宝天曼马尾松和槲栎混交林的竞争关系分析
2018-04-25史训旺叶永忠袁志良房会普刘晓静
陈 诗 海 鑫 史训旺 叶永忠 袁志良 房会普 刘晓静 王 婷
(1. 河南农业大学林学院,河南 郑州 450002;2. 河南宝天曼国家级自然保护区管理局,河南 南阳 474350)
在森林生态系统中,树木竞争是一种十分常见而又复杂的现象。具有相似生态要求的物种在资源的需求和获取资源的手段上竞争十分激烈,一个种群的增长会导致其他种群所需的环境资源不足甚至受到抑制[1]。竞争不仅影响林木的个体生长以及种群结构,而且在物种进化、种群的空间分布及动态、群落的演替过程中扮演着十分重要的角色[2],是生态学研究的核心问题之一。林木竞争中,一个种群不仅会受到同类种群密度压力导致的种内竞争,还会受到不同种群为争夺环境资源而产生的种间竞争[3]。为了将植物之间的竞争关系量化,自20世纪60年代以来,有很多学者先后提出了针对不同物种的多种竞争指数系统[4-6],竞争指数的实质反映了植物对环境资源的需求与现实生境下植物对环境资源占有量之间的矛盾[7]。引起林木种内种间竞争作用的机制十分复杂,研究林木竞争规律对于了解群落结构与功能,预测群落的发展动态,进而辅以必要的人工管理措施以促进森林生态系统功能的完善具有重要的理论和现实意义[8]。
马尾松 (Pinusmassoniana) 是我国亚热带地区主要树种之一,研究区宝天曼国家级自然保护区位于秦岭东段,处于南暖温带向北亚热带的过渡地区,南北植物区系在此地交汇融合,也是马尾松水平分布的北界,在我国森林生态系统研究中占有重要地位。马尾松-槲栎 (Quercusaliena) 混交林是该保护区内分布较广、具备典型地带性的群落之一[9],在维护区域生态平衡、涵养水源以及保护生物多样性等方面起着重要作用[10]。目前有关宝天曼松栎混交林的研究主要集中于生物量和净生产力[11]、种群空间格局[12]、物种多样性与稳定性[13]、土壤空间异质性[14]以及种-面积关系[15]等方面,而对马尾松混交林的种群径级结构以及对种群动态产生重要影响的种内和种间竞争规律等方面的研究较少。本试验以宝天曼马尾松-槲栎混交林为研究对象,研究主要物种的径级结构特征,并运用Hegyi[16]的单木竞争指数模型,揭示马尾松的种内和种间竞争规律,有利于探讨马尾松-槲栎混交林群落的演替进程及形成机制,以期为保护区森林生物资源保护与管理提供借鉴。
1 研究区概况
河南省宝天曼国家级自然保护区地处东经111°53′~112°04′,北纬33°25′~33°33′,位于秦岭东段的伏牛山南坡,山势呈西北-东南走向,区内最高峰海拔1 830 m,保护区总面积约100 km2,属季风型大陆性气候,四季分明,年平均气温15.1 ℃,年降雨量885.6 mm,年蒸发量991.6 mm,相对湿度68%,极端最高温41.2 ℃,极端最低温-14.8 ℃[17]。保护区地处南暖温带向北亚热带过渡区,植被属暖温带落叶阔叶林向亚热带常绿阔叶林的过渡型,植被分布随海拔变化明显[18]。保护区内物种丰富,植被覆盖率高达95%,群落结构复杂,是我国同纬度地区原始森林生态系统中保存较为完整、物种最丰富的地区之一[12]。马尾松-槲栎混交林是保护区内典型的地带性群落,主要乔木树种有马尾松、槲栎、红皮云杉 (Piceakoraiensis)、麻栎 (Quercusacutissima) 等,马尾松为优势种。灌木主要以连翘 (Forsythiasuspensa)、山胡椒 (Linderaglauca) 等为主,草本植物以红根草 (Salviaprionitis) 和龙牙草 (Agrimoniapilosa) 等为主。土壤主要可划分为山地棕壤、山地黄棕壤和山地褐土,土壤pH 6.5左右[19]。
2 研究方法
2.1 样地调查
2016年7月在宝天曼野獐保护站马尾松-槲栎混交林典型分布区,设置一个1 hm2(100 m × 100 m) 固定样地,将样地划分为小样方 (5 m × 5 m),以样地西南角为原点,东西方向为x轴,南北方向为y轴,对样地内所有胸径 (DBH)≥ 1 cm的立木进行每木检尺,记录物种名、坐标 (x,y)、胸径、树高、冠幅等信息。
2.2 竞争范围确定
根据样地内马尾松径级分布和其个体的空间位置分布确定对象木。参照刘方炎等[20]研究,确定竞争范围,以树冠重叠法为依据。该样地马尾松-槲栎混交林中,马尾松的树冠最大宽度为10 m左右,因此,将竞争范围确定为10 m。以每株对象木为圆心,10 m半径范围内所有调查木本为竞争木,根据对象木和竞争木的坐标位置计算二者之间的距离。
2.3 竞争指数确定
竞争指数是树木间相互作用对树木生长影响的量化,很多学者提出过多种不同类型的竞争指数模型,并将其应用于实际的林木竞争研究中[3]。Hegyi单木竞争指数在计算过程和实践应用方面具有明显优势[21]。本文应用Hegyi指数,以对象木和竞争木之间的距离及胸径为参数进行计算,计算公式如下:
(1)
式中:CI为竞争强度,N为竞争木的株数,Di为对象木的胸径大小,Dj为竞争木的胸径大小,Lij为对象木与竞争木之间的距离。计算对象木马尾松种内和种间的竞争强度,先计算出每株竞争木对对象木的竞争指数,然后累加N株竞争木的竞争强度[5]。CI值越大,竞争强度越激烈。
2.4 数据处理
利用Excel 2013计算单木间的竞争指数,采用SPSS 21.0对马尾松的胸径和竞争指数进行回归拟合。
3 结果与分析
3.1 马尾松-槲栎混交林的林木竞争情况
表1为马尾松和竞争木的胸径分布情况。在宝天曼马尾松-槲栎混交林中,共计调查对象木马尾松61株,最小胸径2.51 cm,最大胸径38.50 cm,平均胸径20.51 cm。径级结构是衡量植物群落稳定性以及林木生长发育状况的重要指标[22]。将对象木马尾松按径级分组,集中在第V至第VII径级,占对象木总数的68.9%,表明马尾松-槲栎混交林中马尾松大树较多,幼树较少,且随着径级的增大,对象木马尾松个体数量逐渐增多。调查的竞争木共1 581株,主要集中在第I至III径级,占竞争木总数的78.9%,说明竞争木中幼龄树较多。所有竞争木中,槲栎个体数量最多,共360株 (表2),占总数的22.8%,其次为马尾松 (280株),占总数的17.7%。麻栎个体数量也较多 (215株),占总数的13.6%。榆树个体数量最少 (31株),占总数的2.0%。平均胸径最大的是马尾松 (26.57 cm),其次是榆树 (12.33 cm),山胡椒的平均胸径最小 (3.36 cm)。
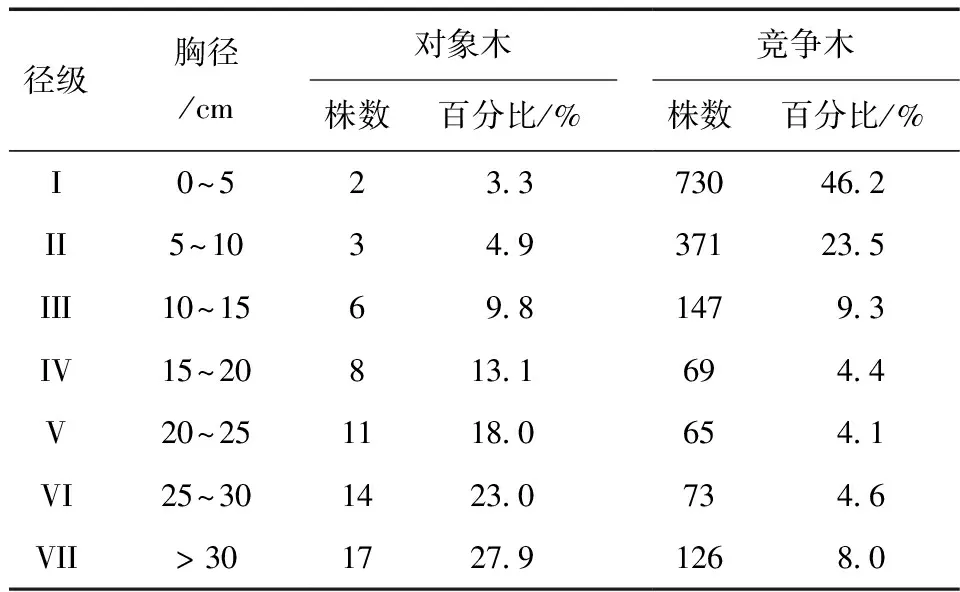
表1 马尾松和竞争木的胸径分布Table 1 The DBH distribution of P.massoniana and competitive trees
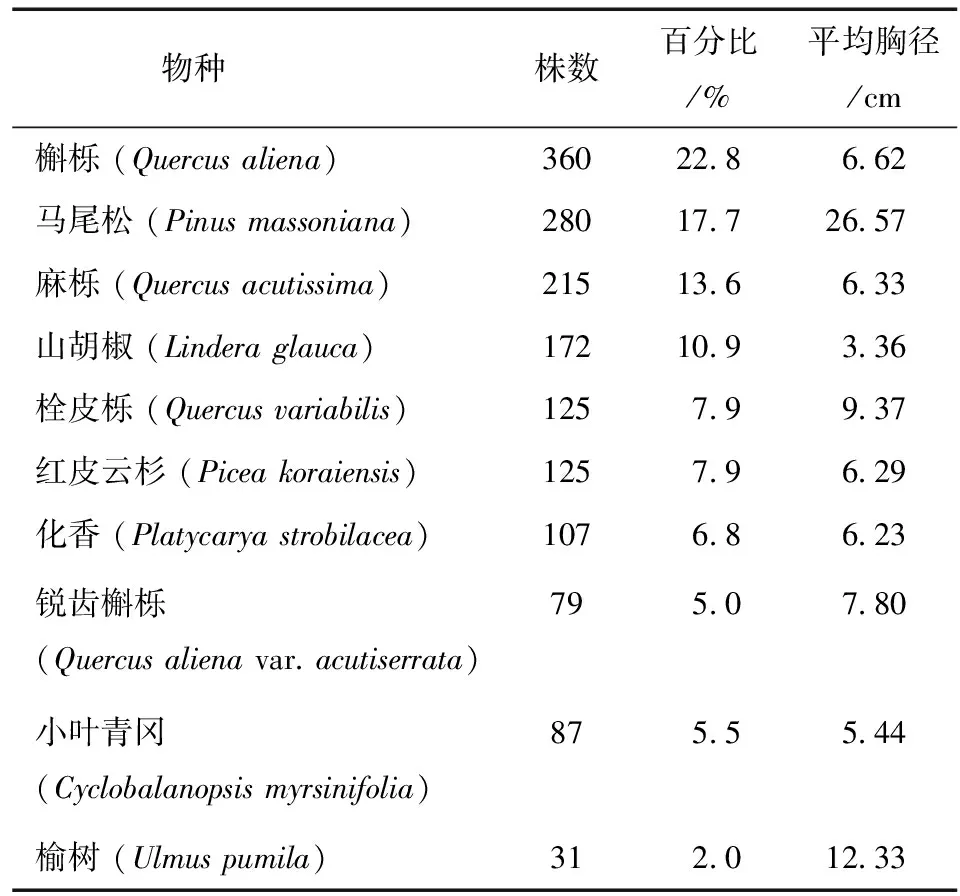
表2 竞争木的物种组成Table 2 The composition of competitive trees
3.2 马尾松种内与种间的竞争关系
样地内马尾松受到的种内竞争总强度为244.92 (表3),种间竞争总强度为176.10,种内竞争强度远大于种间竞争强度,说明宝天曼样地内马尾松的竞争主要来自于种内。由表3可知,马尾松受到的竞争强度大小与主要物种的径级结构以及物种数量密切相关,种内竞争中马尾松在Ⅰ径级 (0~5 cm) 和Ⅱ径级 (5~10 cm) 个体数量比较少,其竞争强度也比较小,分别为6.36和0.61。当胸径在10~25 cm范围时,对象木马尾松受到的种内竞争强度随着径级的增大而减小,在胸径 > 20 cm时,对象木马尾松受到的种内竞争强度又随着径级的增大而增大,特别是胸径 > 30 cm时,对象木马尾松受到的种内竞争强度最大 (160.52),占种内总竞争强度的65.5%,可能是因为该径级马尾松个体较大且数量最多 (119株) 所致。从各径级马尾松受到的种内竞争强度占种内总竞争强度的比例来看,对象木马尾松受到的种内竞争强度大致随着径级的增大而增大。从表3中还可以看出,对象木马尾松受到的种间竞争强度随着径级的增大而减小,在第Ⅰ径级,竞争木虽然个体较小,但数量最多 (719株),远大于马尾松个体数量,导致马尾松受到的种间竞争强度最大 (77.12),在胸径 > 30 cm时,竞争木个体数量较少,马尾松受到的种间竞争强度较小,仅为2.68。
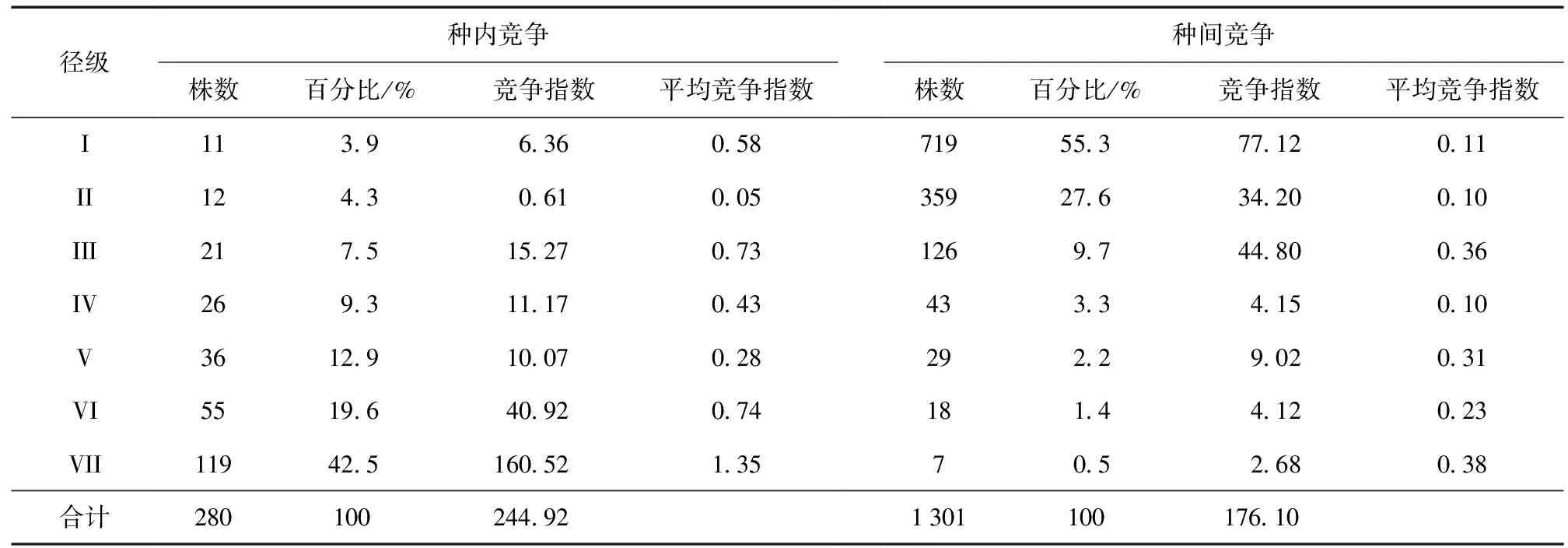
表3 马尾松种内与种间的竞争强度Table 3 Intraspecific and interspecific competition intensity of P.massoniana
样地中对象木马尾松受到的竞争强度大小不仅与竞争木大小以及数量有关,还与竞争木的种类有关,因竞争木种类不同而产生较大差异 (表4)。本研究中伴生种竞争木共9种,竞争总强度为176.10,其中竞争强度最大的是麻栎 (42.93),其次槲栎因个体数量较多且平均胸径较大,竞争强度排第2位 (40.22),山胡椒的个体数量较多,但平均胸径较小,竞争强度排在第8位 (5.23),榆树的平均胸径最大,但个体数量较少,其竞争强度最小 (1.45)。总的来看,样地内9种伴生种对马尾松产生的竞争强度大小顺序依次为:麻栎 > 槲栎 > 化香 > 栓皮栎 >红皮云杉 > 小叶青冈 > 锐齿槲栎 > 山胡椒 > 榆树。
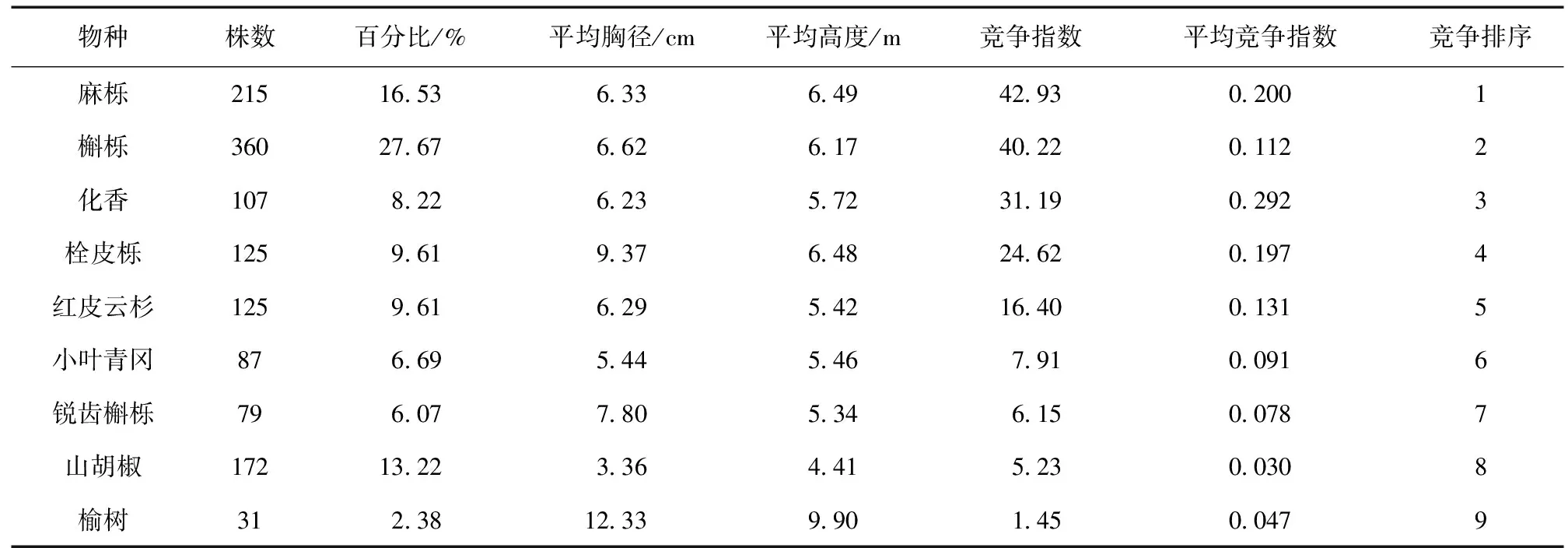
表4 伴生种竞争木的组成及其竞争强度排序Table 4 Composition of associated tree species and ranking of their competitive intensity
3.3 马尾松胸径与竞争指数的关系及其预测
以对象木马尾松胸径为自变量, 竞争指数为因变量,采用多种数学模型对马尾松胸径与竞争指数进行回归拟合,发现竞争指数与对象木胸径存在指数函数关系,即:
CI=AeBD
(2)
式中:CI为竞争强度;A和B为模型参数;D为对象木胸径;e为常数,显著性检验结果如表5所示,均达到极显著水平 (P< 0.01)。
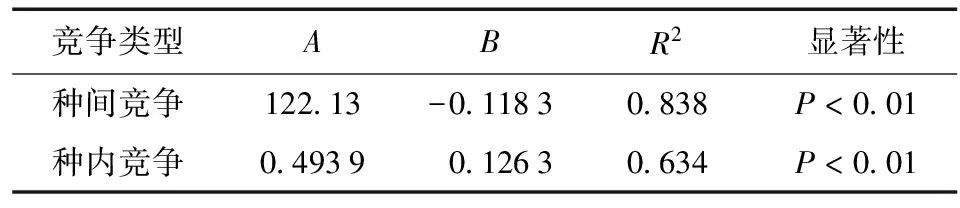
表5 竞争强度与马尾松胸径的模拟参数Table 5 The simulation parameters of competitive intensity and the DBH of P.massoniana
利用上述模型模拟和预测马尾松受到的种内和种间竞争强度 (表6),预测结果表明马尾松受到的种内竞争强度随着对象木胸径的增大而增大,种间竞争强度随着对象木胸径的增大而减小,预测结果与本文前面的分析基本一致,表明上述模型能很好的预测宝天曼马尾松-槲栎混交林中马尾松受到的竞争强度。
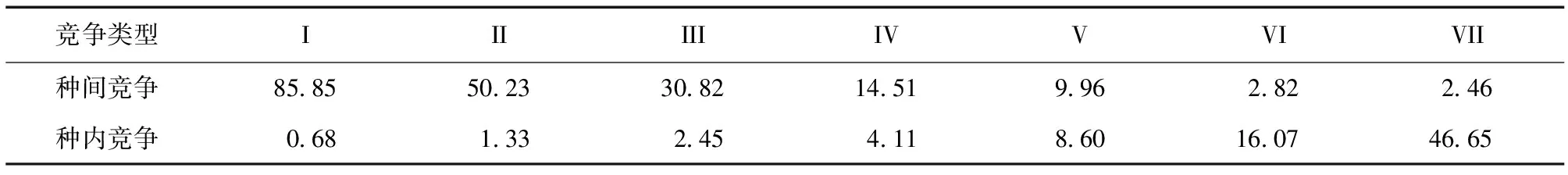
表6 马尾松胸径与竞争强度的模型预测Table 6 The prediction of the DBH and competitive intensity of P.massoniana
4 结论与讨论
宝天曼马尾松-槲栎混交林样地中,马尾松具有较高的相对优势度,其个体数量随着径级的增大逐渐增多。大径级马尾松较多可能是因为野獐马尾松人工林样地早期人工栽培使得马尾松成为绝对的优势物种。另外,第Ⅴ~Ⅶ径级分布的马尾松个体数量所占比例较大,呈 “右偏峰曲线” 分布,表明大径级马尾松较多,马尾松幼苗较少。在同种大树及伴生树种幼苗都较多的情况下,马尾松幼苗不能获得足够的光照与空间,生长受到抑制,不利于马尾松种群的更新。9种伴生树种个体数量随着径级的增大逐渐减少,主要分布于第I径级和第Ⅱ径级,表明伴生树种中大龄树较少,幼龄树较多,可能是因为优势物种马尾松成林后,形成主林层,对光照、水分等的竞争具有绝对优势,限制了伴生树种的生长。
样地内马尾松受到的竞争主要来自于种内,其受到的种内竞争总强度 (244.92) 大于种间竞争总强度 (176.10),即马尾松个体之间存在较大竞争,尤其是当胸径 > 30 cm时,马尾松受到的种内竞争指数 (160.52) 远远大于种间竞争指数 (2.68),这与马尾松大龄树较多,同径级的其他竞争木数量又偏少的现象一致。研究结果与胡刚等[23]对青冈栎 (Cyclobalanopsisglauca) 的种内种间竞争关系研究结果类似,都是种内竞争大于种间竞争。径级较大的马尾松个体数量较多,且生态习性相同,因而种内竞争激烈[7],又由于马尾松在主林层占据优势地位,来自于其他物种的竞争压力较小,因此马尾松受到的种间竞争强度较小。付梦瑶等[10]也对松栎混交林中马尾松的种内种间竞争进行了研究,但其调查样地内对象木马尾松多为幼龄林和中龄林,个体较小且处在群落的下层,对环境资源的争夺不及其他较大个体竞争木,因此马尾松受到的种间竞争强度大于种内竞争强度,与本文结论有所不同。
对象木马尾松受到的种间竞争强度因竞争木的种类不同而产生较大差异,生态习性越接近的植物物种,对资源和空间的利用具有相似性,种间的竞争更激烈[5]。伴生树种中竞争强度最大的是麻栎 (42.93),其次是槲栎 (40.22),可能是因为麻栎和槲栎与马尾松的生态习性相近以及生态位重叠幅度较大,所以彼此之间竞争程度剧烈[24]。
本研究还发现样地内对象木马尾松的胸径与竞争强度之间存在指数函数关系,表现为马尾松受到的种内竞争强度随着径级的增大而增大,种间竞争强度随着径级的增大而减小。黄小波等[7]和项小燕等[25]分别对云南松 (Pinusyunnanensis) 和大别山五针松 (Pinusdabeshanensis) 的种内种间竞争关系进行研究,结果表明随着对象木胸径的增大,其受到的种内和种间竞争强度都逐渐减弱,与本文结论有所差别,可能与样地内竞争木和对象木的数量以及个体大小、生长状况存在较大差异有关。随着径级的增大,对象木和竞争木的个体数量都逐渐减少,当同径级的竞争木数量多于对象木数量的话,对象木受到的竞争压力就比较大。本试验马尾松-槲栎混交林中马尾松是造林树种,在群落中占据主导地位,其竞争能力强于其他伴生树种,同一批播种的马尾松个体大小相当,对环境资源的利用较为均匀,随着林分的发育,逐渐进入林冠层,成为主林层的优势物种,同径级的其他竞争木数量较少,因而受到的种间竞争强度较小,但同径级的马尾松数量就较多,因而受到的种内竞争强度较大,且胸径越大,种内竞争强度越大。
因此,宝天曼马尾松-槲栎混交林中马尾松受到的种内竞争强度高于种间竞争强度,马尾松幼苗较少,种群更新困难,而栎类群落增长潜力较大。可以通过适当的砍伐大径级马尾松,以减少其种内竞争压力。采伐部分伴生树种,尤其是槲栎和麻栎,减少其对马尾松幼苗产生的竞争压力,最大限度地为幼苗提供生长空间,同时人工抚育马尾松幼苗来实现其种群更新。研究结果可为宝天曼国家级自然保护区森林的保护与管理提供一定的借鉴。在全球变化背景下,探讨马尾松在其分布的北部边界因竞争引起的种群动态和分布格局还有待进一步研究。
[参考文献]
[1] 杜峰, 梁宗锁, 胡莉娟. 植物竞争研究综述[J]. 生态学杂志, 2004, 23(4): 157-163.
[2] Sabatia O, Burkhart H E. Competition among loblolly pine trees: does genetic variability of the trees in a stand matter? [J]. Forest Ecology and Management, 2012, 263(1): 122-130.
[3] 徐道炜, 刘金福, 洪伟, 等. 森林群落种内种间竞争研究进展[J]. 亚热带农业研究, 2014, 10(3): 199-200.
[4] Bella I E. A new competition model for individual trees[J]. Forest Science, 1971, 17(3): 364-372.
[5] 张池, 黄忠良, 李炯, 等. 黄果厚壳桂种内与种间竞争的数量关系[J]. 应用生态学报, 2006, 17(1): 22-26.
[6] 张莉, 陆畅, 李晓红, 等. 皇藏峪国家自然保护区青檀种群年龄结构及种内、种间竞争[J]. 上海交通大学学报 (农业科学版), 2012, 30(1): 34-40.
[7] 黄小波, 刘万德, 苏建荣, 等. 云南松天然林的种内和种间竞争[J]. 林业科学研究, 2016, 29(2): 209-215.
[8] 张忠华, 胡刚, 秦川, 等. 青秀山风景区优势种大叶栎种内与种间的竞争关系[J]. 中南林业科技大学学报, 2016, 36(1): 67-71, 85.
[9] 吴昊. 秦岭山地松栎混交林土壤养分空间变异及其与地形因子的关系[J]. 自然资源学报, 2015, 30(5): 858-868.
[10]付梦瑶, 崔秋芳, 吴明, 等. 松栎混交林中马尾松的种内种间竞争[J]. 河南农业大学学报, 2016, 50(3): 311-317.
[11] 刘玉萃, 吴明作, 郭宗民, 等. 宝天曼自然保护区栓皮栎林生物量和净生产力研究[J]. 应用生态学报, 1998, 9(6): 569-574.
[12] 袁志良, 王婷, 朱学灵, 等. 宝天曼落叶阔叶林样地栓皮栎种群空间格局[J]. 生物多样性, 2011, 19(2): 224-231.
[13] 闫东锋, 朱滢, 杨喜田. 宝天曼栎类天然林物种多样性与稳定性[J]. 浙江农林大学学报, 2011, 28(4): 628-633.
[14] 殷卫抗, 袁志良, 王亚平, 等. 宝天曼锐齿栎林土壤空间异质性的相关分析[J]. 河南农业大学学报, 2013, 47(3): 278-283.
[15] 臧婧. 河南宝天曼落叶阔叶林种-面积关系研究[D]. 哈尔滨: 东北林业大学, 2015.
[16] Hegyi F. A simulation model for managing jack-pine stands[M]// Growth Models for Tree and Stand Simulation. Stockholm: Royal College of Forestry, 1974: 74-90.
[17] 郭建荣, 庄静静, 朱学灵, 等. 不同林龄对宝天曼自然保护区锐齿栎林土壤物理性质及其水源涵养功能差异性的影响[J]. 河南农业大学学报, 2012, 46(5): 549-552, 576.
[18] 闫满玉, 杜晓军, 赵爱花, 等. 河南宝天曼落叶阔叶林木本植物单物种-面积关系[J]. 生物多样性, 2015, 23(5): 630-640.
[19] 王婷, 李聪, 张弘, 等. 宝天曼自然保护区不同针叶树径向生长对气候的响应[J]. 生态学报, 2016, 36(17): 5324-5332.
[20] 刘方炎, 李昆, 廖声熙, 等. 濒危植物翠柏的个体生长动态及种群结构与种内竞争[J]. 林业科学, 2010, 46(10): 23-28.
[21] 曾思齐, 龙时胜, 肖化顺, 等. 南方地区青冈栎次生林种内与种间竞争研究[J]. 中南林业科技大学学报, 2016, 36(10): 1-5.
[22] 刘艳会, 刘金福, 何中声, 等. 基于戴云山固定样地黄山松群落物种组成与结构研究[J]. 广西植物, 2017, 37(7): 881-890.
[23] 胡刚, 梁士楚, 张忠华, 等. 桂林岩溶石山青冈栎种内与种间竞争的数量关系[J]. 西北林学院学报, 2008, 22(5): 32-36.
[24] 张谧, 韩烁, 李钧涛, 等. 雾灵山自然保护区油松、白桦及山杨天然林竞争关系研究[J]. 北京师范大学学报 (自然科学版), 2007, 43(2): 184-186.
[25] 项小燕, 吴甘霖, 段仁燕, 等. 大别山五针松种内和种间竞争强度[J]. 生态学报, 2015, 35(2): 389-392.
