云南蝗不同地理种群染色体研究
2018-04-25欧晓红
柳 青 欧晓红
(1. 云南农业大学植物保护学院,云南 昆明 650201;2. 西南林业大学林学院,云南省森林灾害预警与控制重点实验室,云南 昆明 650224)
染色体作为生物遗传物质的载体,在各生物类群中,染色体的数目、大小、形态、组型、减数分裂行为及异染色质分布等特征都具有相对稳定性[1-2]。因此,与形态学数据一样,染色体特征数据已经成为生物鉴别分类和亲缘关系界定的基本数据之一[3]。通过研究和分析不同生物类群染色体特征,可以解决基于形态分类中不能解决的许多疑难问题。
云南蝗 (Yunnanitescoriacea) 隶属直翅目 (Orthoptera) 蝗总科 (Acridoidea) 瘤锥蝗科 (Chrotogonidae) 云南蝗属 (Yunnanites),为我国特有种,主要分布于我国西南地区的云南、贵州和四川等地[4]。前期野外调查时在云南省昆明西山和东川分别采到云南蝗标本,经过形态学比较发现,2个种群云南蝗在个体大小、触角长短和后足股节下膝侧片颜色3个方面表现出明显差别。因此,本实验采用常规的液氮冰冻压片法及BSG显带法对云南蝗2个种群的染色体核型和C带带型进行研究,以期进一步明确云南蝗不同种群的染色体特征,为云南蝗细胞学研究提供参考资料。
1 材料与方法
1.1 实验材料
云南蝗标本均采自野外,具体采集信息见表1。

表1 云南蝗标本来源及研究个体数量Table 1 Specimen source and number of studied individuals of Y.coriacea
1.2 染色体玻片制备
将采集的云南蝗雄虫按个体编号,依据个体大小不同,活体腹腔注射0.05%的秋水仙素溶液4~6 μL,经6~8 h后解剖取出精巢,放入0.08%的生理盐水中低渗10~15 min,再浸入甲醇冰醋酸 (体积比3∶1) 固定液内固定6~12 h,然后移入75%酒精内常温保存[5]。制片时取出预先保存好的精巢材料,在体视解剖镜下用解剖针挑取精小管盲端部分2~3根置于洁净的载玻片上,滴加45%冰醋酸软化5~10 min,盖上盖玻片进行压片,随后置于液氮罐中冰冻3~5 min,取出后快速揭片,在室温下无尘处自然老化2~6 d。
1.3 C带核型处理
C带核型处理采用BSG法进行。将老化好的玻片置于5%新配制的饱和Ba (OH)2溶液中50 ℃恒温水浴处理15~20 min,取出后用同温度的蒸馏水冲洗,再经2 × SSC于60 ℃恒温水浴处理1 h,用同温度的蒸馏水冲洗晾干,5% Giemsa染液 (pH 6.8) 染色10~15 min,蒸馏水冲洗,自然干燥,镜检[6-7]。使用Nikon ECLIPSS E800进行显微拍照,每种选取5~10个染色体形态清晰、分散良好的分裂相细胞进行染色体数目统计、C带核型参数的测量和计算,并用Adobe Photoshop CS6软件对染色体图像切割配对,染色体类型依据Levan等[8]的标准进行划分。
2 结果与分析
2.1 昆明西山猫儿箐种群的染色体特征
2.1.1昆明西山猫儿箐种群的染色体核型
云南蝗昆明西山猫儿箐种群染色体核型见图1,可知其染色体数目为2n(♂)=19,染色体臂指数NF=19;染色体类型为端着丝粒染色体;性别决定机制均为XO型。该种群有丝分裂中期染色体数据详见表2,其中染色体按照相对长度 (RL) 可以分为3组:L组 (大型染色体) 3对 (L1~L3),RL值为10.33~13.06;M组 (中型染色体) 6对 (M4~M9),RL值为6.49~9.85;无小型染色体;性染色体X属大型染色体,RL值为16.16,其相对长度位居第1 (图1, 表2)。因此其染色体组式为3L + 6M + X。
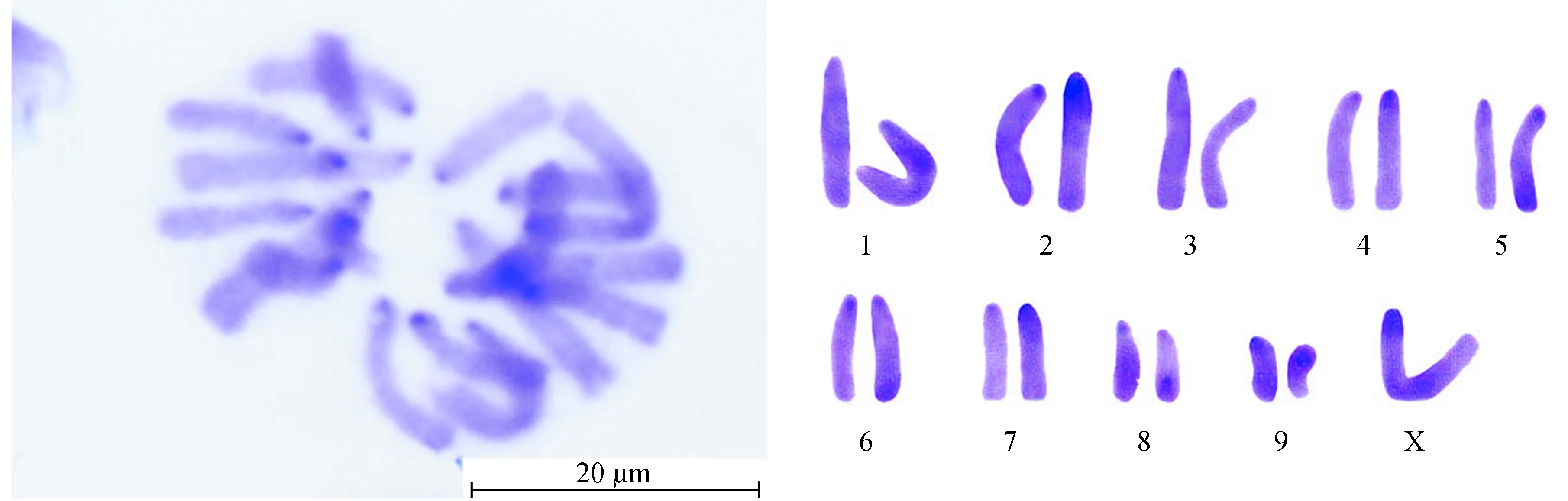
图1云南蝗昆明西山猫儿箐种群染色体核型
Fig.1 Karyotype ofY.coriaceapopulation from Maoerqing
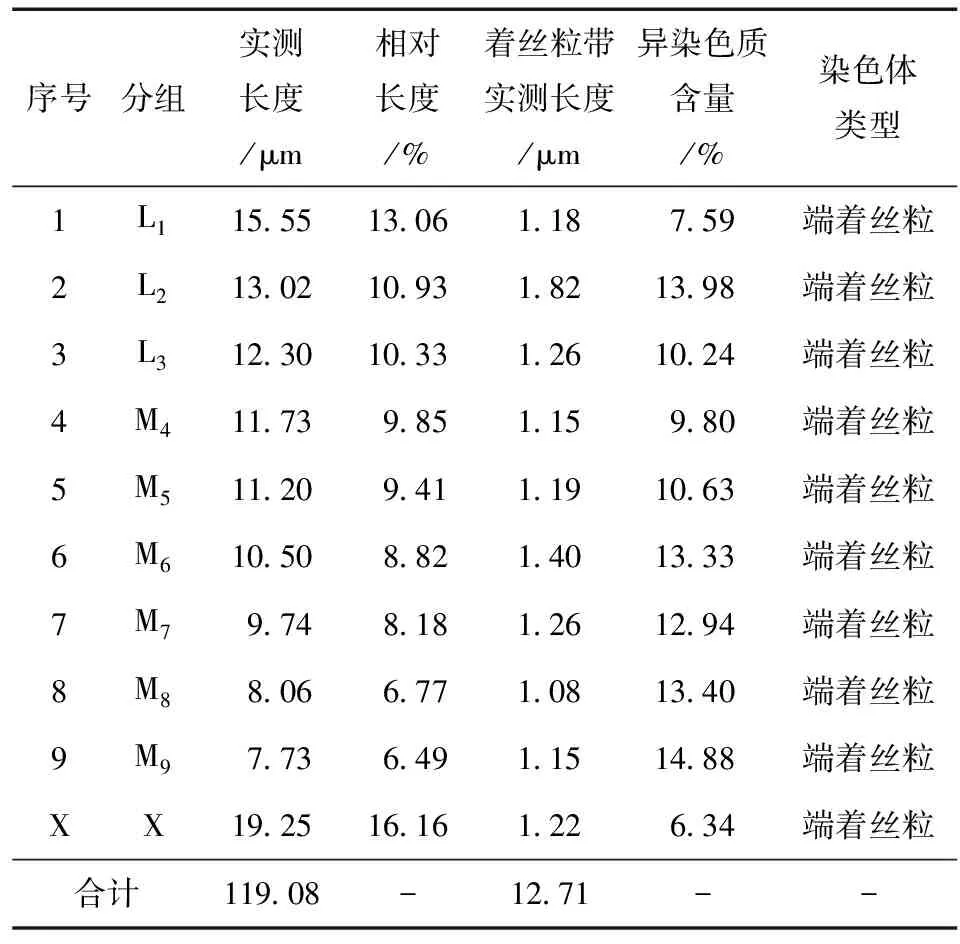
表2 云南蝗昆明西山猫儿箐种群有丝分裂中期染色体数据Table 2 Mitotic metaphase chromosome data of Y.coriacea population from Maoerqing
2.1.2昆明西山猫儿箐种群的C带带型
由图2可知,云南蝗昆明西山猫儿箐种群染色体C带带型单一,每条染色体仅具有着丝粒带,而无其他带型。异染色质在整个染色体组中的总含量为10.67%,M9染色体异染色质含量最高,为14.88%,L2次之,其异染色质含量为13.98%,性染色体X的异染色质含量在整个染色体组中最低,为6.34% (表2)。
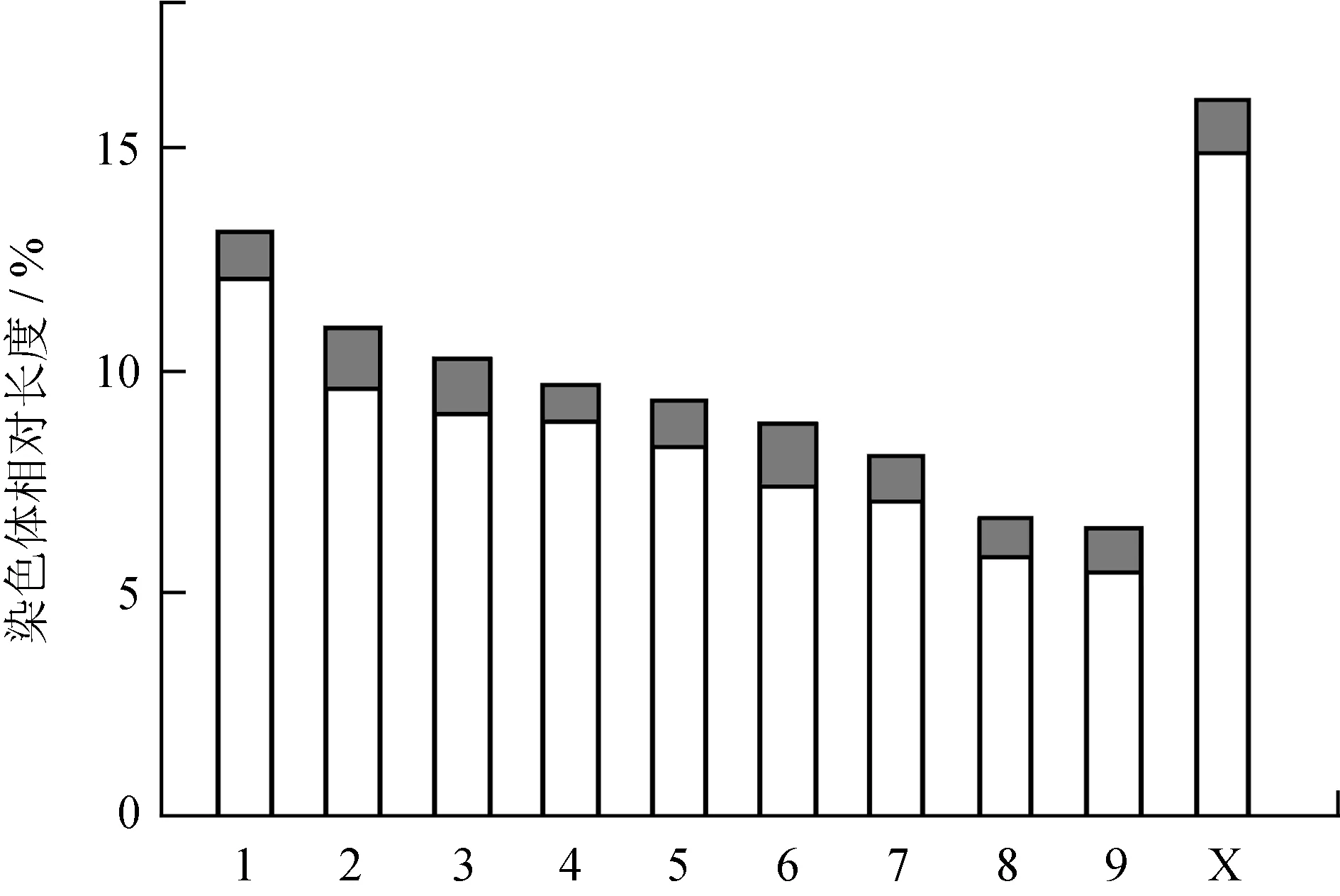
图2云南蝗昆明西山猫儿箐种群染色体C带核型模式图
Fig.2 Chromosome C-banding karyotype pattern ofY.coriaceapopulation from Maoerqing
2.2 昆明东川拖布卡种群的染色体特征
2.2.1昆明东川拖布卡种群的染色体核型
云南蝗昆明东川拖布卡种群染色体核型见图3。可知其染色体数目为2n(♂)=19,染色体臂指数NF=19;染色体类型为端着丝粒染色体;性别决定机制均为XO型。该种群有丝分裂中期染色体数据详见表3,其中染色体按照相对长度可以分为3组:L组 (大型染色体) 3对 (L1~L3),RL值为10.37~13.52;M组 (中型染色体) 6对 (M4~M9),RL值为6.48~9.40;无小型染色体;性染色体X属大型染色体,RL值为18.47,其相对长度位居第1 (图3, 表3)。因此其染色体组式为3L + 6M + X。
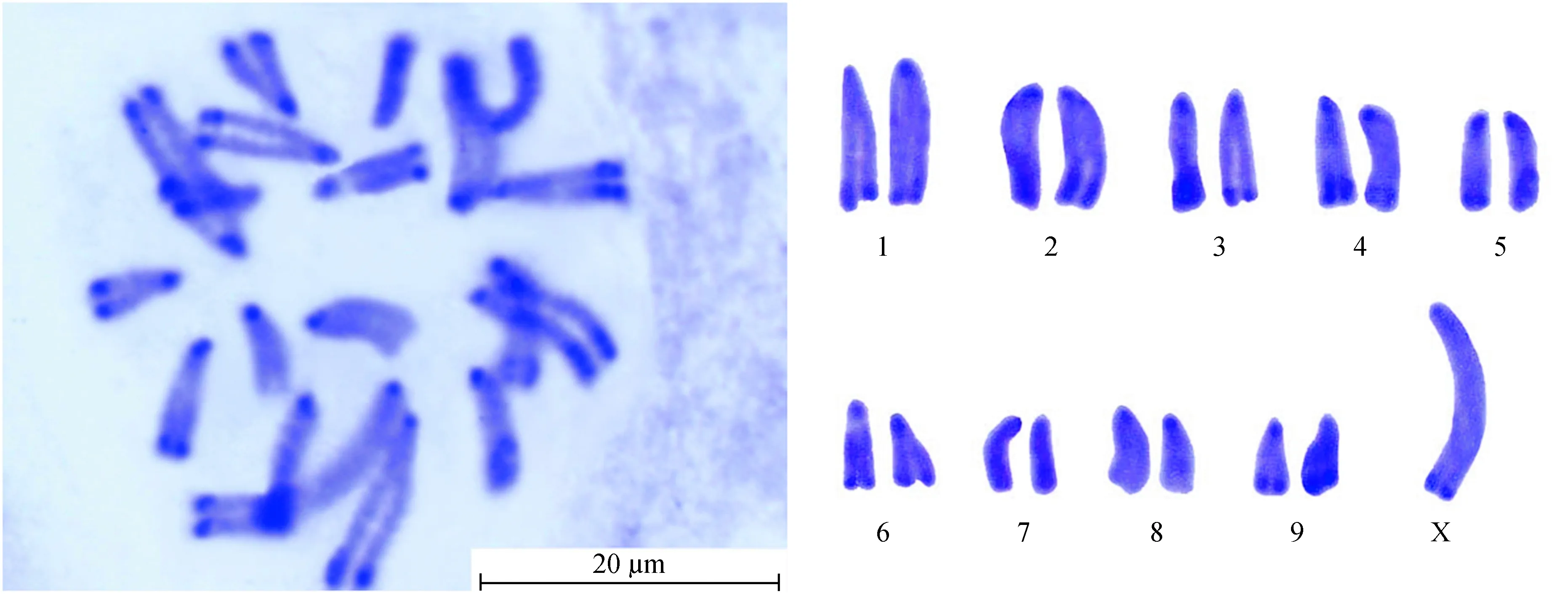
图3云南蝗昆明东川拖布卡种群染色体C带核型
Fig.3 C-banding karyotype ofY.coriaceapopulation from Tuobuka
2.2.2昆明东川拖布卡种群的C带带型
由图4观察发现,昆明东川拖布卡种群具有着丝粒带和端带,且带纹条数较多,除10条着丝粒带外,端带8条分别位于L1、L2、L3、M4、M5、M6、M9和性染色体X上,该种群的异染色质总含量占29.53%。由表3可知:在染色体组中,M5染色体异染色质含量最高,为46.42%;M9染色体次之,其异染色质含量为42.71%;M7染色体的异染色质含量在整个染色体组中最低,为14.87%;性染色体X的异染色质含量为15.09%。
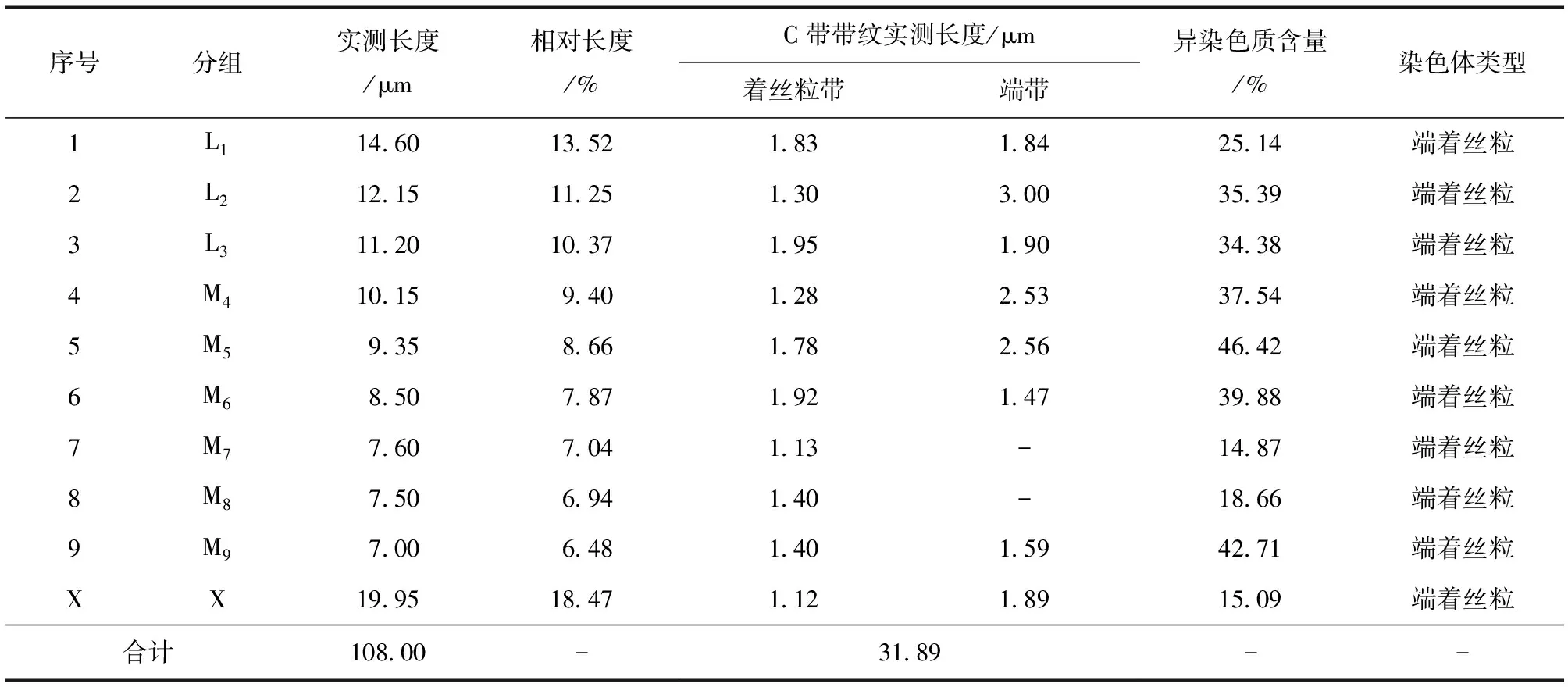
表3 云南蝗昆明东川拖布卡种群有丝分裂中期染色体数据Table 3 Mitotic metaphase chromosome data of Y.coriacea population from Tuobuka
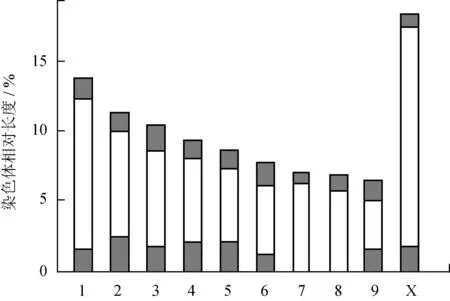
图4云南蝗昆明东川拖布卡种群染色体C带核型模式图
Fig.4 Chromosome C-banding karyotype pattern ofY.coriaceapopulation from Tuobuka
2.3 2种种群染色体特征对比分析
大量的研究表明,基于染色体数据的细胞分类是传统形态分类的很好补充。染色体的数目、组式可以作为科级或属级阶元的分类指标,而染色体C带异染色质分布位置、大小、数量、异染色质总含量及性染色体X的位次等可以作为属、种及种下阶元的分类指标[2,9-11]。从2个种群的染色体特征来看,云南蝗昆明西山猫儿箐种群和昆明东川拖布卡种群染色体特征具有较高的相似性,两者在常规染色体核型包括染色体数目、类型、臂指数、组式、性别决定机制及性染色体位次等方面均表现出高度一致,反映出两者染色体特征在种内水平上的稳定性;两者的差异主要表现在染色体C带带型和异染色质总含量方面,前者C带带型相对单一,每条染色体仅具有着丝粒带,而无其他带型,异染色质总含量为10.67%,而后者具有着丝粒带和端带,带纹条数较多,除10条着丝粒带外还具有8条端带,异染色质总含量明显高于前者,为29.53%,反映出染色体C带核型用于种及种下阶元鉴别分类的重要作用。
3 结论与讨论
前期的野外调查发现,云南蝗昆明西山猫儿箐种群和昆明东川拖布卡种群在外部形态上表现出3点明显差异:1) 昆明西山猫儿箐种群个体较东川拖布卡种群个体明显小;2) 东川拖布卡种群的触角明显较前者的长;3) 后足股节下膝侧片的颜色不同,昆明西山猫儿箐种群的为绿色,而东川拖布卡种群的为淡红色。再结合本研究云南蝗2个种群在染色体C带带型和异染色质总含量方面的差异,建议可将云南蝗2个地理种群视为2个不同的地理亚种。
翅对昆虫的觅食、求偶、迁移和扩散等方面都具有十分重要的作用,昆虫分布生境发生变化,昆虫的翅也会呈现出相应的适应性变化[12]。现有的研究表明,随海拔升高,蝗虫的翅退化强烈[12-13]。云南蝗前翅柳叶状,侧置,仅达第2腹节背板,为典型的短翅型蝗虫种类,因此,其迁移扩散能力弱,这在很大程度上限制了云南蝗不同种群之间的基因交流,从而推动不同地理种群间物种分化的形成。本研究仅研究了云南蝗2个地理种群的染色体C带核型,今后应在更大范围采集不同地区的云南蝗标本,结合外生殖器解剖和DNA分子标记等技术方法对云南蝗不同种群进行更为深入细致的研究。
[参考文献]
[1] 张礼生, 张青文, 蔡青年, 等. 中国昆虫染色体研究现状与展望[J]. 昆虫学报, 2003, 46(6): 773-782.
[2] 马恩波, 欧晓红, 乔格侠, 等. 蝗总科染色体研究及科级综合比较 (直翅目) [J]. 昆虫分类学报, 2000, 22(1): 6-10.
[3] 郝锡联, 闻鸣, 张雪, 等. 4种鸣虫染色体核型研究 (直翅目: 蝗总科) [J]. 吉林农业大学学报, 2016, 38(1): 26-31.
[4] 郑哲民. 蝗虫分类学[M]. 西安: 陕西师范大学出版社, 1993: 42-48.
[5] 柳青, 欧晓红. 斑腿蝗科两种短翅蝗虫的染色体研究[J]. 西南林学院学报, 2005, 25(2): 5-7.
[6] Lin C, Wang N, Gao S J, et al. Comparative study of the karyotypes of two species of grasshoppers in genera Chorthippus Fieber and Euchorthippus Tarbinsky (Orthoptera: Acridoidea) [J]. Entomotaxonomia, 2015, 37(1): 1-7.
[7] 许姝娟, 史红全, 韩亚鹏, 等. 辽宁雏蝗不同地理种群染色体研究[J]. 安徽农业科学, 2017, 45(13): 13-15.
[8] Levan A, Fredga K, Sandberg A A. Nomenclature for centromeric position on chromosomes[J]. Hereditas,1964, 52(2): 201-220.
[9] 李新江, 张道川, 王文强. 蝗虫染色体C-带核型研究进展 (昆虫纲: 直翅目) [J]. 河北大学学报 (自然科学版), 2004, 24(3): 332-336.
[11] Liu Q, Ou X H, Ge H J. C-banding karyotypes of two Caryanda (Orthoptera: Catantopidae) species with short wings from China[J]. Entomological News, 2012, 122(1): 1-9.
[12] 印象初. 青藏高原的蝗虫[M]. 北京: 科学出版社, 1984.
[13] 张红玉, 欧晓红. 西双版纳、滇西北和西藏高原斑腿蝗科区系比较与起源探讨[J]. 云南地理环境研究, 2005, 17(4): 15-19.
