变异
2018-05-08尹明宇朱绪春刘慧敏柳江群乌云塔娜
尹明宇,朱绪春,刘慧敏,柳江群,乌云塔娜
(中国林业科学研究院 经济林研究开发中心,河南 郑州450003)
西伯利亚杏(Armeniacasibirica(L.)Lam)为蔷薇科(Rosaceae)杏属(ArmeniacaMill.)植物[1],主要分布在中国、俄罗斯、蒙古国[2],在我国主要分布于内蒙古、辽宁地区,多为野生或半野生状态,集中成片生长。西伯利亚杏耐干旱、瘠薄,适应性强,作为先锋树种广泛应用于“三北”地区沙漠防护林营造;西伯利亚杏的杏仁粗脂肪含量45.64%,粗蛋白含量28%,苦杏仁苷含量4.5%[3],故其是重要的木本油料和粮食兼用树种。
种质资源评价是资源合理利用的前提,数量性状的合理分级是种质资源评价的基础[4]。2003年出版的《中国果树志:杏卷》[5]对杏的各性状在经验基础上进行了等差分级,但未能反映出性状取值的概率分布情况。2006年出版的《杏种质资源描述规范和数据标准》[6],提出了部分项目的参照品种,但依然没有关于西伯利亚杏的花性状数量分级标准。西伯利亚杏自交不亲和[7],生产上常需要合理配置授粉树或者在花期人工辅助授粉,以确保获得预期的产量和品质。花粉量的多少和花粉生活力的高低直接影响授粉、受精乃至座果,开展花粉量和生活力的研究对种质评价及杂交育种工作有重要意义。生产上杏树普遍存在自然坐果率低、产量不稳定的问题。虽然自交不亲和是西伯利亚杏自然结实率低的主要原因,但雌蕊败育现象在西伯利亚杏中也普遍存在[8],雌蕊败育的花珠被发育不完全,雌蕊发育滞缓,花期子房、花柱短小,不能形成成熟的胚囊,最终导致落花。雌蕊败育花的多少与品种、成熟期、花芽在结果枝上的着生部位、树木生长的立地条件和营养状况有关[9-10]。目前,关于西伯利亚杏生殖生物学研究主要集中在自交不亲和性和S型基因的分离及鉴定[11-12]、花粉形态及孢粉学特征[13]等方面,而有关花表型性状的数量分级、花粉量及其萌发率的系统分类、雌蕊败育规律的研究鲜有报道。本研究对西伯利亚杏的花表型性状、花粉特性、雌蕊发育程度以及彼此间的相关性进行调查,了解花表型变异规律以及花粉和雌蕊特性,针对数量性状予以概率分级,对质量性状进行概率分布描述,以期为西伯利亚杏的种质资源评价和品种选育提供参考。
1 材料与方法
1.1 试验材料及样本采集
试验材料取自内蒙古林木良种繁育中心收集保存的西伯利亚杏资源。结合多年田间观察和试验,从13个种源地一共筛选出256份有代表性的、树龄均在8年以上的种质资源为材料,这13个种源地(种质资源数量)分别为和林格尔县(38个)、扎鲁特旗(25个)、敖汉旗(21个)、巴林右旗(18个) 、阿鲁科尔沁旗(17个)、万家沟(20个)、克什克腾旗(13个)、凉城(28个)、扎赉特旗(15个)、科右中旗(17个)、乌拉山(9个)、乌兰浩特(17个)、科左后旗(18个)。试验于2015年4-5月进行。在盛花期,于每个调查株的东、南、西、北4个方向随机选取无病虫危害的健康花朵,调查花性状、收集花粉并统计坐果率等。试验地内蒙古土默特左旗4月气温变化以及西伯利亚杏盛花期的分布如图1所示。当气温达到10.3 ℃西伯利亚杏开始开花[5],试验地4月17日平均气温达到12 ℃,从而进入花期;从4月20日起平均气温都高于10.5 ℃,开花频率高于13.5%,进入盛花期,盛花期集中分布在4月20日至4月24日(图1)。
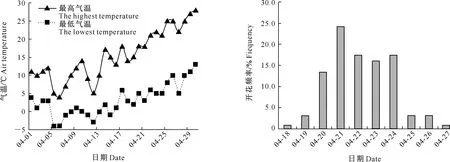
图1 内蒙古土默特左旗4月份气温变化以及西伯利亚杏的花期分布Fig.1 Air temperature variation in Tumote Zuoqi,Inner Mongol and bloom stage of Armeniaca sibiric in April
1.2 方 法
1.2.1 花性状调查和雌蕊发育程度统计 参照《中国果树志:杏卷》[5]和《杏种质资源描述规范和数据标准》[6],共选取了西伯利亚杏花的花长、花冠径、花瓣长、花瓣宽、雄蕊个数、雄蕊长、雌蕊长、花粉量、花粉活力9个数量性状以及花瓣颜色、花萼颜色、花瓣形状、花瓣状态、花瓣类别5个质量性状进行调查。其中,花长、花冠径、花瓣长、花瓣宽、雄蕊长、雌蕊长用游标卡尺测量,游标卡尺读数精确到0.01 mm。雄蕊长测量时的起点与雌蕊相同;测量雌蕊长度时,将雌蕊退化标记为0。对质量性状的调查,按性状示意图(图2)进行。根据雄蕊和雌蕊长,统计雌蕊发育程度,雌蕊发育类型分为:Ⅰ.雌蕊长于雄蕊,Ⅱ.雌蕊与雄蕊等长,Ⅲ.雌蕊短于雄蕊,Ⅳ.雌蕊极短或变黑退化,其中Ⅰ、Ⅱ为完全花,Ⅲ、Ⅳ为雌蕊败育。于盛花期对每株丰产优株在东、西、南、北4个方向计数不少于100朵的总花数,统计4种类型雌蕊个数,并计算完全花比例。在样木上按开花状况选取标准枝,盛花期统计枝上花数量,并对果枝进行标记,花谢10 d后调查坐果率。选择有代表性的健康单株,分东、西、南、北、中5个方位,每个方位又分上、中、下3个层次,测量枝条枝长、枝径,统计完全花数、败育花数、总花数和坐果数,计算完全花比例、坐果率。
完全花比例=完全花数/调查总花数×100%;
坐果率=坐果数/调查总花数×100%。
1.2.2 花粉量测定 在开花前1 d采集健康的花蕾,取完整花药15枚放入硫酸纸盒子中室内晾干,待花药完全爆裂散出花粉后,转入1.5 mL离心管中,加入质量分数1%纤维素酶溶液1 mL处理24 h,使花粉粒均匀分布于溶液中。然后取1 μL溶液滴于凹面载玻片上,在显微镜下观察统计花粉粒数,重复3次[14]。
单朵花花粉量=单朵花花药总数(按每朵花平均26枚花药进行计算)×1 μL溶液花粉数平均值×1 000/15。
1.2.3 花粉活力检测 采用萌发试验检测花粉活力[15-16],培养基为1%琼脂+10%蔗糖+0.01%硼酸+88.99%。具体方法是:取适量加热融化的培养基液滴于凹面载玻片上,将少量刚刚收集的花粉均匀播撒于培养基液表面,置于25 ℃恒温培养箱中培养。4 h后在显微镜下观察花粉的萌发状况。每样品做3个重复玻片,每玻片取3个视野,选取花粉粒数目不少于50粒且分布均匀的视野进行统计。计数时以花粉管长度超过花粉粒直径作为萌发标准。花粉萌发率的计算公式如下。
A=(x/y)×100%。
式中:A表示花粉萌发率,x表示视野中已萌发花粉粒数,y表示视野中花粉粒总数。
1.3 数据处理
对数量性状利用Excel 2007软件和SPSS 20.0软件进行数据整理与分析,分别统计均值、标准差、极小值、极大值、极差、变异系数等,并以此分析不同西伯利亚杏种质资源花的表型变异和多样性。对花的9个数量性状进行K-S检验[17]。对花7个表型性状(花长、花冠径、花瓣长、花瓣宽、雄蕊个数、雄蕊长、雌蕊长)与花粉特性(花粉量、花粉活力)以及雌蕊发育程度(完全花比例)之间进行相关性分析。最后对完全花数、败育花数、完全花比例与枝条方位、枝条层次、枝长、枝径、总花数、坐果数、坐果率的相关性进行分析。

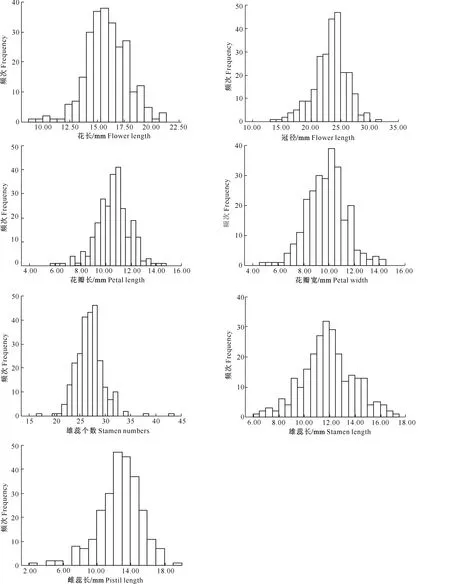
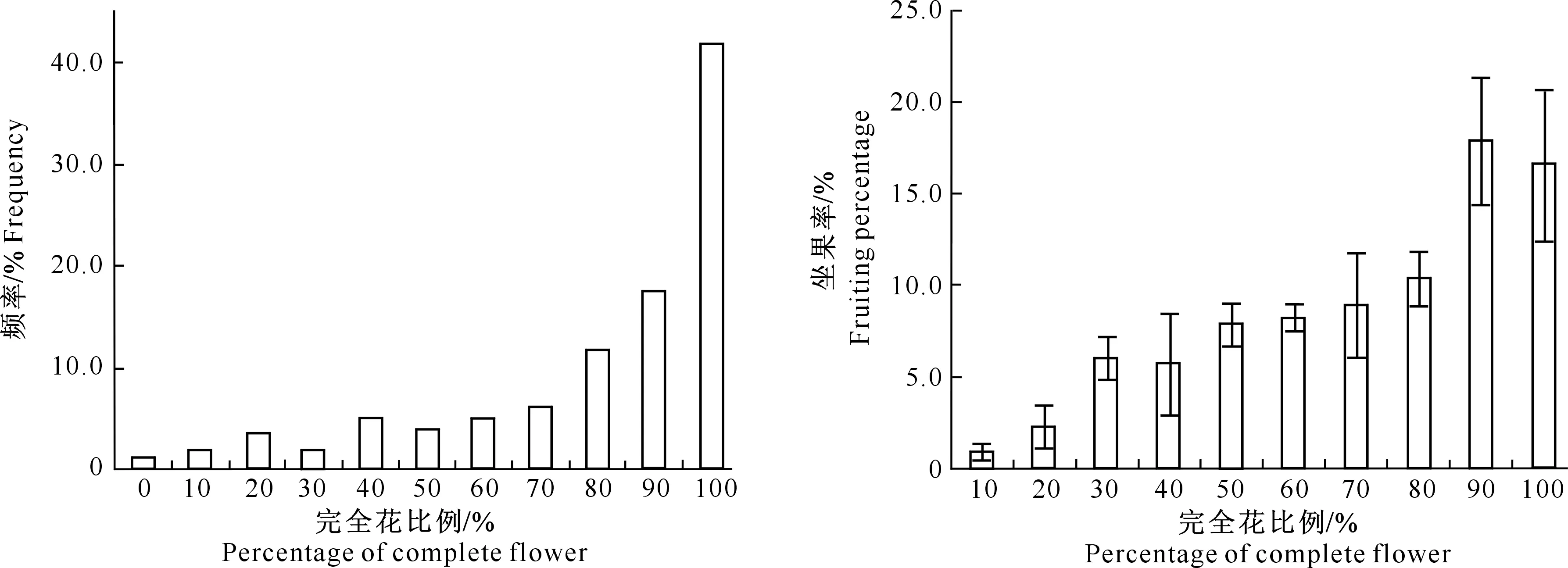
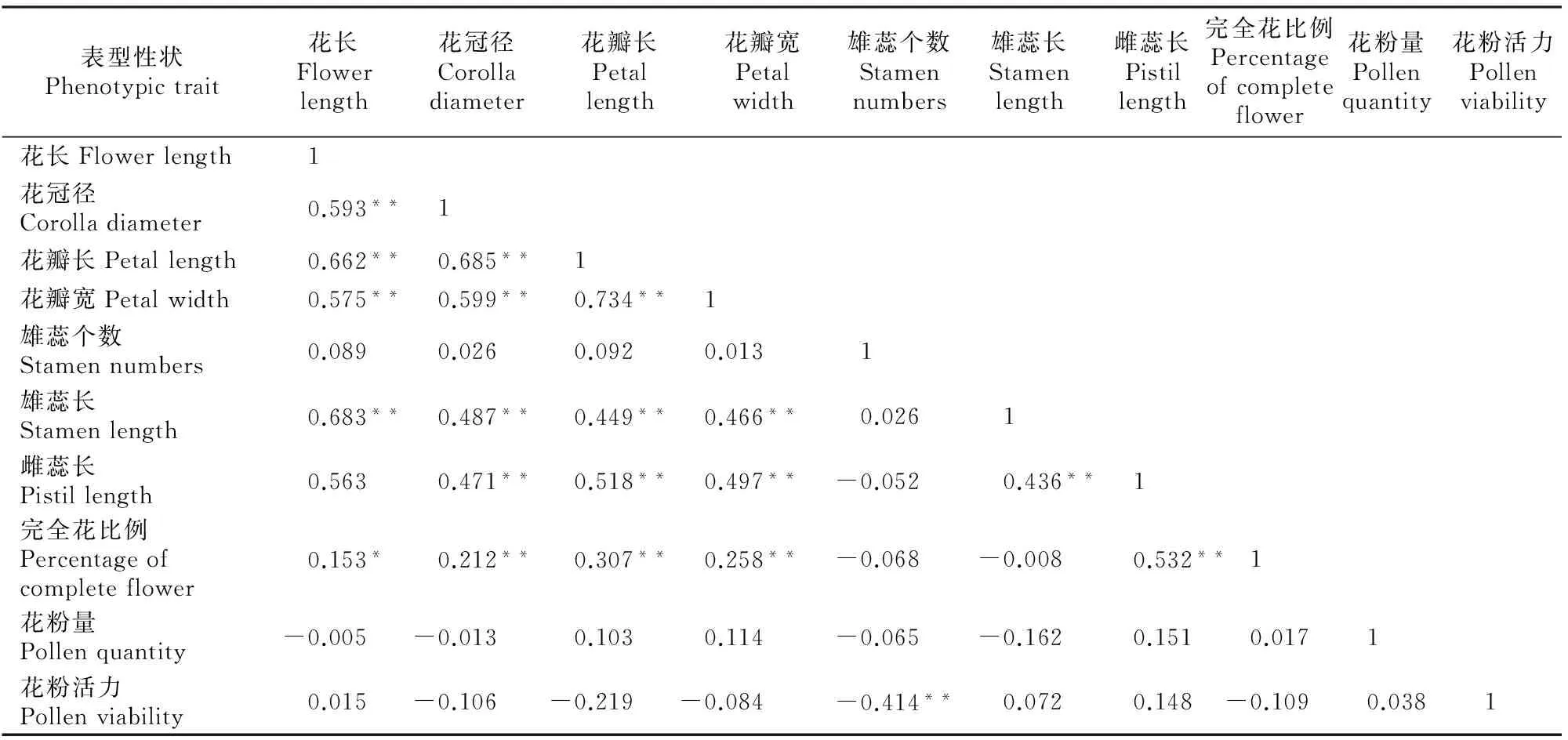
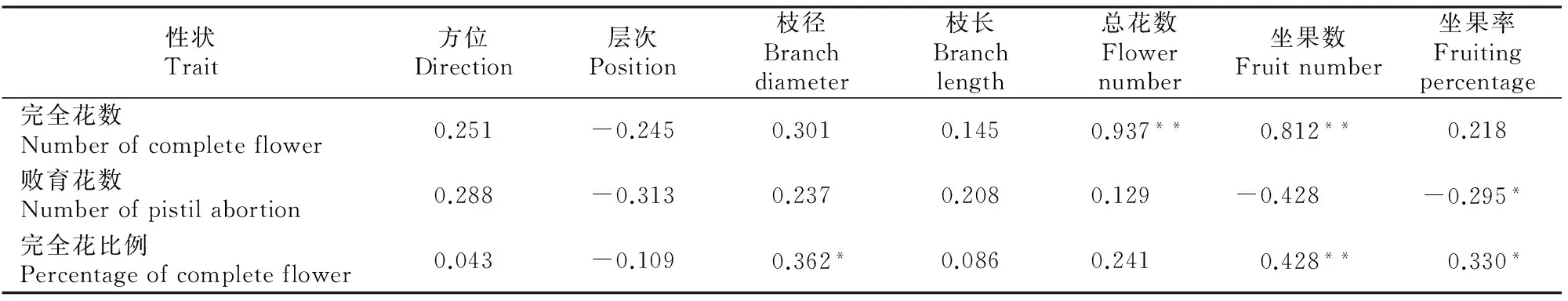
A~D.花颜色示意图:A.白色,B.浅粉色,C.深粉色,D.红色;E~H.花萼颜色示意图,E.红褐色,F.紫红色,G.紫绿色,H.绿色;I~L.花瓣形状 示意图:I.圆形,J.卵圆形,K.椭圆形,L.长圆形;M~N.花瓣状态示意图:M.平,N.皱;O.重瓣花;P.花瓣、雄蕊缺失,雌蕊15~20个;Q~T.雌蕊发育程度示意图:Q.雌蕊长于雄蕊,R.雌蕊与雄蕊等长,S.雌蕊短于雄蕊,T.雌蕊极短或变黑退化,Q~T中箭头指示柱头的位置 A-D.The schematic diagram of petal color:A.White,B.Light pink,C.Deep pink,D.Red;E-H.The schematic diagram of calyx color: E.Brown red,F.Purple,G.Purplish red,H.Green;I-L.The schematic diagram of petal shape:I.Round,J.Ovate,K.Elliptic,L.Oblong;M-N.Petal smoothness:M.Smooth,N.Rough;O.Multiplicate;P.Petals and stamen abortion,pistil 15-20;Q-T.The schematic diagram of pistil development degree:Q.Pistil>stamen,R.Pistil=stamen,S.Pistil 西伯利亚杏花的数量性状特征见表1。由表1可见,西伯利亚杏花的数量性状变异系数在10.26%~41.41%,平均值为19.77%,其中花粉量的变异系数最大(41.41%),变异幅度为9 533.00~55 467.00;其次为花粉活力的变异系数(36.42%),变异幅度为15.89%~85.12%;雄蕊个数的变异系数最小(10.26%),变异幅度为17.00~43.00。与生殖相关的花粉量和花粉活力相对于花表型性状变异更大,具有更加丰富的多样性。西伯利亚杏花的数量性状变异类型多且差异较大,多样性比较丰富。西伯利亚杏花主要性状特征占样本总量的比例见图3。 表1 西伯利亚杏花数量性状的多样性Table 1 Diversity in quantitative traits of Armeniaca sibirica flower 图3 西伯利亚杏花的质量性状及雌蕊发育类型的分布Fig.3 Distribution of quality traits and pistil development degree of Armeniaca sibirica 西伯利亚杏主要性状描述特征为:花瓣颜色为白色(56.13%)或浅粉红色(43.23%);花瓣形状为圆形(62.26%)或卵圆形(26.45%);花萼颜色为红褐色(58.06%)或紫红色(34.52%);花瓣状态平(71.29%);花单瓣(99.35%)(图3)。 西伯利亚杏花的9个数量性状的K-S检验见表2,分布频次见图4和图5。K-S检验结果(表2)显示:花长、花冠径、花瓣长、花瓣宽、雄蕊长、雌蕊长、花粉量、花粉活力等8个性状的Sig.值均大于0.05,从而符合正态分布。雄蕊个数的Sig.值小于0.05,不完全符合正态分布,但仍可近似当做正态分布处理。对符合正态分布性状,用(X-1.281 8S)、(X-0.524 6S)、(X+0.524 6S)和(X+1.281 8S)4个分点值分为5个等级,其中X代表各性状的平均值,S代表各性状的标准差,使1~5级的出现概率分别为10%,20%,40%,20%和10%,将数量性状进行统一化和规范化的描述。不同数量性状的4个分点值见表2。 表2 西伯利亚杏花数量性状的K-S检验及分级Table 2 K-S normal test and probability grading of quantitative characters of Armeniaca sibirica flower 2.3.1 花 长 西伯利亚杏花花长变异幅度为9.29~21.30 mm,平均15.87 mm,变异系数12.19%(表1)。花长概率分级的频率分布: 1级(<13.39 mm)占8.21%,2级(13.39~14.86 mm)占20.78%,3级(14.86~16.89 mm)占41.18%,4级(16.89~18.35 mm)占20.78%,5级(>18.35 mm)占9.02%。 2.3.2 花冠径 西伯利亚杏花花冠径变异幅度为13.98~31.80 mm,平均23.28 mm,变异系数12.32%(表1)。花冠径概率分级的频率分布:1级(<19.61 mm)占10.59%,2级(19.61~21.78 mm)占17.25%,3级(21.78~24.79 mm)占44.31%,4级(24.79~26.96 mm)占19.61%,5级(>26.96 mm)占8.24%。 2.3.3 花瓣长 西伯利亚杏花花瓣长变异幅度为5.97~14.65 mm,平均10.63 mm,变异系数12.43%(表1)。花瓣长概率分级的频率分布:1级(<8.93 mm)占8.63%,2级(8.93~9.93 mm)占19.61%,3级(9.93~11.32 mm)占43.53%,4级(11.32~12.32 mm)占19.61%,5级(>12.32 mm)占8.63%。 2.3.4 花瓣宽 西伯利亚杏花花瓣宽变异幅度为4.78~14.01 mm,平均9.84 mm,变异系数15.67%(表1)。花瓣宽概率分级的频率分布:1级(<7.86 mm)占10.20%,2级(7.86~9.03 mm)占19.61%,3级(9.03~10.64 mm)占41.18%,4级(10.64~11.81 mm)占19.22%,5级(>11.81 mm)占9.80%。 2.3.5 雄蕊个数 西伯利亚杏花雄蕊个数变异幅度为17.00~43.00,平均26.95,变异系数10.26%(表1)。雄蕊个数概率分级的频率分布:1级(<23个)占8.24%,2级(23~26个)占14.12%,3级(26~28个)占52.94%,4级(28~30个)占16.47%,5级(>30个)占8.24%。 2.3.6 雄蕊长 西伯利亚杏花雄蕊长变异幅度为6.37~17.29 mm,平均11.90 mm,变异系数17.08%(表1)。雄蕊长概率分级的频率分布:1级(<9.29 mm)占9.80%,2级(9.29~10.83 mm)占17.65%,3级(10.83~12.96 mm)占45.88%,4级(12.96~14.50 mm)占15.69%,5级(>14.50 mm占)10.98%。 2.3.7 雌蕊长 西伯利亚杏花雌蕊长变异幅度为0.00~19.90 mm,平均12.69 mm,变异系数20.16%(表1)。雌蕊长概率分级的频率分布:1级(<9.41 mm)占9.02%,2级(9.41~11.35 mm)占15.69%,3级(11.35~14.03 mm)占45.49%,4级(14.03~15.97 mm)占22.35%,5级(>15.97 mm)占7.45%。 图4 西伯利亚杏花表型性状的分布频次Fig.4 Frequency distribution of phenotypic traits of Armeniaca sibirica 图5 西伯利亚杏花粉特性的分布频次Fig.5 Frequency distribution of pollen of Armeniaca sibirica 2.3.8 花粉量 西伯利亚杏花单花(26枚花药)花粉量变异幅度为9 533.00~55 467.00粒/朵,平均26 195.98粒/朵,变异系数41.41%(表1)。花粉量概率分级的频率分布:1级(<12 291.00粒/朵)占8.33%,2级(12 291.00~20 505.00粒/朵)占26.19%,3级(20 505.00~31 887.00粒/朵)占35.71%,4级(31 887.00~40 101.00粒/朵)占16.67%,5级(>40 101.00粒/朵)占13.10%。 2.3.9 花粉活力 西伯利亚杏花单花花粉活力变异幅度为15.89%~85.12%,平均47.15%,变异系数36.42%(表1)。花粉活力概率分级的频率分布:1级(<25.14%)占9.52%,2级(25.14%~38.16%)占22.62%,3级(38.16%~56.20%)占39.29%,4级(56.20%~69.21%)占14.29%,5级(>69.21%)占14.29%。其中花粉活力大于50%的样本占42.86%。 将西伯利亚杏的雌蕊发育程度分为4种类型:Ⅰ.雌蕊长于雄蕊;Ⅱ.雌蕊与雄蕊等长;Ⅲ.雌蕊短于雄蕊;Ⅳ.雌蕊退化,各类型在样本中的比例见图3。图3中,Ⅱ类型所占比例最高(57.96%),其次为Ⅰ类型(22.22%),Ⅳ类型和Ⅲ类型所占比例较低,分别为11.13%和8.78%。Ⅰ、Ⅱ为完全花,Ⅲ、Ⅳ为雌蕊败育,完全花比例的平均值为80.11%,在所有样本中的频率分布见图6。 图6 西伯利亚杏完全花比例分布及其与坐果率的关系Fig.6 Distribution of complete flower percentage of Armeniaca sibirica and the relationship with fruiting percentage 由图6可见,样本出现频率随着完全花比例的增高而增加,41.8%的西伯利亚杏完全花比例达到100%,17.6% 的西伯利亚杏完全花比例达到90%,11.7%的西伯利亚杏完全花比例达到80%,其余比例完全花的频率均低于6.3%。86.4%的西伯种亚杏完全花比例高于50%。 完全花比例与坐果率的关系见图6。由图6可见,不同比例完全花的坐果率为0.9%~17.9%,其中完全花比例为50%时坐果率为7.9%,完全花比例为90%时坐果率高达17.9%,完全花比例为100%时坐果率为16.6%。随着完全花比例的增加,坐果率总体上呈先上升后下降趋势。 完全花在方位和层次上的分布情况见表3。由表3 可见,完全花比例在方位上表现为为中>南>东>西>北,在层次上表现为中>上>下。 表3 西伯利亚杏树体不同方位和层次的雌蕊发育程度Table 3 Pistil development of Armeniaca sibirica tree in deferent directions and positions 西伯利亚杏花表型性状与花粉特性以及雌蕊发育程度与坐果率的相关性如表4和表5所示。表4表明,西伯利亚杏花7个表型性状与花粉特性间存在显著或极显著相关性。 表4 西伯利亚杏花表型性状与花粉特性的相关性Table 4 Correlations between phenotypic traits and pollen of Armeniaca sibirica 注:*与**分别表示在P<0.05和P<0.01水平上显著相关。下表同。 Note:* and ** indicate significant correlations atP<0.05 andP<0.01,respectively.The same below. 表5 西伯利亚杏雌蕊发育与坐果率的相关性Table 5 Correlations between pistil development and fruiting percentage of Armeniaca sibirica 表4显示,除花长和雌蕊长之间无显著相关关系外,花长、花冠径、花瓣长、花瓣宽、雄蕊长、雌蕊长6个表型性状间均存在极显著的正相关关系(P<0.01),而这6个性状与雄蕊个数之间均无显著相关关系;完全花比例与花冠径、花瓣长、花瓣宽、雌蕊长之间呈极显著正相关关系(P<0.01),与花长之间呈显著正相关关系(P<0.05);花粉量与花瓣长、花瓣宽、雌蕊长、完全花比例之间的相关性均不显著;花粉活力与雄蕊个数呈极显著负相关关系(P<0.01)。 表5表明,完全花数与总花数、坐果数呈极显著的正相关关系(P<0.01);败育花数与坐果率呈显著负相关关系(P<0.05);完全花比例与坐果数呈极显著的正相关关系(P<0.01),与枝径、坐果率呈显著正相关关系(P<0.05)。 本研究所选的256份种质资源来自于13个不同种源地,样品范围广泛且消除了生态环境的影响,树龄都在8年以上,具有代表性。变异系数越大说明性状在个体间的差异越大,遗传多样性程度越高[18]。已有研究表明,自然地理隔离造成了西伯利亚杏形态特征差异明显,变异类型较多[19]。在本研究中,西伯利亚杏花数量性状变异程度高,变异幅度大,存在丰富的多样性:9个数量性状变异系数均值为19.77%,分布在10.26%~41.41%,花冠径为13.98~31.80 mm,大于《中国果树志:杏卷》中描述的15~27 mm;所有性状的极值相差均在2倍以上,展现了较大的变异幅度。雌蕊、雄蕊是与受精作用直接相关的繁殖器官,其变异程度大于其他花器官性状,这增加了子代变异的可能性。西伯利亚杏花主要性状描述特征如下:花瓣颜色为白色或浅粉红色;花瓣圆形或卵圆形;花萼颜色为红褐色或紫红色,极少数为绿色;花瓣状态平;花单瓣,鲜有重瓣。 雌蕊败育现象在西伯利亚杏中普遍存在,败育花不能正常授粉、受精、结果,是导致其自然结实率低的重要原因。杏获得丰产的基本要求是完全花比例达到10%[5],而西伯利亚杏的雌蕊败育率极低,具有巨大的丰产潜力。本研究中,西伯利亚杏的完全花比例平均值为80.11%,高于普通杏品种仰韶黄杏(47.91%),低于仁用杏品种白玉扁(89.27%)[10],4种雌蕊发育类型中,类型Ⅱ所占比例最高(57.96%)。86.4%的西伯利亚杏完全花比例高于50%,其中41.8%的西伯利亚杏完全花比例达到100%,比刘梦培等[7]对西伯利亚杏抗寒植株的调查结果高。完全花比例的高低直接影响坐果率的大小。本研究中,随着完全花比例的增高,坐果率总体呈先上升后下降趋势。 花粉量和花粉活力的高低直接影响到受精和坐果,开展花粉量和花粉活力的研究对引种栽培和杂交育种工作有重要意义。花粉量的多少与品种特性、树体营养及花芽发育以及气候条件、栽培管理措施等有关[20-23]。花粉直接反映雄配子的育性发育状况,花粉的育性是衡量植株育性最根本、最直接的指标,可育率大于50%时为正常可育[24]。本研究中,西伯利亚杏的花粉量为9 533.00~55 467.00粒/朵,服从正态分布;78.57%花的花粉量分布在12 290.00~40 101.00粒/朵,与刘梦培等[7]的调查结果基本一致;西伯利亚杏的花粉活力为15.89%~85.12%,平均值为47.15%,服从正态分布,42.86%的样本花粉活力大于50%,远大于晚花西伯利亚杏(13.73%)。 本研究中,西伯利亚杏花7个表型性状中,除雄蕊个数外,花长、花冠径、花瓣长、花瓣宽、雄蕊长、雌蕊长6个表型性状间(花长和雌蕊长之间除外)均存在极显著的正相关关系(P<0.01),故可以用6个表型性状中的一个描述、推测另一个性状的变异情况。雌蕊败育率较高花型主要受遗传因素控制(遗传力高达0.99),环境影响较小[25]。本研究中,完全花比例与花冠径、花瓣长、花瓣宽、雌蕊长之间呈极显著正相关关系(P<0.01),与花长之间呈显著正相关关系(P<0.05);花粉量与花瓣长、花瓣宽、雌蕊长、完全花比例之间呈正相关关系,但均未达显著水平;花粉活力与雄蕊个数之间呈极显著负相关关系(P<0.01),这有可能是由于植物本身在花粉活力和雄蕊数量之间的平衡调节所致。雌蕊败育花的多少与品种、成熟期、花芽在结果枝上的着生部位、树木生长的立地条件和营养状况有关[9-10]。本研究中,完全花数与总花数、坐果数之间呈极显著的正相关,这与沈红香等[26]的研究结果类似。总花数越多,完全花比例越高,坐果数越多,而总花数和树体营养状况和大小年现象密切相关。完全花比例与坐果数呈极显著的正相关关系(P<0.01),与枝径、坐果率呈显著正相关关系(P<0.05)。前人研究发现,杏树树冠下部雌蕊败育率大于上部,内膛大于外围,北向的雌蕊败育率最高,西向次之,南向、东向较低,主要是因为树冠中部一般因受光照不足、光和作用弱等因素的影响,花芽分化不充分,完全花比例较低[26]。而本研究中,完全花比例在方位上表现为中>南>东>西>北,在层次上表现为中>上>下,中部的完全花比例最高,这与前人研究[26]不同。有研究认为,雌蕊败育与花芽分化发育后期外界低温有关, 尤其是胚珠原基出现以后的低温[27],如龙王帽和大红袍等仁用杏品种于花前15~20 d,花芽和柱头顶部明显膨大,但随后几天出现了-1.4 ℃的低温,造成部分花柱停止生长、变黑或萎缩干枯[28]。在本研究调查时,即开花前4月5日和4月12日出现了2次明显低温期,温度可能对西伯利亚杏的花芽有一定影响,故使得中部以及内堂的完全花比例反而较高。 [参考文献] [1] 俞德浚.中国植物志:第三十八卷 [M].北京:科学出版社,1986:24-31. Yu D X.Flora of China:vol 38 [M].Beijing:Science Press,1986:24-31. [2] 余海滨.不同地理种源山杏在辽西地区引种适应性研究 [D].沈阳:沈阳农业大学,2014. Yu H B.Research on the adaptability of different geographic provenances wild apricot introduced in western Liaoning [D].Shenyang:Shenyang Agricultural University,2014. [3] 包文泉,乌云塔娜,尹明宇.内蒙古野生山杏优良单株果核性状的遗传变异分析 [J].经济林研究,2014,32(4):13-17,72. Bao W Q,Wuyun T N,Yin M Y.Genetic variation of fine individual plants’ seed characteristics of wild apricot in Inner Mongolia [J].Nonwood Forest Research,2014,32(4):13-17,72. [4] 景士西.关于编制我国果树种质资源评价系统若干问题的商榷 [J].园艺学报,1993,20(4):353-357. Jing S X.On some problem concerned with compiling the descriptors of fruit germplasm in China [J].Acta Horticulturae Sinica,1993,20(4):353-357. [5] 张加延,张 钊.中国果树志:杏卷 [M].北京:中国林业出版社,2003:93-559. Zhang J Y,Zhang Z.Chinese fruit tree:apricot [M].Beijing:China Forestry Publishing House,2003:93-559. [6] 刘 宁,刘威生.杏种质资源描述规范和数据标准 [M].北京:中国农业出版社,2006:9-26. Liu N,Liu W S.Discriptors and data standard for apricot (ArmeniacaMill.) [M].Beijing:China Agriculture Press,2006:9-26. [7] 刘梦培,杜红岩,傅建敏,等.内蒙古居群抗寒西伯利亚杏繁殖生态学研究 [J].西北植物学报,2014,34(6):1143-1151. Liu M P,Du H Y,Fu J M,et al.Reproductive ecology ofArmeniacasibiricawith cold resistant in in Inner Mongolia [J].Acta Bot Boreal-Occident Sin,2014,34(6):1143-1151. [8] 夏春森,陈加美,周 萍,等.杏花雌蕊发育与坐果关系的研究 [J].河北果树,1998(4):10-11. Xia C S,Chen J M,Zhou P,et al.The research of relation between blossom pistil development and fruit-bearing in apricot [J].Hebei Fruits,1998(4):10-11. [9] 魏振东,梁 臣,赵合法,等.仰韶黄杏雌蕊败育及控制技术研究 [J].果树科学,1993,10(1):36-38. Wei Z D,Liang C,Zhao H F,et al.The research of pistil abortive and control technology in “Yangshao” yellow apricot [J].Journal Fruit Science,1993,10(1):36-38. [10] 普崇连,王玉柱.杏树坐果率低的原因调查与分析 [J].中国果树,1986(3):23-25. Pu C L,Wang Y Z.The investigation of low fruit-set rate in apricot [J].China Fruit,1986(3):23-25. [11] 刘梦培,乌云塔娜,朱高浦,等.蔷薇科S-RNase基因型鉴定引物在仁用杏基因组中的通用性分析 [J].林业科学研究,2014,27(6):860-865. Liu M P,Wuyun T N,Zhu G P,et al.Transferability of S-RNase genotype identification primers from rosaceae to kernel-apricot [J].Forest Research,2014,27(6):860-865. [12] Yamane H,Ushijimak,Sassah,et al.The use of the S haplotype-specific F-box protein gene,SFB,as a molecular marker for S-haplotypes and self-compatibility in Japanese apricot (Prunusmume) [J].Theor Appl Genet,2003,107:1357-1361. [13] 刘明国,李 民,吴月亮,等.山杏花粉形态特征与花粉萌发的关系 [J].沈阳农业大学学报,2015,46(2):166-172. Liu M G,Li M,Wu Y L,et al.Study on pollen morphological characteristics and its relationship with pollen germination inArmeniacasibirica[J].Journal of Shenyang Agricultural University,2015,46(2):166-172. [14] 刘梦培,杜红岩,傅大立,等.抗寒西伯利亚杏花粉特性及其相关分析 [J].西北林学院学报,2015,30(2):111-115. Liu M P,Du H Y,Fu D L,et al.Pollen characteristics and correlation ofArmeniacasibiricawith cold-resistance [J].Journal of Northwest Forestry University,2015,30(2):111-115. [15] 何春燕,谭晓风,袁德义,等.7种油茶花粉数量及花粉萌发率的测定 [J].中南林业科技大学学报,2009,29(1):74-78. He C Y,Tan X F,Yuan D Y,et al.Determination of the pollen number and pollen germination rate of sevenCamelliaspecies[J].Journal of Central South University of Forestry & Technology,2009,29(1):74-78. [16] 廖 婷,袁德义,袁 军,等.4个油茶优良无性系花器特征及花粉萌发率 [J].中南林业科技大学学报,2010,30(9):51-54. Liao T,Yuan D Y,Yuan J,et al.Flower characteristics and pollen germ ination rate of fourCamelliaoleiferaclones [J].Journal of Central South University of Forestry & Technology,2010,30(9):51-54. [17] 马小河,赵旗峰,董志刚,等.鲜食葡萄品种资源果实数量性状变异及概率分级 [J].植物遗传资源学报,2013,14(6):1185-1189. Ma X H,Zhao Q F,Dong Z G,et al.Variation and probability grading of main quantitative traits of table grape resources [J].Journal of Plant Genetic Resources,2013,14(6):1185-1189. [18] 聂石辉,彭 琳,王 仙,等.鹰嘴豆种质资源农艺性状遗传多样性分析 [J].植物遗传资源学报,2015,16(1):64-70. Nie S H,Peng L,Wang X,et al.Genetic diversity of agronomic traits in chickpea (CicerarietinumL.) germplasm resources [J].Journal of Plant Genetic Resources,2015,16(1):64-70. [19] 李丽锋,刘明国,李立新.辽西地区山杏立地条件的主成分-数量化分析 [J].生物数学学报,2008,23(4):687-694. Li L F,Liu M G,Li L X,et al.The principal component analysis of site conditions ofArmeniacasibiricain Western Liaoning [J].Journal of Biomathematics,2008,23(4):687-694. [20] 许 方.梨树生物学 [M].北京:科技出版社,1992:35. Xu F.Biology of pear [M].Beijing:Science & Technology Press,1992:35. [21] 龙兴桂.现代果树栽培:落叶果树卷 [M].北京:中国林业出版社,2000:418-457. Long X G.Modern Chinese fruit cultivation:volume of deciduous fruit crops [M].Beijing:Chinese Forestry Publishing House,2000:418-457. [22] Tadashi S,Hideyuki T,Iwao N,et al.Effects of high temperature on the development of pollen mothercells and microspores in barleyHordeumvulgweL. [J].J Plant Res,2000,113:395-402. [23] Sedgley M,Griffin A R.Sexual reproduction of tree crops [M].London:Academic Press,1989:529-540. [24] 朱国英.水稻雄性不育生物学 [M].武汉:武汉大学出版社,2000:143-146. Zhu G Y.Biology of sterile rice [M].Wuhan:Wuhan University Press,2000:143-146. [25] 撒文清,魏安智,杨途熙,等.黄土沟壑区旱地仁用杏品种不同花型的数量分布及遗传分析 [J].西北林学院学报,2004,19(1):50-51. Sa W Q,Wei A Z,Yang T X,et al.Heredity analysis and flower pattern quatity distribution of the kernel apricot of drought land in Loess Gully [J].Journal of Northwest Forestry University,2004,19(1):50-51. [26] 沈红香,孔 云,姚允聪,等.杏花雌蕊败育的研究进展 [J].中国果树,2007(3):57-59. Shen H X,Kong Y,Yao Y C,et al.The research of pistil abortion in apricot [J].China Fruit,2007(3):57-59. [27] 李隐生,黄 郊,霍天喜,等.杏花芽分化观察 [J].园艺学报,1986,13(1):68-70. Li Y S,Huang J,Huo T X,et al.Flower bud differentiation observation of apricot [J].Acta Horticulturae Sinica,1986,13(1):68-70. [28] 王保明,丁改秀,童德中,等.仁用杏雌性器官败育研究 [J].山西农业科学,2000,28(1):57-61. Wang B M,Ding G X,Tong D Z,et al.A study on the pistils abortion of the kernel apricot [J].Journal of Shanxi Agricultural Sciences,2000,28(1):57-61.2 结果与分析
2.1 西伯利亚杏花性状的多样性
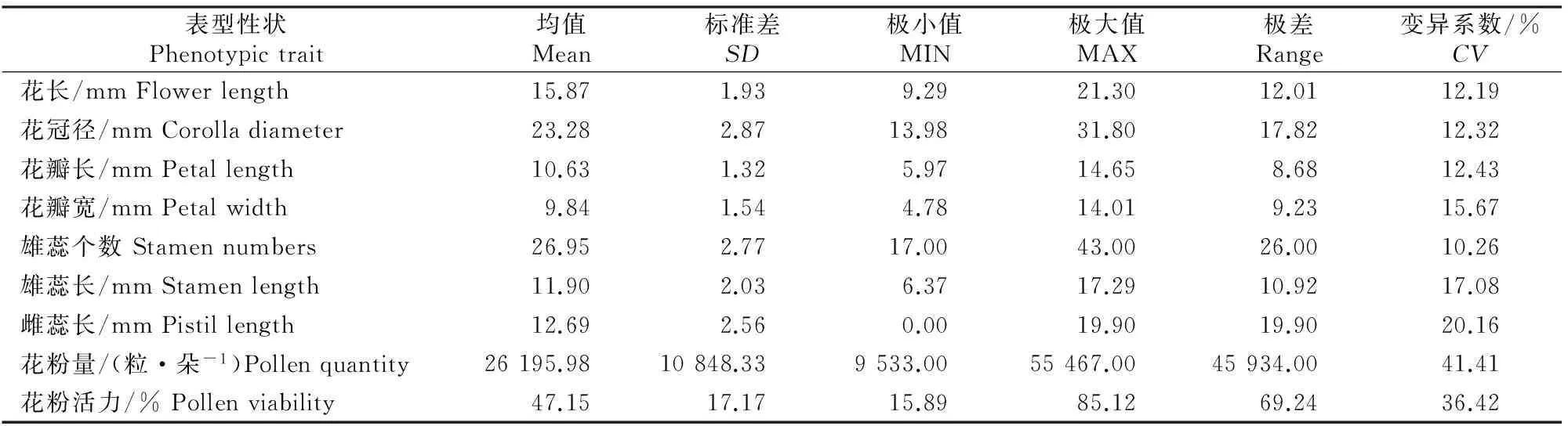
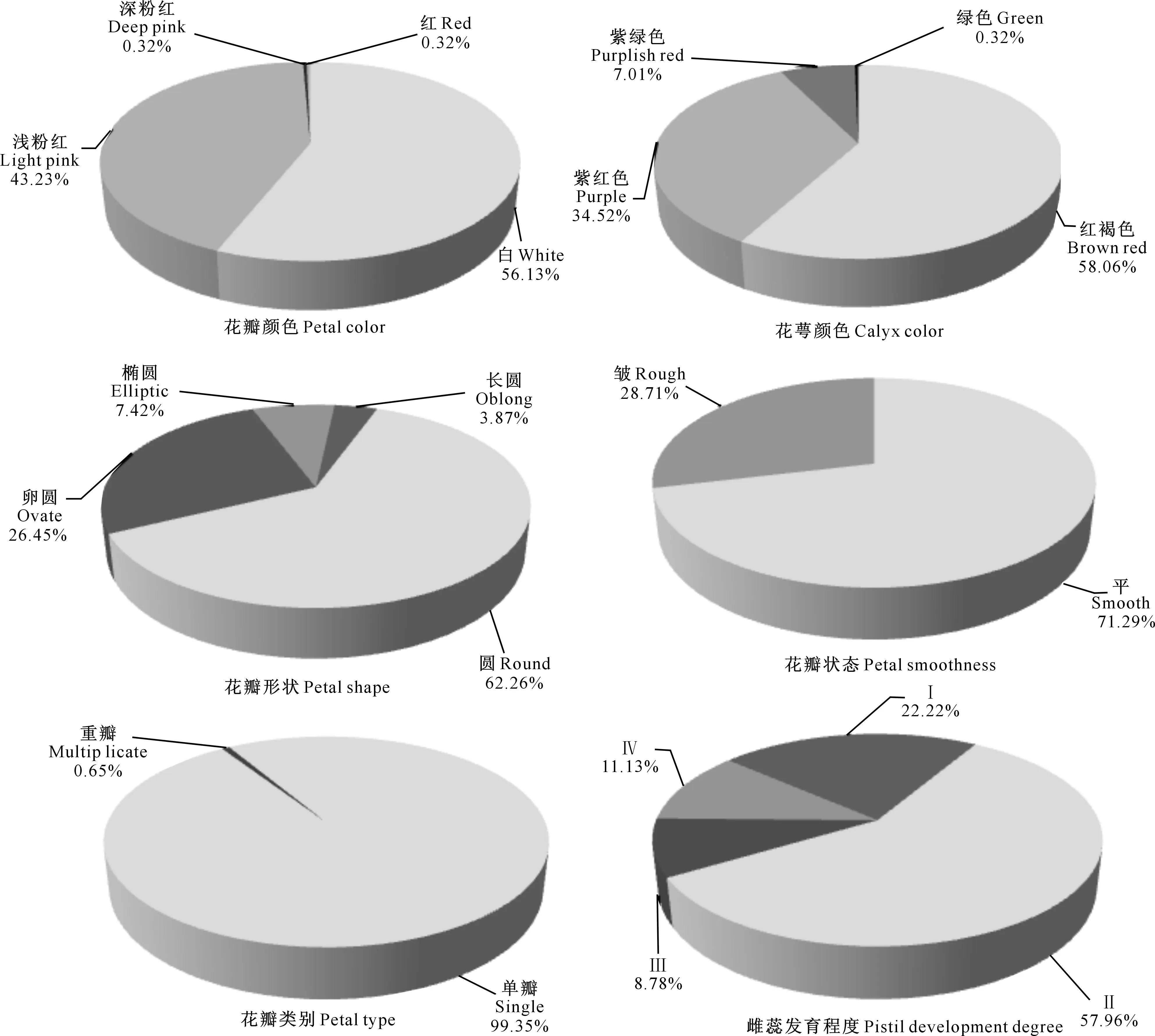
2.2 西伯利亚杏花数量性状的正态性检验及概率分级
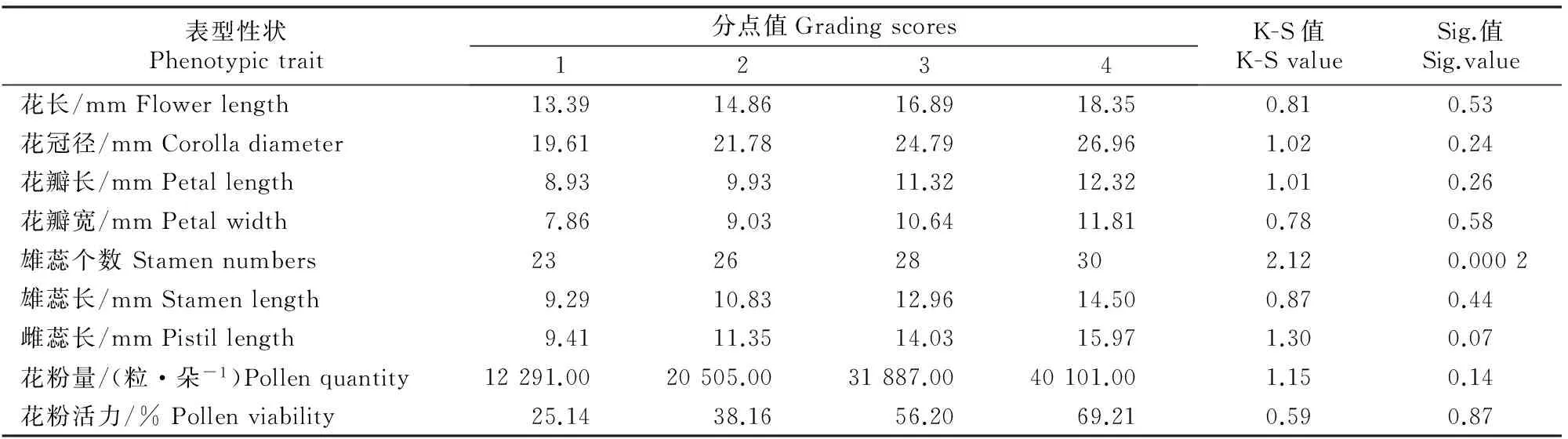
2.3 西伯利亚杏花数量性状概率分级的频率分布

2.4 西伯利亚杏的雌蕊发育程度以及在方位和层次上的分布
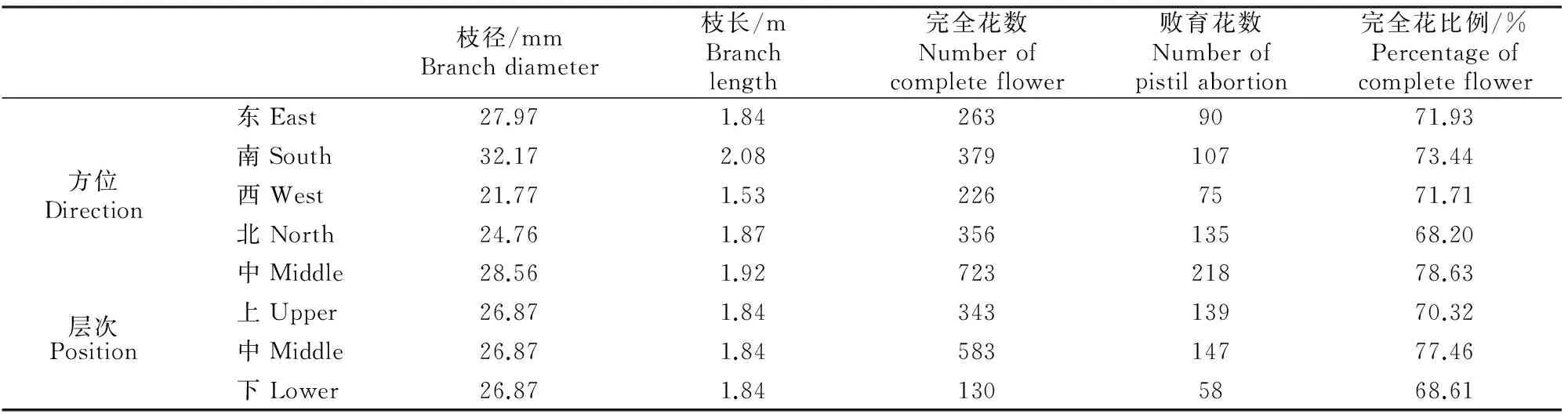
2.5 西伯利亚杏花表型性状与花粉特性以及雌蕊发育程度与坐果率的相关性
3 讨 论
3.1 西伯利亚杏花表型特性
3.2 西伯利亚杏雌蕊发育程度、花粉量和花粉活力
3.3 西伯利亚杏花表型性状、花粉特性以及雌蕊发育程度与坐果率的相关性
