ldentification of the strain-specifically truncated nonstructural protein 10 of porcine reproductive and respiratory syndrome virus in infected cells
2018-05-08ZHANGZhibangXULeiWENXuexiaDONGJianguoZHOULeiGEXinnaYANGHanchunGUOXin
ZHANG Zhi-bang, XU Lei, WEN Xue-xia, DONG Jian-guo, ZHOU Lei, GE Xin-na, YANG Han-chun,GUO Xin
Key Laboratory of Animal Epidemiology and Zoonosis, Ministry of Agriculture/College of Veterinary Medicine, China Agricultural University/State Key Laboratory of Agrobiotechnology, China Agricultural University, Beijing 100193, P.R.China
1. lntroduction
Porcine reproductive and respiratory syndrome (PRRS) is one of the most important viral diseases that have caused huge economic losses to the pig industry worldwide (Zhou and Yang 2010; Holtkampet al. 2013). PRRS is mainly characterized with respiratory problems in piglets and severe reproductive illness in sows and gilts (Pejsaket al. 1997).The etiologic agent, porcine reproductive and respiratory syndrome virus (PRRSV), belongs to theArterivirusgenus of theArteriviridaefamily, together with equine arteritis virus (EAV), simian hemorrhagic fever virus (SHFV) and lactate dehydrogenase-elevating virus (LDV) (Snijderet al. 2013). Given the genetic and antigenic diversity,PRRSV can be classi fied into two genotypes: European(genotype 1) and North American (genotype 2), which share 55–70% nucleotide identity (Menget al. 1994; Nelsenet al. 1999). In 2006, atypical PRRS outbreak caused by the highly pathogenic PRRSV (HP-PRRSV) emerged in China, which brought fatal pathogenicity to pigs compared to previous PRRS caused by classical low pathogenic PRRSV (LP-PRRSV) strains (Tianet al. 2007; Tonget al.2007; Zhouet al. 2009). More recently, the NADC30-like PRRSV isolates were reported (Zhouet al. 2015) and experimental animal assays revealed that importation and recombination are responsible for the latest emergence of highly pathogenic PRRSV in China (Zhaoet al. 2015).Both the North American and European PRRSV isolates are circulating in China and the HP-PRRSVs have become the dominant strains (Chenet al. 2011; Liet al. 2011; Xieet al. 2014).
PRRSV is an enveloped, positive-stranded RNA virus with a genome of about 15 kb in length, containing two large open reading frames (ORF1a and ORF1b) and several downstream ORFs (ORFs 2 to 7) (Meulenberg 2000). ORFs 2 to 7 encode the structural proteins GP2, E,GP3, GP4, GP5, M, N and the newly discovered ORF5a protein, respectively (Snijder and Meulenberg 1998; Firthet al. 2011; Johnsonet al. 2011). ORF1a and ORF1b encode the viral replicase polyproteins pp1a and pp1ab,which are subsequently processed to 16 non-structural proteins (nsps) by viral proteases (Fanget al. 2012; Rascón-Casteloet al. 2015). The expression of ORF1b involved a –1 ribosomal frameshift in the overlapping ORF1a/1b region (den Boonet al. 1991). ORF1b is composed of nsp9 (RNA-dependent RNA polymerase), nsp10 (helicase),nsp11 (endoribonuclease) and nsp12 (unknown function)(Snijderet al. 2013; Shiet al. 2016). In particular, nsp9 and nsp10 are believed to be the center components during viral replication and genomic transcription (van Dintenet al.2000). Our latest studies indicated that nsp9 and nsp10 regions of HP-PRRSV not only affected the replication efficiency of HP-PRRSVin vitrobut also contributed to its fatal virulence for piglets (Liet al. 2014).
In this study, we found that HP-PRRSV strain JXwn06 infection could produce a truncated form of nsp10 (termed nsp10a). In contrast to the full-length nsp10, nsp10a is probably short of about 70 amino acids at the N terminus.Next, we identified the residues in nsp10 that affect the generation of nsp10a and analyzed the influence of nsp10a on the growth of PRRSVin vitro.
2. Materials and methods
2.1. Cells, viruses and antibody
Porcine alveolar macrophages (PAMs) were obtained from the lung lavage fluid of specific-pathogen-free (SPF) pigs as previously described (Zhanget al. 2009) and maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum and penicillin/streptomycin. The MARC-145 cell line and the human embryonic kidney HEK-293FT cell line were cultured in Dulbecco’s modified Eagle’s medium (DMEM)with 10% FBS at 37°C. Two PRRSV strains, JXwn06 and HB-1/3.9 were isolated and kept in our laboratory (Zhouet al. 2009). Five vaccine strains, HuN4-F112 (GenBank no.unavailable), TJM-F92 (GenBank no. unavailable), JXA1-R(GenBank no. unavailable), RespPRRS MLV (GenBank no. AF066183) and CH-1R (GenBank no. EU807840),were purchased from market. Mouse anti-PRRSV nsp10 monoclonal antibody (mAb) was prepared in our laboratory(Zhanget al. 2017).
2.2. Plasmid construction and DNA transfection
Plasmids were constructed by standard recombinant DNA procedures. A series of eukaryotic plasmids expressing different fragments of nsp10 were constructed. The DNA fragments of nsp10 gene were amplified by PCR using the plasmid pWSK-CMV-JXwn06 as the template with different primer sets (Table 1) and then subcloned into the eukaryotic vector pEGFP-N1viaBamHI/KpnI sites. A stop codon was placed just downstream of inserted DNA fragments to prevent fusing with GFP tag. In this way, the expressed nsp10 fragments will reflect their real sizes as they don’t include any irrelevant amino acid.
Transfection was conducted using Lipofectamine®LTX with Plus™ Reagent (Invitrogen, USA) according to themanufacturer’s instruction. Briefly, HEK-293FT cells grown to 80% confluence in 6-well plates were transfected with transfection mix, which contains 2.5 μg of plasmid, 2.5 μL of Plus reagent and 5 μL of Lipofectamine®LTX in Opti-MEM.HEK-293FT cells were cultured for 24 h post-transfection for protein expression.

Table 1 The primers for construction of nsp10 deletion mutants
2.3. Recovery of recombinant viruses from infectious clones
The infectious clone plasmids of JXwn06 and HB-1/3.9,pWSK-CMV-JXwn06 and pWSK-CMV-HB-1/3.9 were constructed and kept in our laboratory. Site-directed mutants of nsp10 gene were generated by PCR-directed mutagenesis and cloned into plasmid pWSK-CMV-JXwn06 or pWSKCMV-HB-1/3.9 to generate infectious clones containing expected mutants. All the constructed infectious clones were verified by DNA sequencing.
MARC-145 cells grown to 70–80% confluence in sixwell plates were transfected with 2.5 μg of infectious cDNA clones using Lipofectamine®LTX with with Plus™Reagent. The culture supernatants of MARC-145 cells were harvested at 5 d post-transfection when obvious cytopathic effect (CPE) appeared, and the rescued viruses were designated ‘passage-1’. The designed mutant bases in the RNAs of third-passage viruses were con firmed by DNA sequencing.
2.4. Silver staining and mass spectrometric (MS)analysis
The cell lysates of PRRSV-infected cells were resolved by electrophoresis on 10% SDS-PAGE gels and stained using a Silver Stain Kit for Mass Spectrometry (Thermo Scientific, Rockford, IL) according to the manufacturer’s protocol. Prestained protein ladder was used for estimating the approximate sizes of separated proteins. The protein band corresponding to nsp10a in the gel was excised and subjected to MS analysis as described previously (Duet al.2016).
2.5. Western blotting analysis
PRRSV-infected cells were lysed with RIPA buffer and then the cell lysates were cleared by centrifuging at 12 000 r min–1for 20 min. The protein concentrations in the supernatants were quantified by using a bicinchoninic acid (BCA) protein assay kit (Pierce, USA). Every protein samples with 30 μg were first fractionated by SDS-PAGE, and then transferred onto a polyvinylidene fluoride (PVDF) membrane. After blocking with 5% skim milk powder in PBST (0.05% Tween 20), the membrane was probed with anti-nsp10 mAb. The membrane was washed with 0.05% PBST and incubated with horseradish peroxidase (HRP)-conjugated goat antimouse IgG (ZSGB-BIO, China). At last, the membrane was washed and developed with the ECL Western Blotting System (Pierce, USA).
2.6. lmmuno fluorescence microscopy
MARC-145 cells grown on coverslips in six-well plates were infected with PRRSV strain JXwn06 at multiplicity of infection(MOI) of 0.1. At 12, 16, 20, 24 and 32 h post-infection (hpi),the cells were fixed with 3.7% paraformaldehyde for 10 min at room temperature (RT), and permeabilized with PBS containing 0.1% Triton X-100 and 2% BSA for 10 min, then blocked with 2% BSA in PBS for 30 min. MARC-145 cells were stained with nsp10-specific mAb in a humidity chamber for 1 h at RT. After three times of 5 min washing with PBS, the cells were then incubated with Alexa Fluor 488-conjugated goat anti-mouse IgG F(ab´)2fragment (Molecular Probes)antibody for 1 h at RT. Nuclear DNA was stained with 4´,6-diamidino-2-phenylindole (DAPI) (Molecular Probes) for 5 min at RT. The images were collected with an Olympus confocal microscope (Fluoview1000, Japan).
2.7. Virus infection and titration
MARC-145 cells grown to full con fluence in 24-well plates were infected with RvJX-E69G and RvHB-G69E or their parental viruses (RvJXwn06 and RvHB-1/3.9) at MOI of 0.01 in DMEM with 5% FBS. The titers of viruses in the supernatants at different time points were determined using a microtitration infectivity assay and expressed as 50% tissue culture infective dose per mL (TCID50mL–1) as previously described (Zhouet al. 2009). Each time point was independently repeated three times.
2.8. Statistical analysis
The data were expressed as means±standard deviations(SD). All statistical analyses were performed using GraphPad Prism version 5.0 software. Statistical significance among different groups was determined by two-way ANOVA analysis of variance.
3. Results
3.1. PRRSVs produce a truncated nsp10 protein in a strain-specific manner
At the beginning of this study, two PRRSV strains,HP-PRRSV JXwn06 and LP-PRRSV HB-1/3.9, were chosen to infect MARC-145 cells, respectively. At 48 hpi, the cell lysates were prepared and subjected to Western blotting probed by anti-nsp10 mAb. Interestingly, two specific bands, with estimated molecular weight of 50 and 42 kD,were detected in samples from JXwn06 infected MARC-145 cells (Fig. 1-A). The upper band is corresponding to the expected full-length nsp10, while the lower one is probably a truncated form of nsp10. For convenience, we named the full-length one nsp10 and the truncated form nsp10a hereafter. However, unlike JXwn06, HB-1/3.9 infection only produced the full-length nsp10 in MARC-145 cells (Fig. 1-A). Next, we tested the expression of nsp10 products in PRRSV-infected PAMs, the target cells for PRRSV. The result in PAMs is similar to that in MARC-145 cells, indicating the production of nsp10a is not dependent on specific cell types (Fig. 1-A).
To determine if the production of nsp10a is a unique phenomenon for the strain JXwn06 or it’s universal for other PRRSV strains, five PRRSV vaccine strains that were easily available from market were chosen to infect MARC-145 cells. The infection of TJM-F92, JXA1-R, RespPRRS MLV or CH-1R produced both nsp10 and nsp10a, while HuN4-F112 infection only generated nsp10 (Fig. 1-B). Taken together,the generation of nsp10a is strain-specific.

Fig. 1 The production of nsp10a is in a strain-specific manner.A, Identification of nsp10 products in JXwn06- or HB-1/3.9-infected cells. MARC-145 cells or PAMs were infected with JXwn06 and HB-1/3.9. At 48 h post-infection (hpi), the cells were harvested and the cell lysates were separated by SDSPAGE followed by Western blotting with the nsp10-specific mAb. B, Western blotting of nsp10 products in MARC-145 cells infected with different vaccine strains.
3.2. The expression kinetics of nsp10 products during PRRSV infection
In order to investigate the kinetics of nsp10 and nsp10a during PRRSV infection, MARC-145 cells were infected with JXwn06 and the infected cells were subjected to confocal microscopy and Western blotting analysis at indicated time points post-infection using anti-nsp10 mAb. The fluorescence signal appeared at 12 hpi and its intensity increased as the time goes on (Fig. 2-A). By 32 hpi, bright fluorescent foci appeared in ~80% of the cells (Fig. 2-A).The intracellular localization of PRRSV nsp10 is similar to that of PRRSV nsp2, nsp4, nsp7α, nsp7β and nsp8, in the perinuclear region of infected cells (Liet al. 2012). In Western blotting analysis, the full-length nsp10 was first detected at 24 hpi, reached peak level at 60 hpi and started to wane at 72 hpi in JXwn06-infected cells (Fig. 2-B). While nsp10a appeared later than 24 hpi, however, increased until 72 hpi and became the predominant form of nsp10 products at the later time points (Fig. 2-B).
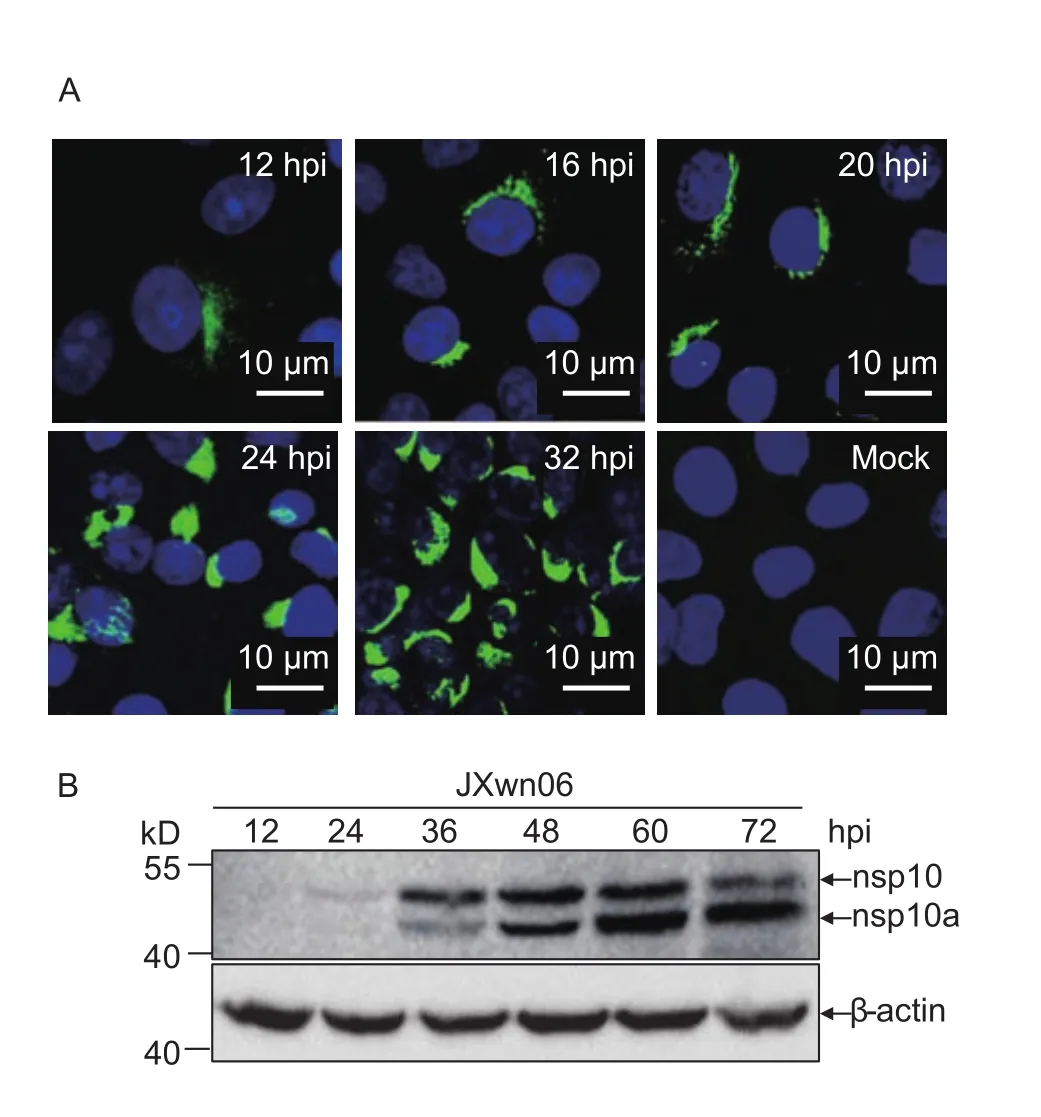
Fig. 2 The expression kinetics of nsp10 products in porcine reproductive and respiratory syndrome virus (PRRSV)-infected MARC-145 cells. A, Identification of nsp10 products by immuno fluorescent assay (IFA). MARC-145 cells were infected with JXwn06 at multiplicity of infection (MOI) 0.1. The infected cells were subjected to IFA using anti-nsp10 mAb at indicated time points post-infection. B, the expression trend of nsp10 products in JXwn06-infected cells. MARC-145 cells were infected with JXwn06 at MOI of 0.01. The cell lysates were separated by SDS-PAGE and subjected to Western blotting with anti-nsp10 mAb. hpi, hours post-infection. Bar=10 μm.
3.3. Nsp10a may be an N-terminally truncated form of full-length nsp10
Since nsp10a could be recognized by anti-nsp10 mAb,nsp10a is likely a truncated form of nsp10. To con firm the identity of nsp10a, JXwn06-infected cell lysates were harvested at 48 hpi and separated by SDS-PAGE, and then the gels were stained using a Silver Stain Kit (Thermo,Rockford, IL). The nsp10a bands in gels were identified by comparing to that of the protein standards as well as the full-length nsp10. The nsp10a-containing bands were excised from the gels and subjected to mass spectrometry(MS) analysis. The identified peptide sequences were searched on the NCBI website using the protein BLAST(blastp) suite. The peptides that matched to PRRSV nsp10 were aligned with the amino acid sequences of nsp10 and showed in Fig. 3-A. In Fig. 3-A, the green lines represent the identified peptides whose amino acid sequences are identical to that shown above the green lines, while the number of green lines stand for how many times of the peptides been identified in separate experiments. The results of MS revealed that nsp10a was indeed a truncated form of nsp10. The lengths of the peptides mostly ranged from 9 to 20 amino acids except one with 34 amino acids and all the identified peptides mapped to the sequence of nsp10 between amino acids 78 and 423 (Fig. 3-A).
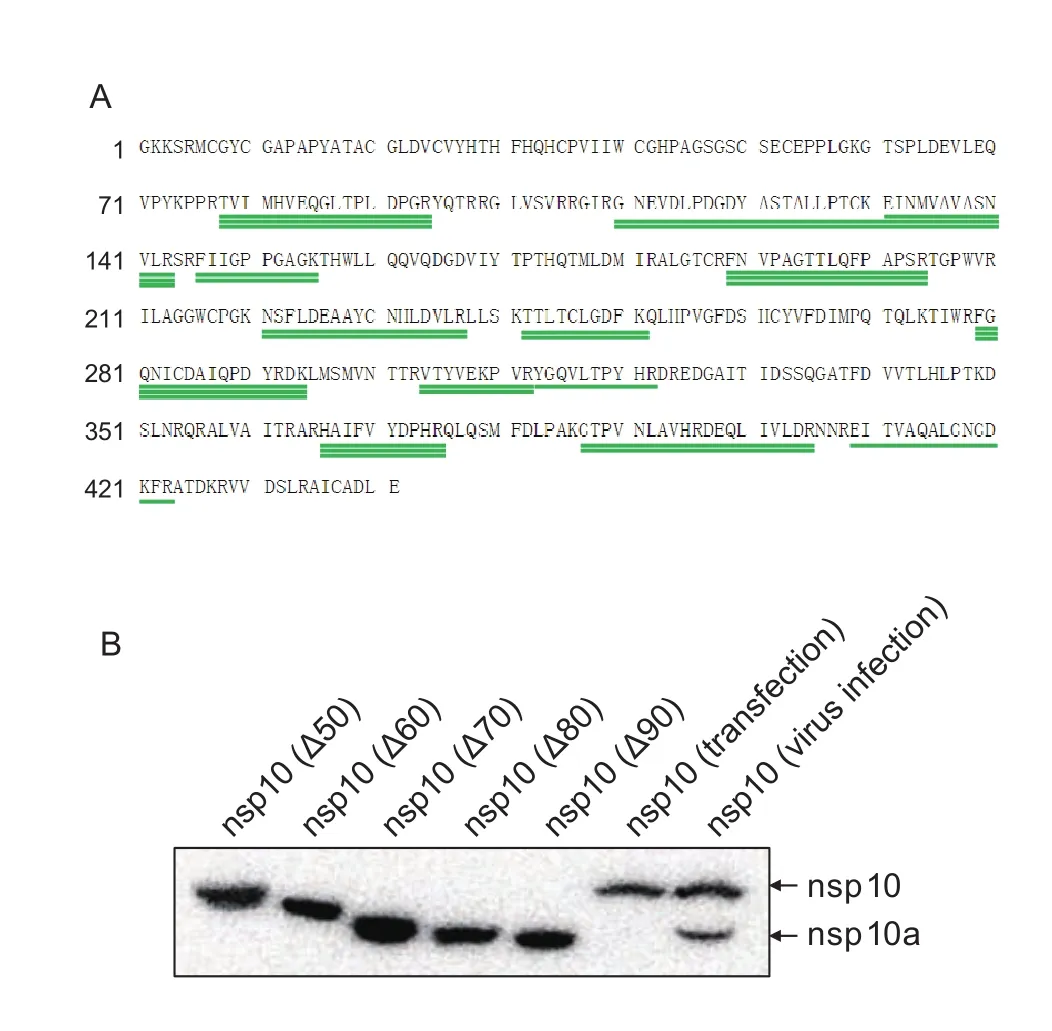
Fig. 3 Identification of nsp10a by mass spectrometric (MS) and deletion mutagenesis. A, identify the amino acid sequence of nsp10a by MS analysis. JXwn06-infected cell lysates were separated by electrophoresis on a 10% polyacrylamide gel, and stained by silver staining. The protein band corresponding to nsp10a was excised from the gel and subjected to MS analysis.The identified peptides were shown by green lines. The amino acids above each green line are the amino acid sequences of the identified peptide and the frequencies of each green line represent the identified times of the peptides from separated tests. B, eukaryotic expression of nsp10 and its deletion mutants. The nsp10 products from plasmid transfected cells were compared to that from JXwn06-infected cells by Western blotting analysis.
To identify the size of nsp10a, a series of nsp10 proteins with designed deletions were expressedin vitroand compared to nsp10a from virus-infected cell lysates.Five recombinant plasmids based on the eukaryotic vector pEGFP-N1 were constructed by deletion mutagenesis,which expressed nsp10 proteins with deletions ranging from 50 to 90 amino acids at the N-terminus. HEK-293FT cells transfected with the constructed plasmids were harvested 24 h post-transfection, and subjected to Western blotting analysis by anti-nsp10 mAb. The results indicated that the eukaryotic expressed full-length nsp10 protein had a gel migration pattern similar to that of natural nsp10 protein from JXwn06-infected cells (Fig. 3-B). While the truncated nsp10 with 70 amino acids deletion migrated as fast as nsp10a in gel. Western blotting results coupled with the MS data revealed that nsp10 and nsp10a may share the same C-terminus and nsp10a is probably short of about 70 amino acids in the N-terminus.
3.4. Amino acid 69-Glu in nsp10 is the key residue that affects the production of nsp10a
As shown above, the production of nsp10a is strain-dependent. However, the determinants affecting the generation of nsp10a are still unknown. To address this question, four chimeric viruses, RvHJn10, RvHJn9n10, RvJHn10, and RvJHn9n10, were used to infect MARC-145 cells. The four chimeric viruses were made previously by swapping nsp10 gene alone or nsp9 and nsp10 genes together between JXwn06 and HB-1/3.9 (Liet al. 2014). The infected cell lysates were prepared at 48 hpi for Western blotting analysis. The Western blotting results showed that when nsp10 gene or both nsp9 and nsp10 genes in JXwn06 were substituted by that of HB-1/3.9, the chimeric viruses RvJHn10 and RvJHn9n10 could not generate nsp10a (Fig. 4-A). In contrast, the chimeric viruses RvHJn10 and RvHJn9n10,which replaced nsp10 gene or nsp9 and nsp10 genes on HB-1/3.9 backbone with that of JXwn06, produced nsp10a(Fig. 4-A). These data suggested that the generation of nsp10a was dictated by nsp10 gene itself, but not other genes of PRRSV.
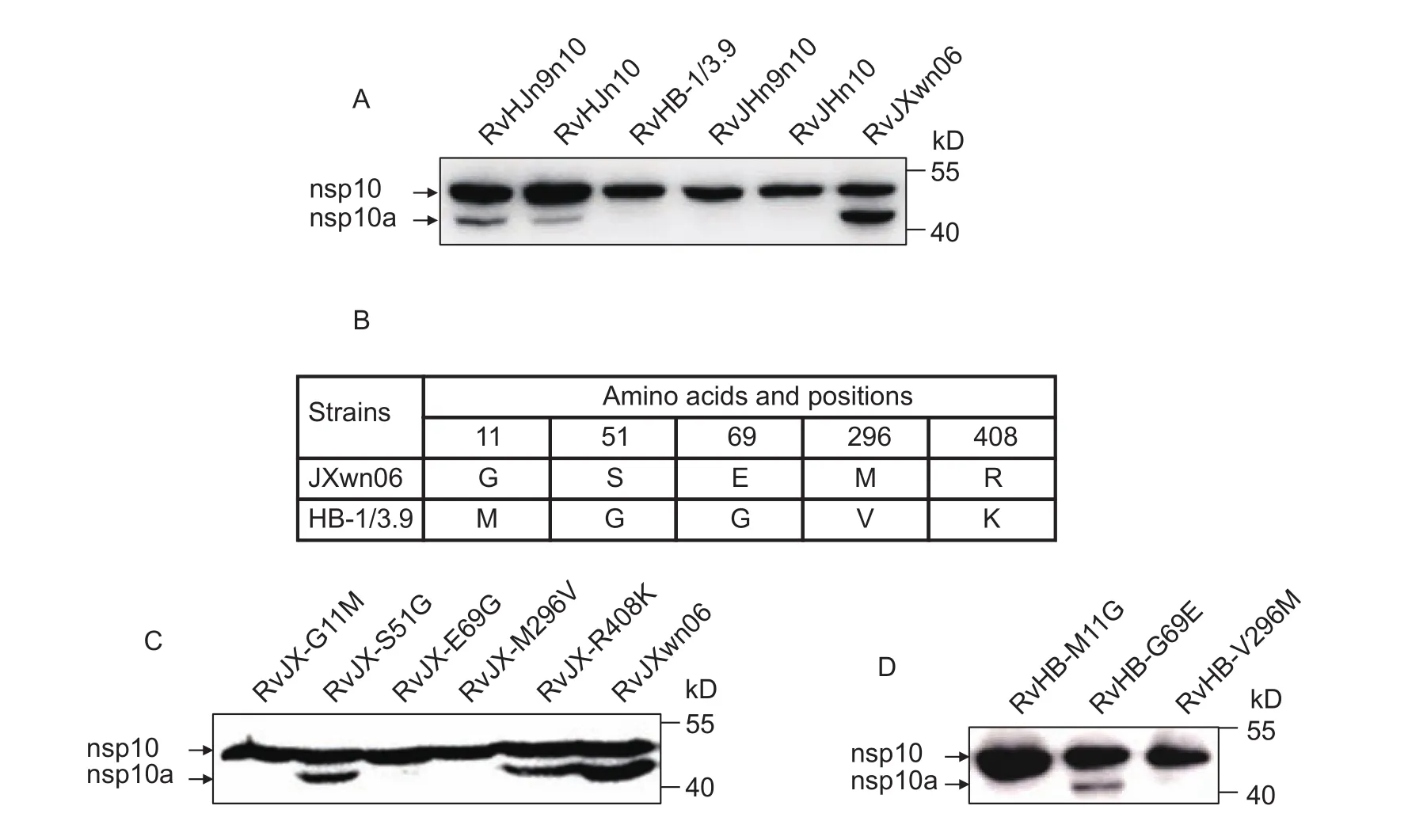
Fig. 4 Identification of the amino acids that affect nsp10a production. A, the production of nsp10a was dictated by nsp10 gene.Four chimeric viruses, RvHJn10, RvHJn9n10, RvJHn10, RvJHn9n10, with swapped nsp10 gene or nsp9 and nsp10 genes together,were used to infect MARC-145 cells. Porcine reproductive and respiratory syndrome virus (PRRSV)-infected cell lysates were subjected to Western blotting with the nsp10-specific mAb. B, there are five different amino acids in nsp10 between JXwn06 and HB-1/3.9. C and D, detection of nsp10 products in single point mutant virus-infected cell lysates by Western blotting with the nsp10-specific mAb.
Sequence alignment of nsp10 between JXwn06 and HB-1/3.9 showed that there were five different amino acid residues, which were located at 11, 51, 69, 296, 408 amino acids in nsp10 (Fig. 4-B). To determine the key determinants that involved in nsp10a production, we constructed five viruses on the backbone of JXwn06 by mutating the five amino acids one by one to the corresponding residues of HB-1/3.9. Western blotting analysis showed that the rescued viruses RvJX-G11M, RvJX-E69G and RvJX-M296V could not generate nsp10a in infected cells. The results suggested that the amino acids at 11, 69 and 296 affect the production of nsp10a (Fig. 4-C). Then, we rescued three viruses on the backbone of HB-1/3.9 by mutating M to G, G to E and V to M at 11, 69 and 296 in nsp10, respectively. Further analysis revealed that only the rescued virus RvHB-G69E partially gained the ability to produce nsp10a (Fig. 4-D). Together,the data indicated that the amino acid 69-Glu in nsp10 is the key amino acid determining the production of nsp10a.
3.5. Nsp10a is not essential for PRRSV replication in cell culture
To analyze the effect of the production of nsp10a on PRRSV replication, the growth kinetics of RvJX-E69G and RvHBG69E in parallel with their parental viruses, RvJXwn06 and RvHB-1/3.9, were evaluated in MARC-145 cells. The results showed that RvJX-E69G and RvHB-G69E exhibited comparable growth kinetics to their parental viruses at indicated time points (Fig. 5). As shown above, RvJX-E69G infection could not produce nsp10a while RvHB-G69E infection could generate nsp10a, which were both different from their parental viruses, respectively. Therefore, the generation of nsp10a had little effect on PRRSV replicationin vitro.
4. Discussion
The truncated form of nsp10 protein (nsp10a) generated in PRRSV-infected cell lysates was reported for the first time in the present study. We have demonstrated that the generation of nsp10a is dictated by nsp10 gene, but we cannot rule out the possibility that viral other genes are involved in this process. In fact, as shown in Fig. 4-A, nsp9 and nsp10 genes together displayed a synergistic effect for nsp10a production. Although the overall quantities of nsp10 products in RvHJn9n10 group are less than that in RvHJn10 group, the amount of nsp10a in RvHJn9n10 group is more than that in RvHJn10 group (Fig. 4-A). Therefore, other genes of PRRSV, such as nsp9, although did not determine the production of nsp10a, but might influence the efficiency of nsp10a production.
Previous study has shown that nsp9 and nsp10 of HPPRRSV contributed to its fatal virulence (Liet al.2014). In the present study, we demonstrated that the production of nsp10a had little in fluence on PRRSV replication. Whether nsp10a correlated with viral virulence is unknown. Based on the results of this study, we can infer that nsp10a was probably not correlated with PRRSV virulence, as both the HP-PRRSV strain JXwn06 and the LP-PRRSV strain CH-1R could produce nsp10a. However, to get the fact, anin vivoexperiment is necessary.

Fig. 5 Comparison of the growth kinetics of RvJX-E69G and RvHB-G69E with their parental viruses. MARC-145 cells were infected RvJX-E69G, RvHB-G69E, RvJXwn06 and RvHB-1/3.9 at multiplicity of infection (MOI) of 0.01, respectively.Virus titers at 12 to 72 h post-infection (hpi) were determined by microtitration infectivity assays. The data were shown as means±standard deviations (error bars) from three independent trials.
The conclusion that the amino acid 69-Glu in nsp10 is the key residue determining the production of nsp10a was drawn based on the strains JXwn06 and HB-1/3.9. However,there is a discrepancy of the amino acids that affect the production of nsp10a between the two strains JXwn06 and HB-1/3.9. The change of amino acids at 11, 69 and 296 resulted in a loss of nsp10a production on the backbone of JXwn06, while only the amino acid at 69 in HB-1/3.9 nsp10 could alter the phenotype. A probable explanation for this is that the generation of nsp10a depend on a suitable spatial structure. Maybe the change of 11th and 296th amino acids in JXwn06 nsp10 resulted in the change of spatial structure of nsp10 which blocks the production of nsp10a. Similarly, it is possible that the Gly to Glu change at site 69 in HB-1/3.9 nsp10 produce the suitable spatial structure, which is ready for the production of nsp10a. The conclusion, the amino acid 69-Glu in nsp10 is the key residue determining the production of nsp10a, can be further veri fied by the result shown in Fig. 1-B. Four out of five vaccine strains have amino acid 69-Glu in nsp10 (Table 2) and these four strains generated nsp10a upon infection (Fig. 1-B). However,HuN4-F112 strain has a Gly at the site of 69 in nsp10,and it could not produce nsp10a in infected cells (Fig. 1-B and Table 2). Another example is the NADC30-like strain CHsx1401, which has a Arg at 69th amino acid in nsp10(Table 2), and CHsx1401 infection cannot produce nsp10a(Zhanget al. 2017). All the tested stains mentioned above belong to North American type and the results support the conclusion that the 69-Glu in nsp10 is the key residue determining the production of nsp10a. Since the anti-nsp10 mAb generated in our laboratory could not recognize the European PRRSV strains, we could not determine whether European PRRSVs can generate nsp10a, such as European strain GZ11-G1 (Zhanget al. 2017).
The mechanism to generate different forms of PRRSV nsp10 remains unclear. Various mechanisms are employed to generate different forms of proteins, such as leaky scanning, alternative splicing, proteolytic cleavage and translation reinitiation (Liuet al. 1991a, b; Carter and Roizman 1996; Firthet al. 2011). For example, a novel structural protein in PRRSV termed ORF5a was generatedvialeaky ribosomal scanning (Firthet al. 2011). ORF5a is encoded by an alternative ORF which is not in-frame with that of GP5, although they are expressed from the same subgenomic mRNA (sgRNA) (Johnsonet al. 2011). Thus,the amino acid sequences of ORF5a and GP5 have no homology. Another example is the newfound accessory protein NS7a encoded by porcine deltacoronavirus (PDCoV)(Fanget al. 2017). NS7a is encoded by a separate sgRNA with a non-canonical transcription regulatory sequence(TRS). The separate sgRNA encoding NS7a is identical to the 3´ end of the sgRNA encoding NS7, which is generated by splicing the TRS and coding region of NS7.The earlier study showed that PRRSV nsp2 existed as several isoforms with apparently different C termini in virusinfected MARC-145 cells. Those different nsp2 isoforms were likely processed under proteolytic cleavage occurred cotranslationally orvia–1 and –2 ribosomal frameshifting(Hanet al. 2010; Fanget al. 2012).
Obviously, nsp10a was not derived from alternative splicing or translation reinitiation as the former does not occur, and the latter would mean an M at the N-terminus.There are two possible explanations for nsp10a production,proteolytic cleavage or degradation of nsp10. According to the amount of nsp10avs. that of the full-length nsp10 at different time points post-infection, it seemed that the increase of nsp10a was negatively correlated with the decrease of nsp10 (Fig. 2-B). We propose that nsp10a may be generated through the proteolytic cleavage of the full-length nsp10. To test if the viral proteases, like nsp4,nsp2, nsp1α and nsp1β, could cleave nsp10, they were co-expressedwith JXwn06 nsp10 alone or in combination in HEK-293FT cells. However, the Western blotting results showed that all these viral proteases could not cleavage nsp10 (data not shown). We also established a stable MARC-145 cell line that expresses a double tagged nsp10,HA-nsp10-Flag. The recombinant protein HA-nsp10-Flagwas not cleaved whether the nsp10-expressing cell line was infected with JXwn06 or not (data not shown).
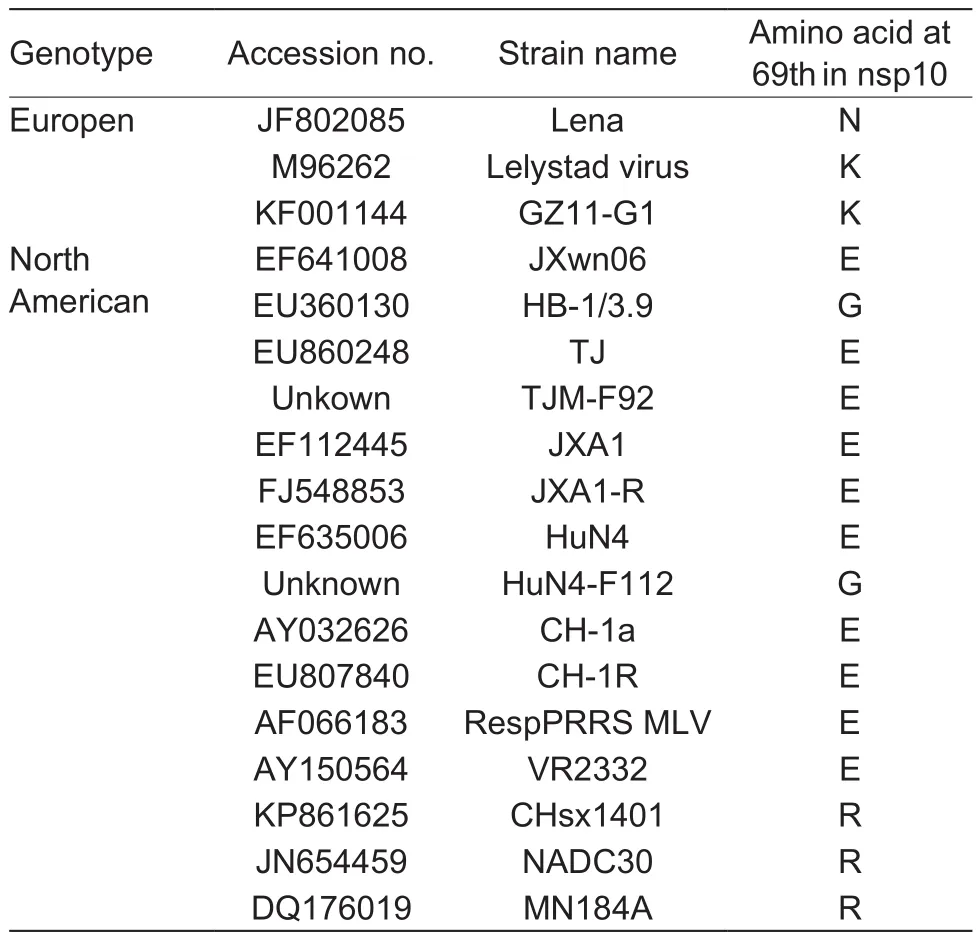
Table 2 The 69th amino acid in nsp10 of different porcine reproductive and respiratory syndrome virus (PRRSV) strains
Equine arteritis virus (EAV) is the prototype of theArterivirusgenus and the studies of EAV provided a general model for PRRSV research. The crystal structure of PRRSV nsp10 is unavailable, whereas the crystal structure of EAV nsp10 has been described recently (Denget al. 2014). EAV nsp10 contained an N-terminus zinc-binding domain (ZBD),a superfamily 1 helicase domain (HEL1) in the C-terminal half and a linker connecting the two domains, and ZBD interacts extensively with the HEL1 domain. The integrity of nsp10 is essential for its helicase properties (Denget al.2014) and this was in line with the previous studies ofArterivirusnsp10, which showed that the ZBD was required for its helicase activity and involved in genome replication(Seybertet al. 2000, 2005). PRRSV nsp10 has ATPase activity and can unwind dsRNA and dsDNA in a 5´ to 3´direction (Bautistaet al. 2002; Zhanget al. 2015).
Protein homology modeling revealed that the structural characteristics of PRRSV nsp10 were similar to that of EAV nsp10. In EAV, the N-terminus 82 amino acids of nsp10 is the ZBD (Bautistaet al. 2002). Accordingly, the N-terminus 64 amino acids in PRRSV nsp10 is the ZBD (http://www.uniprot.org/uniprot/I6XH19). Nsp10a was a truncation of nsp10. The structural deficiency of nsp10a would affect or even block helicase activity. Thus, we inferred that the generation of nsp10a might be unfavorable for PRRSV replication. However, the results showed that the production of nsp10a exerted little influence on the replication of PRRSV in cell culture. It is possible that inactivation of helicase function is instead favorable at later stages of infection,when active RNA replication should be replaced by active packaging and virus egress. Since the tests in the present study were conducted for only 72 hpi,experiments with longer duration are needed to further con firm the speculation bothin vitroandin vivo.
5. Conclusion
In summary, a truncated form of nsp10 termed nsp10a is found in PRRSV-infected cell lysates. Nsp10a may share the same C-terminus with nsp10 but differ in their N-termini.The Glu-69 in nsp10 is the key amino acid determining nsp10a production and the generation of nsp10a imposed little effect on PRRSV replication in cell culture.
Acknowledgements
This work was supported by the National Key Technology R&D Program of China (2015BAD12B01-2), the Major Program of National Natural Science Foundation of China(31490603) and the earmarked fund for Modern Agroindustry Technology Research System of China (CARS-36).
Bautista E M, Faaberg K S, Mickelson D, McGruder E D. 2002.Functional properties of the predicted helicase of porcine reproductive and respiratory syndrome virus.Virology,298, 258–270.
den Boon J A, Snijder E J, Chirnside E D, de Vries A A,Horzinek M C, Spaan W J. 1991. Equine arteritis virus is not a togavirus but belongs to the coronaviruslike superfamily.Journal of Virology,65, 2910–2920.
Carter K L, Roizman B. 1996. The promoter and transcriptional unit of a novel herpes simplex virus 1 alpha gene are contained in, and encode a protein in frame with, the open reading frame of the alpha 22 gene.Journal of Virology,70, 172–178.
Chen N, Cao Z, Yu X, Deng X, Zhao T, Wang L, Liu Q, Li X, Tian K. 2011. Emergence of novel European genotype porcine reproductive and respiratory syndrome virus in mainland China.Journal of General Virology, 92, 880–892.
Deng Z, Lehmann K C, Li X, Feng C, Wang G, Zhang Q, Qi X, Yu L, Zhang X, Feng W, Wu W, Gong P, Tao Y, Posthuma C C, Snijder E J, Gorbalenya A E, Chen Z. 2014. Structural basis for the regulatory function of a complex zinc-binding domain in a replicative arterivirus helicase resembling a nonsense-mediated mRNA decay helicase.Nucleic Acids Research, 42, 3464–3477.
van Dinten L C, van Tol H, Gorbalenya A E, Snijder E J.2000. The predicted metal-binding region of the arterivirus helicase protein is involved in subgenomic mRNA synthesis,genome replication, and virion biogenesis.Journal of Virology, 74, 5213–5223.
Du J, Ge X, Dong H, Zhang N, Zhou L, Guo X, Yang H. 2016.The cellular interactome for glycoprotein 5 of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus.Journal of Integrative Agriculture, 15,1833–1845.
Fang P, Fang L, Hong Y, Liu X, Dong N, Ma P, Bi J, Wang D, Xiao S. 2017. Discovery of a novel accessory protein NS7a encoded by porcine deltacoronavirus.Journal of General Virology, 98, 173–178.
Fang Y, Treffers E E, Li Y, Tas A, Sun Z, van der Meer Y, de Ru A H, van Veelen P A, Atkins J F, Snijder E J, Firth A E.2012. Ef ficient –2 frameshifting by mammalian ribosomes to synthesize an additional arterivirus protein.Proceedings of the National Academy of Sciences of the United States of America, 109, 2920–2928.
Firth A E, Zevenhoven-Dobbe J C, Wills N M, Go Y Y, Balasuriya U B, Atkins J F, Snijder E J, Posthuma C C. 2011. Discovery of a small arterivirus gene that overlaps the GP5 coding sequence and is important for virus production.Journal of General Virology, 92, 1097–1106.
Han J, Rutherford M S, Faaberg K S. 2010. Proteolytic products of the porcine reproductive and respiratory syndrome virus nsp2 replicase protein.Journal of Virology, 84,10102–10112.
Holtkamp D J, Kliebenstein J B, Neumann E J, Zimmerman J J, Rotto H F, Yoder T K, Wang C, Yeske P, Mowrer C L, Haley C. 2013. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers.Journal of Swine Health and Production, 21, 72–84.
Johnson C R, Griggs T F, Gnanandarajah J, Murtaugh M P.2011. Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5 present in all arteriviruses.Journal of General Virology, 92,1107–1116.
Li B, Fang L, Guo X, Gao J, Song T, Bi J, He K, Chen H, Xiao S. 2011. Epidemiology and evolutionary characteristics of the porcine reproductive and respiratory syndrome virus in China between 2006 and 2010.Journal of Clinical Microbiology, 49, 3175–3183.
Li Y, Tas A, Snijder E J, Fang Y. 2012. Identification of porcine reproductive and respiratory syndrome virus ORF1aencoded non-structural proteins in virus-infected cells.Journal of General Virology,93, 829–839.
Li Y, Zhou L, Zhang J, Ge X, Zhou R, Zheng H, Geng G, Guo X, Yang H. 2014. Nsp9 and nsp10 contribute to the fatal virulence of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China.PLoS Pathogens, 10, 1004216–1004216.
Liu F Y, Roizman B. 1991a. The herpes simplex virus 1 gene encoding a protease also contains within its coding domain the gene encoding the more abundant substrate.Journal of virology, 65, 5149–5156.
Liu F Y, Roizman B. 1991b. The promoter, transcriptional unit,and coding sequence of herpes simplex virus 1 family 35 proteins are contained within and in frame with the UL26 open reading frame.Journal of Virology,65, 206–212.
Meng X J, Paul P S, Halbur P G. 1994. Molecular cloning and nucleotide sequencing of the 3´-terminal genomic RNA of the porcine reproductive and respiratory syndrome virus.Journal of General Virology,75, 1795–1801.
Meulenberg J J. 2000. PRRSV, the virus.Veterinary Research,31, 11–21.
Nelsen C J, Murtaugh M P, Faaberg K S. 1999. Porcine reproductive and respiratory syndrome virus comparison:Divergent evolution on two continents.Journal of Virology,73, 270–280.
Pejsak Z, Stadejek T, Markowskadaniel I. 1997. Clinical signs and economic losses caused by porcine reproductive and respiratory syndrome virus in a large breeding farm.Veterinary Microbiology, 55, 317–322.
Rascón-Castelo E, Burgara-Estrella A, Mateu E, Hernández J. 2015. Immunological features of the non-structural proteins of porcine reproductive and respiratory syndrome virus.Viruses, 7, 873–886.
Seybert A, van Dinten L C, Snijder E J, Ziebuhr J. 2000.Biochemical characterization of the equine arteritis virus helicase suggests a close functional relationship between arterivirus and coronavirus helicases.Journal of Virology,74, 9586–9593.
Seybert A, Posthuma C C, van Dinten L C, Snijder E J,Gorbalenya A E, Ziebuhr J. 2005. A complex zinc finger controls the enzymatic activities of nidovirus helicases.Journal of Virology, 79, 696–704.
Shi Y, Li Y, Lei Y, Ye G, Shen Z, Sun L, Luo R, Wang D, Fu Z F, Xiao S, Peng G. 2016. A dimerization-dependent mechanism drives the endoribonuclease function of porcine reproductive and respiratory syndrome virus nsp11.Journal of Virology, 90, 4579–4592.
Snijder E J, Kikkert M, Fang Y. 2013. Arterivirus molecular biology and pathogenesis.Journal of General Virology,94, 2141–2163.
Snijder E J, Meulenberg J J. 1998. The molecular biology of arteriviruses.Journal of General Virology, 79, 961–979.
Tian K, Yu X, Zhao T, Feng Y, Cao Z, Wang C, Hu Y, Chen X, Hu D, Tian X, Liu D, Zhang S, Deng X, Ding Y, Yang L, Zhang Y, Xiao H, Qiao M, Wang B,et al. 2007. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark.PLoS ONE, 2, e526.
Tong G, Zhou Y, Hao X, Tian Z, An T, Qiu H. 2007. Highly pathogenic porcine reproductive and respiratory syndrome,China.Emerging Infectious Diseases, 13, 1434–1436.
Xie J, Cui T, Cui J, Chen Y, Zhang M, Zhou P, Deng S, Su S, Zhang G. 2014. Epidemiological and evolutionary characteristics of the PRRSV in Southern China from 2010 to 2013.Microbial Pathogenesis, 75, 7–15.
Zhang H, Guo X, Ge X, Chen Y, Sun Q, Yang H. 2009. Changes in the cellular proteins of pulmonary alveolar macrophage infected with porcine reproductive and respiratory syndrome virus by proteomics analysis.Journal of Proteome Research, 8, 3091–3097.
Zhang Y, Li H, Peng G, Zhang Y, Gao X, Xiao S, Cao S, Chen H, Song Y. 2015. Mutational analysis of the functional sites in porcine reproductive and respiratory syndrome virus non-structural protein 10.Journal of General Virology, 96,547–552.
Zhang Z, Wen X, Dong J, Ge X, Zhou L, Yang H, Guo X. 2017.Epitope mapping and characterization of a novel nsp10-specific monoclonal antibody that differentiates genotype 2 PRRSV from genotype 1 PRRSV.Virology Journal, 14, 116.
Zhao K, Ye C, Chang X, Jiang C, Wang S, Cai X, Tong G, Tian Z, Shi M, An T. 2015. Importation and recombination are responsible for the latest emergence of highly pathogenic PRRSV in China.Journal of virology, 89, 10712-10716.
Zhou L, Wang Z, Ding Y, Ge X, Guo X, Yang H. 2015. NADC30-like strain of porcine reproductive and respiratory syndrome virus, China.Emerging Infectious Diseases, 21, 2256–2257.
Zhou L, Yang H. 2010. Porcine reproductive and respiratory syndrome in China.Virus Research, 154, 31–37.
Zhou L, Zhang J, Zeng J, Yin S, Li Y, Zheng L, Guo X, Ge X,Yang H. 2009. The 30-amino-acid deletion in the Nsp2 of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China is not related to its virulence.Journal of Virology, 83, 5156–5167.
杂志排行

Journal of Integrative Agriculture的其它文章
- Detection and characterization of an isolate of Tomato mottle mosaic virus infecting tomato in China
- Yield and water use responses of winter wheat to irrigation and nitrogen application in the North China Plain
- A simulation of winter wheat crop responses to irrigation management using CERES-Wheat model in the North China Plain
- Effect of dietary supplementation with flavonoid from Scutellaria baicalensis Georgi on growth performance, meat quality and antioxidative ability of broilers
- ldenti fication of miRNAs and target genes regulating catechin biosynthesis in tea (Camellia sinensis)
- Variability in total antioxidant capacity, antioxidant leaf pigments and foliage yield of vegetable amaranth