接种真菌对马尾松幼苗抗铜性的影响
2018-04-24鲁彦君
高 丙,黄 建,鲁彦君
(西北农林科技大学 林学院,陕西 杨陵 712100)
铜矿区的生态修复及重金属污染治理已经成为一个日益突出的问题[1]。近年来研究表明,外生菌根真菌在矿区废弃地植物修复与造林工程中潜力巨大。外生菌根及菌丝体既能改良土壤结构[2],又能促进水分与营养元素的吸收[3],还能发挥“屏障”作用阻止重金属离子向宿主植物地上部分转运[4],促进植物在重金属污染的土壤中生长[5]。近年来的研究显示,外生菌根除了直接调节宿主植物根系对水分与营养元素的吸收,还可以改善宿主植物响应重金属胁迫的光合系统、抗氧化系统以及植物激素等一系列生理响应过程,提高宿主植物的解毒作用[6]。
因此,采用从铜矿区马尾松菌根群落调查中分离的彩色豆马勃中的一个抗铜性强的固体纯培养菌株(Ptch)侵染马尾松,研究菌根化马尾松幼苗在铜胁迫条件下的生长、光合特性与抗氧化酶活性的变化规律,探讨具有重金属耐性的外生菌根真菌对宿主植物抗铜性的影响,为筛选与合理利用外生菌根菌修复铜矿区废弃地提供试验依据。
1 材料与方法
1.1 外生菌根真菌培养与菌剂制备
外生菌根真菌彩色豆马勃用于侵染的菌株(Ptch)由本实验室前期分离鉴定获得,采用MMN培养基进行固体培养,培养时使用直径为6 mm的打孔器将其切割转移至含有MMN固体培养基的培养皿(直径9 cm)中,温度25℃,相对湿度60%,暗培养4周,待菌丝布满培养基后用于马尾松接种。
1.2 马尾松培养与接种
试验树种马尾松(Pinusmassoniana)的种子由陕西省林业厅林业技术推广总站提供。种子发芽处理采用马琼[7]等(2005)的方法,2015年12月,选取籽粒饱满种子,流水冲洗,消毒,温水浸种,置于25℃恒温箱催芽,待30%~40%种子裂嘴露白时播种。种子播入孔穴盘(灭菌沙子)育苗,4周后挑选长势一致的幼苗移栽。幼苗移栽在容量为1 L的花盆中,基质为经过高温(121℃,灭菌2 h)灭菌的河沙与土(体积比为2∶1),灭菌放置7 d后使用,共栽种250盆。
每个花盆移栽2株,同时进行接种,接种的方法参考刘藩[8](2015),将培养基内的固体培养基接种于花盆中,每盆接种4个菌饼,放在根系周围,每月新接种一个菌饼,加入MMN营养液,盆栽幼苗放置在西北农林科技大学温室内(25℃,光周期14 h光照/d,湿度70%)培养。6个月后,随机选择接菌与对照幼苗5株,用自来水冲洗根后,用显微镜观察菌根的形成情况。
1.3 铜胁迫处理
将菌根化苗(M)与非菌根化苗(N)各70盆,各分为2组:一组对照,一组铜胁迫处理;在1/2 Hoagland营养液中加入CuSO4·5H2O(90 μmol·L-1)进行40 d处理,以每周30 μmol·L-1×1 L加入,累计3次终浓度达到90 μmol·L-1,非菌根化苗铜胁迫处理记为NCu,菌根化苗铜胁迫处理记为MCu,以1/2 Hoagland 营养液(含0.3 μmol·L-1CuSO4·5H2O)作为对照,非菌根化苗对照记为NC,菌根化苗对照记为MC。
1.4 菌根化苗的生理试验
1.4.1 生长指标测定 在铜胁迫处理结束时,随机选取不同处理下的菌根化苗与非菌根化苗各5盆,用电子天平(精度0.000 1 g)称量针叶、茎、根的鲜质量(Fw),后放入105℃烘箱杀青15 ~ 20 min,然后80℃烘干至恒重,测定干质量(Dw),并计算组织相对含水量(占鲜重%)=(Fw-Dw)/Fw×100%。
1.4.2 光合气体交换参数与叶绿素荧光参数测定 应用Li-6400(标准叶室2×3=6 cm2)便携式光合测量系统(Li-Cor Inc.,Lincoln,Nebraska,USA)测定针叶的净光合速率(Pn)、气孔导度(Cond)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。参数设置参照高琼[9]等(2014)的方法,测定时间为晴天9:00-11:00,选取每株幼苗中部生长良好的的8片针叶作为测试叶,平铺放置充满叶室面积,待数据稳定后取3个数据,取其平均值。
应用脉冲调制式荧光系统(FMS-2型,英国Hansatech公司),测定不同程度铜胁迫下幼苗针叶的叶绿素荧光动力学参数。参数设置与计算方法参照王琰[10]等(2011)的方法,选取每株幼苗中部生长良好的针叶8片,平铺放置于叶室测定。试验条件相对稳定,气温(25±0.5)℃,空气相对湿度(45±1)%。
1.4.3 抗氧化酶活性与丙二醛含量测定 采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性;采用愈创木酚显色法测定过氧化物酶(POD)活性;采用紫外吸收法测定过氧化氢酶(CAT)活性;丙二醛(MDA)含量的测定采用双组分光光度法[11]。
1.4.4 数据分析 数据采用SPSS(13.0)统计软件处理,用单因素方差分析(oneway-ANOVA)检验不同浓度铜胁迫处理间的样本数据结果的差异显著性,采用Duncan(P<0.05)法进行显著性多重比较,采用origin 9软件作图。
2 结果与分析
2.1 生长适应性
用Ptch接种马尾松,共生6个月后形成典型的外生菌根结构,通过随机抽检,发现侵染率约为60%。对照条件下,菌根化苗与非菌根化苗的茎、叶干重无明显区别,但根干重比非菌根化苗高43.9%,根、茎、叶的相对含水量(RWC)比非菌根化苗高6.0%~14.0%(表1)。铜胁迫后,2种苗的干重与相对含水量均呈下降趋势,非菌根化苗根、茎、叶干重与相对含水量分别下降了56.1%~75.8%和20.8%~44.7%,而菌根化苗下降了16.0%~48.4%与9.1%~27.6%。可见,铜胁迫明显抑制非菌根化苗的生长,接种真菌增强了宿主植物的生长适应性。

表1 铜胁迫对菌根化苗与非菌根化苗根、茎、叶干重与相对含水量的影响
注:每一个值(±SE)是5个样本的重复,小写字母表示不同处理间差异显著(P<0.05)。下图同。
2.2 光合气体交换
菌根化苗与非菌根化苗显示了不同的光合作用能力,对照条件下,菌根化苗净光合速率(Pn)明显高于非菌根化苗(高19.9%),但气孔导度(Cond)与蒸腾速率(Tr)比非菌根化苗低(32.9%与17.7%),铜胁迫诱导2种苗的Pn、Cond与Tr有不同程度下降,但非菌根化苗下降幅度明显大于菌根化苗,菌根化苗的Pn、Cond与Tr分别下降25.4%,56.0%与50.6%,非菌根化苗为38.9%,66.7%与70.8%(图1A,B,C)。值得注意的是,铜胁迫诱导菌根化苗胞间CO2浓度(Ci)显著下降38.4%(P<0.05),而非菌根化苗并无明显变化(图1D)。
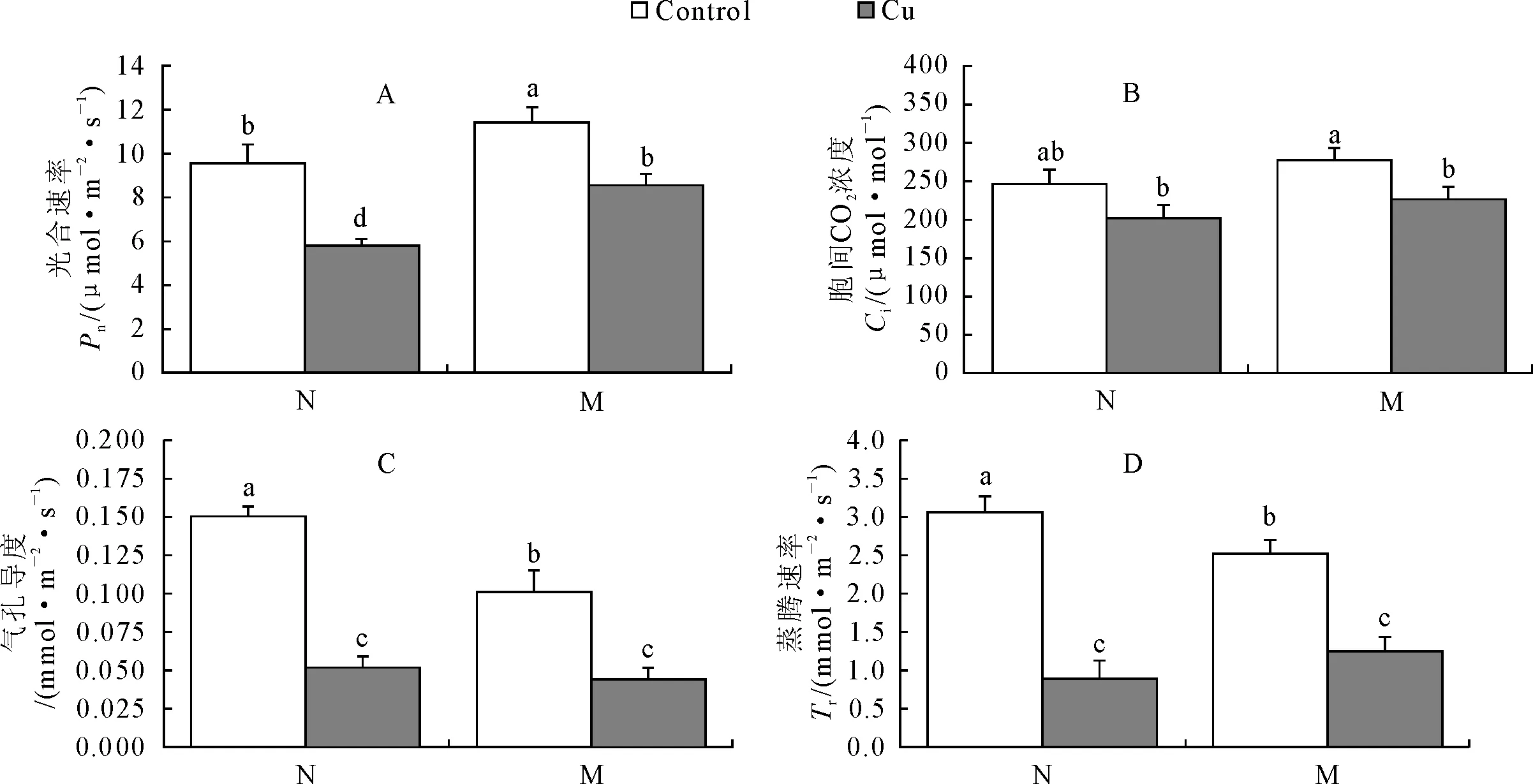
图1 菌根化苗与非菌根化苗在铜胁迫下的光合速率,胞间CO2浓度、气孔导度、蒸腾速率的变化
2.3 叶绿素荧光参数
叶绿素荧光产量为我们进一步获得光化学与非光化学过程中的光合性能提供了有用的信息。铜胁迫下,非菌根化苗初始荧光(Fo)比对照有显著升高(升高了31.0%),最大荧光(Fm)、最大光化学效率(Fv/Fm)、光化学淬灭(qP)、实际光化学效率(ΦPSⅡ)以及光化学淬灭(NPQ)均较对照明显下降,下降幅度在15.5%~39.7%,并且ΦPSⅡ的下降幅度大于Fv/Fm(图2),表明非菌根化苗的光抑制已经产生。与非菌根化苗不同的是,菌根化苗在铜胁迫下Fo、Fv/Fm无明显变化,Fm、qP与ΦPSⅡ分别下降7.3%、30.5%与20.6%,NPQ却增加了17.1%(图2),表明铜胁迫可增加菌根化苗针叶PSⅡ天线色素的热耗散,在一定程度上缓解胁迫对光合作用的影响。
2.4 抗氧化酶活性与丙二醛
菌根化与非菌根化苗在铜胁迫下的超氧化物歧化酶(SOD)、过氧化物酶(POD)与过氧化氢酶(CAT)以及丙二醛(MDA)含量显示了不同的变化规律,对照条件下,菌根化苗与非菌根化苗的SOD、POD、CAT活性以及MDA含量均无明显差异(P<0.05),铜胁迫后,2种苗的SOD、POD以及CAT的活性均表现出升高趋势,增加幅度分别是27.5%~176.5%与54.6%~108.9%(图3),表明铜胁迫诱导SOD、POD与CAT活性升高,保护光合作用机构免受损伤。铜胁迫后,非菌根化苗MDA含量升高为对照的31.5%,菌根化苗MDA含量并无显著变化,表明铜胁迫下,非菌根化苗针叶活性氧产生大于细胞活性氧清除能力。
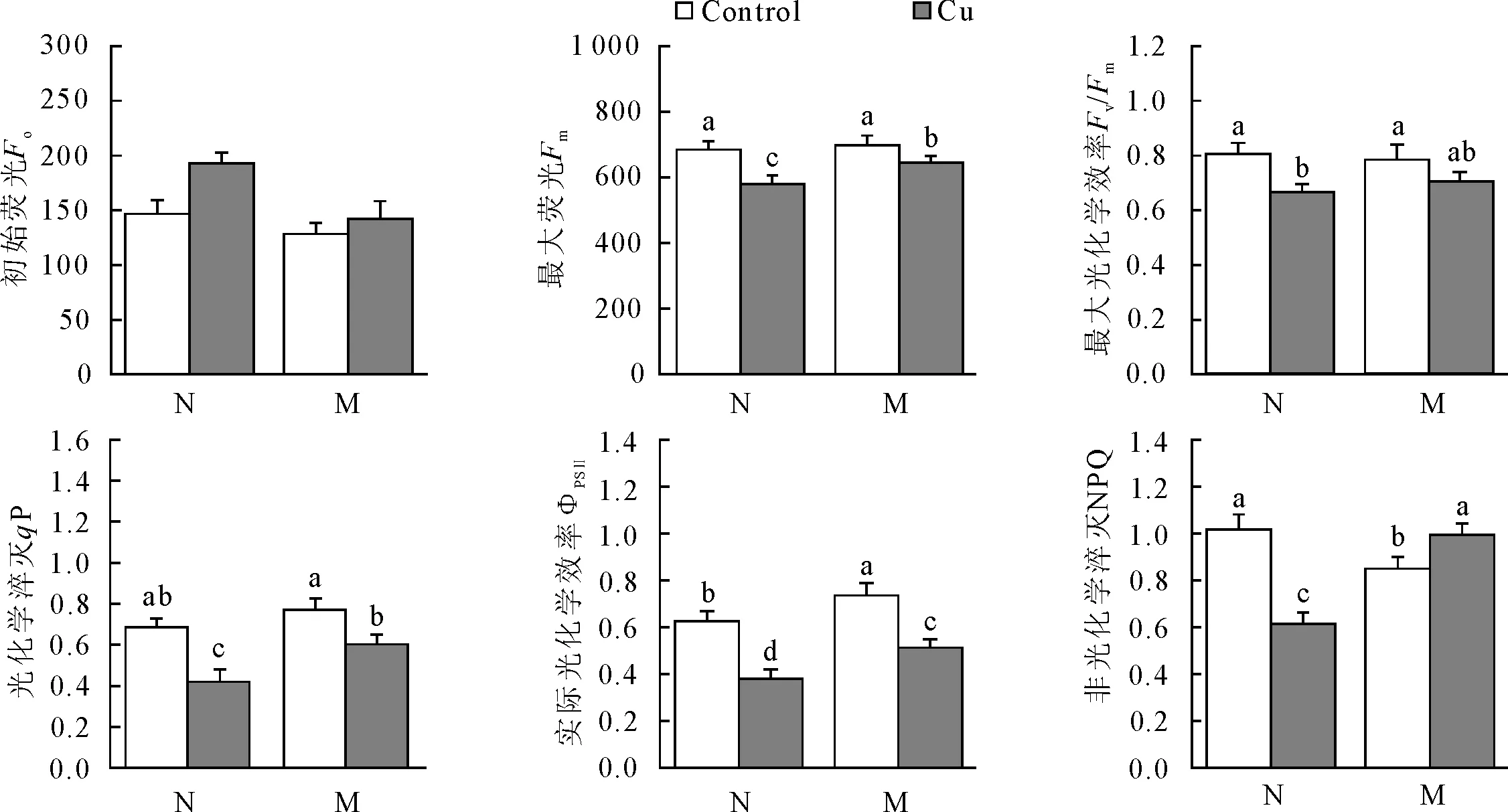
图2 Cu胁迫下菌根化苗与非菌根化苗叶绿素荧光参数的变化
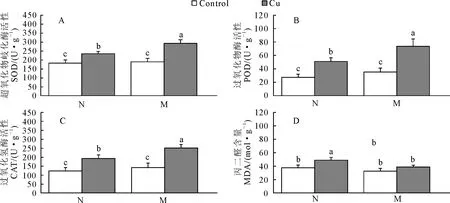
图3 铜胁迫下菌根化苗与非菌根化苗的超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)的活性与丙二醛(MDA)含量的变化
3 结论与讨论
对接种了Ptch的马尾松苗与未接种的马尾松苗在铜胁迫处理条件下的生长与光合生理特性进行了比较,初步探索菌根在马尾松苗抗铜胁迫中的作用。一般植物在胁迫条件下表现出明显的生长(茎鲜重/干重,植物高度,叶面积以及叶数量等)抑制现象[12],但本研究中菌根化苗相对非菌根化苗依然维持了较高的根系生长量(表1),对于植物在胁迫条件下吸收水分与营养元素至关重要[13]。维持较高的相对含水量(RWC)有助于植物正常生理生化反应过程的进行[13]。菌根化苗各器官在对照条件下保持了较高RWC,铜胁迫虽诱导其下降,但下降程度明显小于非菌根化苗,说明接种真菌有助于宿主植物水分吸收,从而维持正常生理过程。
植物生物量积累与光合作用密切相关[14]。铜胁迫诱导菌根化苗与非菌根化苗的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)随之下降,并且菌根化苗下降程度低于非菌根化苗,说明菌根结构可以在铜胁迫条件下提高马尾松苗的光合作用能力,有利于其生物量的积累。通常影响植物光合作用的因素可分为气孔因素和非气孔因素,前者是水分胁迫导致气孔导度减小,阻碍CO2进入叶片使光合下降;后者由于叶肉细胞的光合活性下降使净光合速率下降[15]。结合非菌根化苗在铜胁迫下的Gs下降幅度远大于菌根化苗,Ci却没有明显降低,推测其Pn降低的原因由非气孔因素造成,铜胁迫会导致叶肉光合活性下降,而菌根结构对叶肉光合活性具有一定保护作用。
叶绿素荧光动力学手段,能快速灵敏、无损伤地反映“内在的”光系统Ⅱ(PSⅡ)对光能的吸收、传递、耗散、分配等方面的状况,是进一步证明植物光合能力以及对环境胁迫响应的有效手段[16]。非菌根化苗的Fo显著升高,Fm、Fv/Fm、ΦPSⅡ以及NPQ都显著下降(图 2),表明光系统受到一定程度损伤,热耗散受抑制,光抑制增加,由此推测,非菌根化苗光系统PSⅡ的损伤是Pn下降的非气孔限制的可能原因,因为一定程度的胁迫可以使松树针叶的PSⅡ活性中心受损[17];而菌根化苗的PSⅡ实际光化学效率(ΦPSⅡ)与光化学猝灭(qP)均显著降低,但程度小于非菌根化苗,最大荧光(Fm)、最大光化学效率(Fv/Fm)与初始荧光(Fo)并未发生显著变化,但非光化学猝灭(NPQ)却显著升高,表明菌根化苗在铜胁迫下启动了光响应防御机制,天线色素捕获的光能向PSⅡ反应中心传递的效率下降,热耗散能力有一定增加,减少了反应中心过剩光能的积累,保护了光合机构免受破坏[16]。
铜胁迫很容易引起植物氧化损伤,产生过量活性氧,作用于许多细胞组分,造成细胞组分破坏[18]。铜胁迫诱导菌根化与非菌根化苗的抗氧化酶活性(SOD、POD与CAT)升高,说明马尾松苗针叶PSⅡ的热耗散和酶活性协调发挥作用,维持光合结构的正常功能。但菌根化苗抗氧化酶活性增幅大于非菌根化苗,MDA增幅则远小于非菌根化苗,表明Ptch可提高马尾松苗在铜胁迫下的抗氧化酶活性,减少光合系统损伤,降低光抑制程度,保持较为稳定的净光合速率,增加菌根化苗生物量的积累,有助于提高马尾松苗的抗铜性。
参考文献:
[1] SHEN K,HUANG J,NARA K,etal.Inoculation of ectomycorrhizal fungi contributes to the survival of tree seedlings in a copper mine tailing[J].Journal of Forest Research,2017,20(6):493-500.
[2] RILLIG M C,MUMMEY D L.Mycorrhizas and soil structure[J].New Phytologist,2006,171(1):41-53.
[3] DRUEBERT C,LANG C,VALTANEN K,etal.Beech carbon productivity as driver of ectomycorrhizal abundance and diversity[J].Plant Cell & Environment,2009,32(8):992-1003.
[4] BELLION M,COURBOT M,JACOB C,etal.Extracellular and cellular mechanisms sustaining metal tolerance in ectomycorrhizal fungi[J].Fems Microbiology Letters,2006,254(2):173-181.
[5] 黄艺,黄志基.外生菌根与植物抗重金属胁迫机理[J].生态学杂志,2005,24(4):422-427.
[6] MA Y L,HE J L,MA C F,etal.Ectomycorrhizas withPaxillusinvolutusenhance cadmium uptake and tolerance inPopulus×canescens[J].Plant Cell and Environment,2014,3(37):627-642.
[7] 马琼,黄建国,蒋剑波.接种外生菌根真菌对马尾松幼苗生长的影响[J].福建林业科技.2005,32(2):85-88.
[8] 刘藩,杨晨,刘同先,等.杨树接种外生菌根真菌卷缘桩菇对杨扇舟蛾生长发育的影响[J].西北林学院学报.2015,30(3):145-148.
[9] 高琼,王维有,梁冬,等.8个种源油松生长和光合生理指标的比较研究[J].北京林业大学学报,2014,36(2):87-93.
[10] 王琰,陈建文,狄晓艳.不同油松种源光合和荧光参数对水分胁迫的响应特征[J].生态学报,2011,31(23):7031-7038.
[11] 李明军,刘萍.植物生理学实验技术(21世纪生物学基础课系列实验教材)[M].北京:科学出版社,2007.
[13] BOJARCZUK K,KARLINSKI L,HAZUBSKA-PRZYBYT,etal.Influence of mycorrhizal inoculation on growth of micropropagatedPopulus×canescenslines in metal-contaminated soils[J].New Forests,2015,46(2):195-215.
[14] 龚佳,倪细炉,李健.NaHCO3胁迫对宁夏4种灌木生长及光合特性的影响[J].西北林学院学报,2017,32(2):8-15.
GONG J,NI X L,LI J.Effects of NaHCO3etress on the growth and photosynthetic charactreristics of four shrubs in Ningxia[J].Journal of Northwest Forestry University,2017,32(2):8-15.(in Chinese)
[15] 马富举,李丹丹,蔡剑,等.干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响[J].应用生态学报.2012,23(3):724-730.
MA F J,LI D D,CAI J,etal.Responses of wheat seedlings root growth and leaf photosynthesis to drought stress[J].Chinese Journal of Applied Ecology,2012,23(3):724-730.(in Chinese)
[16] 裴斌,张光灿,张淑勇,等.土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响[J].生态学报,2013,33(5):1386-1396.
[17] 刘亚丽,王庆成,杨远彪.水分胁迫对脂松幼苗叶绿素荧光特征的影响[J].植物研究,2011,31(2):175-179.
[18] 冯欢,豆青,王海华,等.2种外生菌根真菌的铅耐受性及相关机制[J].西北林学院学报,2017,32(2):188-196.
FENG H,DOU Q,WANG H H,etal.Lead tolerance of two ectomycorrhizal fungi and related mechanisms[J].Journal of Northwest Forestry University,2017,32(2):188-196.(in Chinese)
