水产养殖车间LED光环境设计研究
2018-04-26宋昌斌刘立莉卢鹏志杨华李贤高霄龙陈涛熊巍雷堡乐马贺杜建军
宋昌斌,刘立莉,卢鹏志,杨华,李贤,高霄龙,陈涛,熊巍,雷堡乐,马贺,杜建军
(1.中国科学院半导体研究所,北京100083;2.中国科学院海洋研究所,山东青岛266071;3.深圳市超频三科技股份有限公司,广东深圳518116;4.大连海洋大学水产与生命学院,辽宁大连116023)
在工业化循环水养殖系统中,温度、溶解氧、pH、氨氮、盐度等是影响水生动物行为与生理的重要因素,但光照条件同样重要,目前,国内相关研究领域中,光照对水产动物的影响研究及其在养殖生产中的应用设计水平仍处在起步阶段,并且在对光源的认识与使用方法上存有一定争议。
国内大部分水产养殖车间使用的光照仍然是传统粗放型的白炽灯、日光灯、钠灯等。在设计人工光照方案时,如何选择适合特定水产动物生长的光照参数并在此基础上降低养殖一次性投入成本,一直是困扰水产业的难题。由于LED具有节能环保、光电转换效率高、寿命长、发热低、智能可调控等优点,其在工厂化水产养殖行业中的应用优势日益凸显。本研究中,在参考有关文献资料并结合近几年LED农业应用实践,针对水产养殖车间光照设计时经常遇到的光环境因子优化及控制灯具成本等主要问题,进行了水产养殖车间LED人工光照设计方案研究,以期为提高水产养殖生产效率,促进养殖产业可持续健康发展提供有益的科学依据。
1 设计基础依据
了解水产动物视觉生理结构是设计水产养殖车间LED光照方案的基础依据。
1.1 水产动物的野生环境
自然界除月光、星光等极低的冷光源外,太阳光是主宰万物生灵的自然光源。索饵、繁殖、捕食、逃避敌害、集群等是水产动物在自然环境中的主要行为,而每一种行为均与光照有着密切的联系。水产动物在亿万年的进化过程中,基于环境特点而形成了独特的视觉生理结构,如鱼类通过视网膜与松果体的光感受器接收光信号,以同步感知并适应自然栖息地的光环境。因此,水产动物在自然界中的光环境特征是开展人工养殖条件下光环境因子优化和调控的首要参考要素。
1.2 水产动物视觉生理结构
板鳃鱼类与硬骨鱼类均有发达的眼睛,眼结构与高等脊椎动物基本相似。鱼眼视网膜内层视杆与视锥两种视觉细胞的区别主要是:视杆细胞接纳光线的能力极弱,属低阈值细胞,光敏感性高,无辨色和精细分辨能力;视锥细胞接纳的光波长,属高阈值细胞,有辨色与精细分辨能力[1],但必须在亮光视觉条件下才能起作用[2]。两种细胞光照度阈值均小于1 Lx。大多数板鳃鱼类的视网膜只有视杆细胞,少数星鲨Mustelus manazo等与硬骨鱼类一样有视杆与视锥细胞,而许多深海鱼类的视网膜则完全没有视锥细胞。夜出性水产动物,视网膜完全由视杆细胞组成,而一些昼出性水产动物,视网膜则完全由视锥细胞组成[3]。生活在光照条件较好的水体中依靠视觉捕食的硬骨鱼类,视锥细胞发达,可辨别颜色,光刺激阈值高,敏感性低;而视觉不发达的深海与夜间活动的鱼类,光刺激阈值低,敏感性强,大多数没有辨色能力。
贝类中无板纲、单板纲、多板纲与瓣鳃纲无头眼,而多板纲一部分只有微眼,腹足纲的眼则由皮肤凸陷形成视网膜。头足纲除二腮类眼部结构复杂外,其余眼睛构造简单,通常无柄[4]。其他贝类视力较弱,甚至无眼,只靠皮肤感光。如属于腹足纲的鲍鱼眼部外观形态和结构均较为原始,由单一网状纤维和结缔组织组成,缺少高等动物眼部表层相应的保护、过滤及折射等结构,基于鲍鱼昼伏夜出的节律性运动规律,可初步判断其视网膜内存在视杆细胞,而视锥细胞是否存在还有待进一步研究[5]。海胆、海参等棘皮动物无头无脑,对于光照变化,其可以通过自身调节做出适当的反应。Stubbs[6]和Yoshida等[7]认为,一些棘皮动物通过散布于真皮的感光点对光照做出反应。通过文献资料中关于水产动物等视觉生理结构的分析及试验结果可知,大部分适宜鱼类生长的光照强度要高于贝类、棘皮动物数十倍水平。如Ellis等[8]推荐养殖巴哈马石斑鱼Epinephelus striatus适宜的光强为1636 Lx;王卫民等[9]对中国常见的栉孔扇贝Chlamys farreri等4种扇贝进行了研究,认为暗光有利于幼体的均匀分布,一般控制在500 Lx以下;张辉等[10]认为,10 Lx以下的弱光环境对刺参Apostichopus japonicus的养殖较为适宜;董贯仓[11]对刺参光照试验结果表明,光照为5.18~278 Lx时,随着光照增强,刺参避光行为的比率逐渐增加 (刺参分布比率为43.96%~8.17%)。
1.3 水产动物在不同光色层吸收光波长的差异
水产生物对光的敏感性差别由其栖息地的生态环境所决定,是长期自然选择的结果。各种波长的光在水中被吸收的程度差别较大,红外线最易被海水吸收,其次是红光。随着波长的减少,吸收率也相应下降,短波段的蓝紫光在海水中的穿透力最强。在公海的最初10 m中,红、橙、黄光基本已被吸收[12],可见光中短波段的蓝紫光在海水中的分布可达百米以下,在浮游植物和颗粒物质较多的近岸淡水中,可见光中长波长的光谱占主导地位[13],一般来说,鱼类视网膜所具有的视色素与特定种群所在栖息地光谱的光波峰是一致的。故蓝紫光色层活动的鱼类光谱敏感曲线在短波长范围内,即对蓝光敏感,而活动在近岸海水与淡水中的鱼类吸收的光则偏重于长波长范围内[14]。中华绒螯蟹Eriocheir sinensis的自然天性是在江河湖泊生长至2龄,性腺发育至第Ⅳ期后,向浅海做生殖洄游;在海水孵化出的溞状幼体至大眼幼体阶段对蓝光具有明显的趋光性,发育后期转变为对红光敏感[15]。蓝刻齿雀鲷Chrysiptera cyanea栖息在0~10 m的水面,红光会刺激其性腺发育[16]。珊瑚虫大多栖息在数十米以下的海底,用偏重于蓝色的LED可使人工养殖的丛生盔型珊瑚Galaxea fascicularis较快生长[17]。黑线鳕Melanogrammus aeglefinus常见于40~200 m深处,使用蓝光会显著增加其摄食量[18]。与此类似,在调控欧洲鲈Dicentrarchus labrax、塞内加尔鳎 Solea senegalensis、大西洋鳕Gadus morhual的幼鱼生长发育过程中,也应选择短波长的蓝、绿光,避免使用长波长的红光[19]。
1.4 防止设计光照极端化
尽管光在水中的传输由于复杂的吸收与散射常伴有强烈的衰减,但除在洞穴、深海栖息的物种外,绝大多数的水生生物均具有复杂的视觉生理结构,极少数物种能不靠光而存活下来。养殖车间全天黑暗或全天高强度的光照均不可取[20]。太平洋蓝鳍金枪鱼 Thunnus orientails幼鱼在小于15 Lx光照下,表现出明显的焦虑行为,从而使存活率降低[21]。欧洲鲈Dicentrarchus labrax幼鱼在24 h光照下出现畸形,全天黑暗环境下则孵化率降低,小于30%[19]。因此,设计极端的光环境一定要谨慎,并需有理论与试验依据。
2 自然光谱与人工光谱的区别
2.1 全光谱
游奎等[22]的试验证明,金卤灯照明条件下凡纳滨对虾Litopenaeus vannamei的特定生长率比日光灯连续照明时要高55.89%,并改善体色,他们认为灯具的光谱是导致差异的主要原因,金卤灯的光谱接近太阳光谱 (图1)的程度明显高于日光灯。牛顿于1672年在他的科学论文中记载了将太阳光通过棱镜折射七色光谱的实验,说明白色的光本身是没有的,其主要是由不同颜色的光混合而成[23]。全光谱目前没有严格的物理学定义,主要是指光谱波长覆盖所有可见光区,并有少量紫外光和红外光[24]。人工光源的显色指数高、色温低不能代表该光源是全光谱光源 (图1)。

图1 太阳光谱分布图Fig.1 The solar spectrum
日常环境中较难用肉眼对白光光谱进行区别,而照明使用最多的LED白光是GaN芯片发出蓝光激发黄色荧光粉得到的,其光谱主要是蓝光成分(图2)。在无试验数据基础或无法弄清楚哪种波长的光色对养殖生物有利的情况下,应尽量模拟并使用与水产动物栖息地光照环境特点相似的光源。中国科学院半导体研究所使用3颗不同波长的单色光LED芯片和蓝光LED激发红绿黄三色荧光粉组成的白光LED,利用不同电流控制每种LED输出的功率,组合成白光光谱,实现了连续谱的自然全光谱拟合 (图3),实验室做出的灯具与太阳光光谱达到了较高的保真度[25](图4)。目前,科技工作者正努力制造出与太阳光拟合度高的人工光源灯具。
2.2 LED色温低与高显色性的白光并非自然白光
目前,LED发出的白光大部分以InGaN蓝色LED芯片为基础光源激发稀土石榴石 (YAG)黄色荧光粉的混色方法制成,这种方法工艺成熟、简单、成本低。从LED发光光谱上分析,由于此种混色方案中,蓝光光谱为主峰,而红光光谱偏少,使得此类白光LED色温只能是5000 K以上的高色温、低显色性的光源。如果增加氮化物荧光粉成分,则可以降低色温,提高显色指数,但光视效率随之降低。总之,无论色温低或显色指数高的白光均不是自然太阳光谱 (图1)。
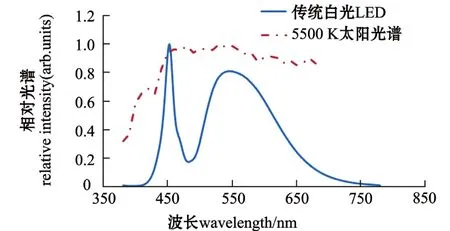
图2 白光LED光谱与5500 K太阳光谱对比图Fig.2 Comparison of white light LED spectrum with 5500 K solar spectrum

图3 中国科学院半导体研究所全光谱与太阳光谱拟合图Fig.3 Fitting of complete spectrum and the solar spectrum from Institute of Semiconductors,Chinese Academy of Sciences(CAS)

图4 中国科学院半导体研究所全光谱光源分布图Fig.4 Complete spectrum diagram of Institute of Semiconductors, CAS
3 水产养殖车间光环境的设计
3.1 光周期调控的作用
许多生物依赖于周年光周期的变化而生长、发育和繁殖,对鱼类而言,昼长是一个重要的 “授时因子”。研究表明,光周期对宝石蓝雀鲷Chrysiptera springer、长须鲸 Balaenoptera physalus、大西洋鲑Salmo salar等的生长和繁殖均具有重要影响[16,26-27]。在生产中, 通过光周期调控可以加速或延缓许多生物的性腺启动时间,例如大西洋鲑、虹鳟Oncorhynchus mykiss、欧洲鲈、金头鲷、大西洋鳕等[28]。在养殖生产过程中,如能应用光周期进行调控则最为简单,而又不会增加成本。根据养殖生物的特性,LED智能控制系统可实现相应的光周期调控,但突然开、关照明灯具会使水产动物产生光胁迫应急反应,增加死亡率。太平洋金枪鱼Thunnus thynnus在突然断电灭灯时鱼群会发生剧烈碰撞[21]。研究表明,灯具的开与关,应以逐亮、渐灭的方式完成,可明显降低死亡率。
3.2 光照调控的实现
智能光照管理系统包括集中管理系统、网关设备、调光控制器、电源与光源。运用通信网络接口协议ZigBee、PLC、RS485、以太网和 GPRS等多种物联网及IT技术,可实现单灯开关、调光、监测计量等管理功能,但每个协议各有优缺点。PLC是利用现有电力线,通过载波方式传输信号[29];RS485采用双绞线屏蔽电缆传输差分信号;而Zig-Bee则是经常采用低速、低功耗无线控制协议,特色是可以自动组成网络,网络的每个节点可以借力传输数据,但网络中需要一个集中节点来管理整个网络,也就意味着ZigBee网络中必须有一个类似路由器的角色,完成ZigBee协议到互联网协议的转换。无论哪个通信协议,每个灯必须安装对其发出指令的硬件调光管理装置,明显不足之处就是增加了成本和安装的复杂度。如果仅给光源渐亮、渐灭控制功能,则可以建立独立触摸屏和独立循环控制系统。智能触控屏给调光控制模块发送场景或回路调光指令,输出1~10 V调光信号给LED灯具配套的调光驱动,LED根据不同的信号值呈现不同的亮度,从而实现LED灯亮度渐亮或渐暗变化。
3.3 人工光环境设计与成本的核算
LED光源在水产车间使用的最大障碍是灯具成本问题,而决定灯具成本的首要因素是光源的电功率。目前,市场上的大功率灯具为5~8元/W,即百瓦的灯具,价格在1千元左右。所以针对特定养殖生物选择合适的光源电功率是降低成本的关键因素。
3.3.1 优选光色以降低成本 Migaud等[30]发现,大西洋鳕幼鱼在3种不同光谱和3种不同光强条件下,短波长光谱对促进其生长的贡献明显优于光强。所以在光强与光谱均可起到同样作用的条件下,尽量优选光谱,从而降低灯具功率,减少成本。
3.3.2 选下限阈值的光强以节省成本 水产动物有极其庞大的种群,研究表明,每种鱼对光的反应均有差异,但绝大部分鱼类生长所需要的光照都有最小临界值。只有极少数深海底栖或浑浊水体中的鱼类在生长发育过程中只需低于1 Lx的光。Honryo等[21]试验证明,夜晚当光强超过150 Lx时,太平洋金枪鱼幼鱼成活率升高 (可达75.8%),死亡率大幅度降低。如果增大光强并不能有效提高成活率,在灯具设计时不妨取此下限阈值150 Lx。
3.3.3 避免长时间使用低功率驱动的灯具 不同种类的水生生物对光的反应有差异,即便同一物种在其不同生长发育阶段对光照的需求也可能不同[19]。有些养殖生产者要求灯具有普适性,于不同种或同一种鱼不同生长阶段均可使用,这就要求必须智能控制调光,光强变化为1%~100%。生产商可提供此类灯具,但问题在于,若一只100 W的灯具,经常被设置并使用50%的功率驱动,相当于50 W灯具以额定功率工作,但一次性成本投入必须是100 W灯具的价格,这种设计显然不可取。养殖实践中,建议订制若干个50 W功率灯具,备用少量50 W功率灯具。短时间需要加大光照时,可以考虑使用备用灯具,增加辐射照度。
3.3.4 光环境模拟 模拟光强的前提条件是依据白色光谱的计算机软件进行照明计算。目前,在市场上比较流行的十几种设计软件中,水产车间比较实用的是DIALux,这款软件用于封闭空间照明设计,易学易用,引入数据简便,系统引入光度数据开放,结果评估准确,光照场景渲染输出直观、真实[31]。例如,一个边长为4 m的正方形养殖池,在其正上方2.9 m处悬挂IP65防水等级、整灯光效70 Lm/W的18 W三防LED灯具。软件模拟空间形象彩图与工作面光强分布曲线图如图5、图6所示。
模拟参数表明:水池水面平均光强为28 Lx,最小光强为10 Lx,最大光强为62 Lx,最小值/平均照度为0.36,维护系数为0.80。基于养殖生物需要的光强和均匀度,可通过与模拟结果对比调整灯具的功率与LED的光束角及透镜。从模拟的最小值/平均照度值可知均匀度,一般来说,游泳性水产动物对光的均匀度要求并不严格,但趋光性强的水产动物由于长时间聚集在一起,局部溶氧度降低易导致危害,此种情况下要求灯具的配光曲线光束扩散角要大一些,光强分布尽可能均匀。

图5 模拟空间伪彩图Fig.5 Simulation of pseudo color space
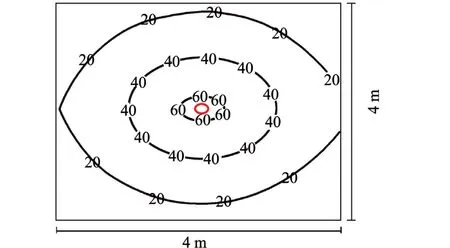
图6 模拟光强分布等照度图Fig.6 ISO intensity diagram of simulated intensity distribution
3.3.5 灯具选型 水产光照主要选择的LED灯型有球泡灯、线型管灯、工矿灯、投光灯等。前两种功率较小,后两种功率较大,这些灯具的寿命基本均可达到3万小时以上。防护等级须不低于IP65,同时要考虑整灯的发光效率,目前,白光大功率整灯理想的发光效率在80 Lm/W以上。无论使用线型灯还是点光源灯型,主要考虑的是结合养殖池几何形状观察灯具的配光曲线。何时选择球泡灯、线型管灯主要是从成本角度考虑,因为这两种灯具是全球室内功能性照明的主力灯型。因为生产量巨大,所以其成本相对较低。但若从防水角度考虑,
球泡灯的灯头须为密闭防水,T8或T5管需要加上透明的PC三防罩。如果需要特殊外形的灯具,可通过新开模具、设计制作电路板等工序由生产厂家生产出所需产品。
4 一个水产车间光环境设计实例
大连富谷水产有限公司黄条鰤车间共有130个养殖池,每个池子面积为56 m2。车间光环境由中国科学院半导体研究所设计,深圳市超频三科技股份有限公司负责灯具与控制系统制造与现场施工。自然环境黄条鰤Seriola aureovittata属中上层温水性远洋洄游鱼类[32],使用深圳超频三科技股份有限公司FG-2型60 W投光灯,用DIALux软件模拟,平均照度为126 Lx,均匀度为0.213。安装后的效果基本与设计模拟结果相符。
智能控制运用通信网络协议ZigBee实现。节点控制器通过1~10 V调光线控制电源输出,网关通过ZigBee协议与节点控制器实现通讯。平板电脑通过wifi实现与网关通讯,且平板电脑 (服务器)按养殖方需要实行渐亮渐灭调控,以及光强和光周期灯光调控。
5 结语
从全球半导体照明发展趋势分析,LED照明技术正向高光效、高可靠性、智能化和模块化等方向发展,先进的LED光源应用潜力巨大。基于水生生物的视觉特性和对光照条件敏感性的差异,使得人工调控光照并建立适宜不同物种以及同一物种不同生长发育阶段需要的特定光照条件成为必然。随着LED在水产光照领域应用研究的逐步深入,智能调控、尺寸灵活、安装便利、成本低廉的LED设计与制造工艺也会持续改进。
参考文献:
[1] 李明德.鱼类形态与生物学[M].厦门:厦门大学出版社,2011:87.
[2] 殷名称.鱼类生态学[M].北京:中国农业出版社,1995:153-154.
[3] 林浩然.鱼类生理学[M].广州:中山大学出版社,2011:460-461.
[4] 蔡英亚,张英,魏若飞.贝类学概论[M].上海:上海科学技术出版社,1995:50-87.
[5] 高霄龙.光照对皱纹盘鲍生长、行为、生理的影响及其机制研究[D].青岛:中国科学院研究生院(海洋研究所),2016:43.
[6] Stubbs T R.The neurophysiology of photosensitivity in ophiuroids[C]//Lawrence J M.Echinoderms:Proceedings of the International Conference,Tampa Bay.Rotterdam:A A Balkema,1982:403-408.
[7] Yoshida M,Takasu N,Tamotsu S.Photoreception in echinoderms[M]//Ali M A.Photoreception and Vision in Invertebrates.Boston,MA:Springer,1984:743-771.
[8] Ellis E P,Watanabe W O,Ellis S C,et al.Effects of turbulence,salinity,and light intensity on hatching rate and survival of larval Nassau grouper,Epinephelus striatus[J].Journal of Applied Aquaculture,1997,7(3):33-43.
[9] 王卫民,温海深.名特水产动物养殖学[M].2版.北京:中国农业出版社,2017:69.
[10] 张辉,王印庚,荣小军,等.刺参的趋光性以及对附着基颜色的感应行为[J].生态学杂志,2009,28(3):477-482.
[11] 董贯仓.光照及投礁方式对刺参(Apostichopus japonicus)行为生长的影响及其机制[D].青岛:中国海洋大学,2009:31.
[12] 俞文钊.鱼类趋光生理[M].北京:农业出版社,1980:9.
[13] McFarland W N.The visual world of coral reef fishes[M]//Sale P F.The Ecology of Fishes on Coral Reefs.San Diego,CA:Academic Press,1991:16-38.
[14] Yeh N,Yeh P,Shih N,et al.Applications of light-emitting diodes in researches conducted in aquatic environment[J].Renewable and Sustainable Energy Reviews,2014,32:611-618.
[15] 陈明卫,高维玉,姜玉声,等.光色对中华绒螯蟹幼体诱集与仔蟹摄食的影响[J].大连海洋大学学报,2016,31(4):362-367.
[16] Bapary M A J,Fainuulelei P,Takemura A.Environmental control of gonadal development in the tropical damselfish Chrysiptera cyanea[J].Mar Biol Res,2009,5(5):462-469.
[17] Wijgerde T,Henkemans P,Osinga R.Effects of irradiance and light spectrum on growth of the scleractinian coral Galaxea fascicularis applicability of LEP and LED lighting to coral aquaculture[J].Aquaculture,2012,344-349:188-193.
[18] Downing G,Litvak M K.The effect of light intensity and spectrum on the incidence of first feeding by larval haddock[J].J Fish Biol,2001,59(6):1566-1578.
[19] Villamizar N,Blanco-Vives B,Migaud H,et al.Effects of light during early larval development of some aquacultured teleosts:a review[J].Aquaculture,2011,315(1-2):86-94.
[20] Boeuf G,Le Bail P Y.Does light have an influence on fish growth?[J].Aquaculture,1999,177(1-4):129-152.
[21] Honryo T,Kurata M,Okada T,et al.Effects of night-time light intensity on the survival rate and stress responses in juvenile Pacific bluefin tuna Thunnus orientalis(Temminck and Schlegel)[J].Aquaculture Research,2013,44(7):1058-1065.
[22] 游奎,杨红生,刘鹰,等.不同光源及光照时间对凡纳滨对虾(Litopenaeus vannamei)游离虾青素含量及生长的影响[J].海洋与湖沼,2005,36(4):296-301.
[23] 倪光炯,王炎森.物理与文化[M].2版.北京:高等教育出版社,2009:106.
[24] 李琪,辛易.全光谱LED发展现状及应用前景[J].中国照明电器,2017(3):12-16.
[25] Lu P Z,Yang H,Pei Y R,et al.Generation of solar spectrum by using LEDs[C]//Fifteenth International Conference on Solid State Lighting and LED-based Illumination Systems.San Diego,California,United States:SPIE,2016,9954:99540N.
[26] Bromage N,Porter M,Randall C.The environmental regulation of maturation in farmed finfish with special reference to the role of photoperiod and melatonin[M]//Lee C S,Donaldson E M.Reproductive Biotechnology in Finfish Aquaculture.Amsterdam:Elsevier,2001:63-98.
[27] 仇登高,徐世宏,刘鹰,等.光环境因子对循环水养殖系统中大西洋鲑生长和摄食的影响[J].中国水产科学,2015,22(1):68-78.
[28] Skulstad O F,Taylor J,Davie A,et al.Effects of light regime on diurnal plasma melatonin levels and vertical distribution in farmed Atlantic cod(Gadus morhua L.)[J].Aquaculture,2013,414-415:280-287.
[29] 周太明.照明设计——从传统光源到LED[M].上海:复旦大学出版社,2015:187.
[30] Migaud H,Davie A,Carboni S,et al.Effects of light on Atlantic cod(Gadus morhua)larvae performances:focus on spectrum[C]//Symposium:Special Publication,European Aquaculture Society.Ghent,Belgium:[s.n.],2009:265-269.
[31] 李农.照明设计师[M].北京:中国劳动社会保障出版社,2009:1-130.
[32] 李荣,徐永江,柳学周,等.黄条鰤(Seriola aureovittata)形态度量与内部结构特征[J].渔业科学进展,2017,38(1):142-149.
