过敏性哮喘的研究进展*
2018-04-20余芳
余 芳
(贵州医科大学附属医院临床检验中心,临床医学研究中心,贵州医科大学免疫转化医学重点实验室,贵阳 550004)
近数十年来,过敏病例在世界很多地区呈快速上升趋势。常见过敏性疾病包括过敏性鼻炎、哮喘、结膜炎、湿疹、食物过敏、药物过敏和全身性严重过敏反应等。世界卫生组织已把过敏性疾病列为21世纪需重点研究和防治的三大疾病之一。流行病学调查显示世界人口的5%~16%都罹患有过敏性哮喘,特别是儿童患有过敏性哮喘的比例持续增加[1,2]。我们临床工作发现,有大于15%的以久咳或难治性支气管炎、肺炎等就诊的患儿,经血清总IgE及特异性过敏原检测发现,实为过敏原诱发的呼吸道病症。过敏性哮喘不仅影响发病儿童生活质量,而且影响其正常的生长发育。作为一个复杂疾病,哮喘的发病机制与多达100种以上的遗传位点和复杂环境因素的相互作用相关联[3], 而对于过敏性疾病迄今为止还没有令人满意的治疗方法。进一步认识发病机制,寻找对过敏疾病有重要调控作用的关键分子或相关基因,采用多靶点联合治疗,具有极大的实用价值,这也一直是该领域研究者的关注点。目前研究集中在几大方向:过敏免疫微环境变化(其中天然免疫系统与特异性免疫系统相互作用尤为突出),表观基因组学与过敏性哮喘,微生态与疾病,以及过敏原特异性免疫治疗等。现就过敏性哮喘的相关研究现状综述如下。
1 过敏性哮喘免疫微环境
气道上皮细胞(Epithelial cells,ECs)是呼吸道的第一道防线[4],表达多种模式识别受体,如TLR4,识别病原相关分子模式(Pathogen-associated molecular patterns,PAMPs)或损伤相关分子模式(Damage-associated molecular patterns,DAMPs),活化产生多种细胞因子,包括胸腺基质淋巴细胞生成素(Thymic stromal lymphopoietin,TSLP)、GM-CSF、IL-1 α、IL-1β、IL-25以及IL-33等,进一步激活天然免疫细胞如诱导树突状细胞(Dendritic cell,DC)向促进Ⅱ型免疫反应方向分化[5],或作用于适应性免疫细胞如Th2细胞分泌Ⅱ型细胞因子,如IL-4、IL-5、IL-13等,进而诱导嗜酸性粒细胞、肥大细胞等聚集,以及反过来影响ECs细胞连接及皮下黏液层修复等[4,6]。ECs还产生生长因子如VEGF-A、VEGF-C等,诱导微环境中生成新的血管和淋巴管,有利于免疫细胞趋化及募集,如DC向淋巴结趋化[7]。
由于机体局部及系统大多可检测到Ⅱ型炎性因子,如IL-4、IL-5、IL-13等,以及血清中大量升高的IgE,认为其是Th2细胞为中心的病理过程,故而被纳入Ⅱ型炎性反应疾病类型。由此,围绕Th2细胞来开展的机制研究,包括其在由天然免疫系统递呈的过敏原刺激下产生的炎性介质,介质对下游其他效应细胞,如嗜酸性粒细胞、B细胞、肥大细胞、平滑肌细胞等的作用。动物哮喘疾病模型对这个机制上有很细节的解释,而目前也有很多针对Ⅱ型炎性反应通路中重要介质的阻断治疗,尽管一些早期临床实验数据表明也具有一定的疗效,但遗憾的是除了IL-5拮抗剂外,其他都没有得到批准用于哮喘的临床治疗。过敏性炎症因子(如5-HT、U46619、组胺、LTD4等)与神经递质存在有显著的协同效应,其中TMEM16A氯通道是该协同效应的关键分子,抑制该氯通道可以极大地抑制呼吸道高反应性、降低呼吸道阻力、明显改善哮喘症状[8]。
2 天然淋巴样细胞参与过敏性疾病
尽管Ⅱ型免疫反应中Th2细胞的中心作用已有很好的证据,但对Th2反应的始动源于何处,仍不很明确。近来研究发现,Ⅱ型天然淋巴样细胞(Innate lymphoid cells,ILCs)、ILC2[或Nuocyte,Nature helper cells(NHC)、Innate helper cells 2(Ih2)],可能是Ⅱ型细胞因子如IL-5、IL-13的主要来源[9,10],在肺部炎症中架设天然免疫和获得性免疫的桥梁[11,12]。ILC2可被由ECs受到外源信号刺激产生的如IL-33、IL-25、TSLP及IL-7等所活化,并产生多种效应介质,与其他效应细胞相互作用,影响疾病过程。ILC2活化可能是由细胞内蛋白激酶C-θ(PKC-θ)介导[13],蛋白激酶C-θ(PKC-θ)缺失小鼠的ILC2细胞数量减少,肺组织IRF4和NFAT1表达水平降低,进而影响下游Th2细胞的活化。IL-33单独即可活化ILC2[14],而IL-25则可能是通过刺激基质产生IL-33和TSLP而对ILC2起作用[15]。
ILC2可能是应对过敏原最早产生IL-13的细胞[16,17],诱导替代性活化的巨噬细胞(Alternatively activated macrophages,AAMs)活化、树突状细胞迁移,IL-5可募集活化嗜酸性粒细胞,而与B细胞,则可能是通过活化Th2细胞产生的IL-4以及ICOS-L,活化B细胞分泌更多IgE。ILC2可能经诱导可表达MHCII,充当抗原递呈细胞,与T细胞相互作用,还能分泌 IL-4及OX40L,诱导Th2细胞产生更多IL-4、IL-5、IL-13、IL-9等,而Th2细胞产生的IL-9和IL-2等也可以促进ILC2细胞活性。ILC2也是气道炎性反应中IL-9的主要来源[18],虽然其作用还不明确,但IL-9是肥大细胞的生长因子,提示ILC2与肥大细胞间的联系。ILC2表达半胱氨酰白细胞三烯1受体(Cysteinyl leukotrienes 1 receptor,CysLT1R),介导白细胞三烯C4(LTC4)的效应,与 IL-33联合作用可诱导更严重的Ⅱ型免疫反应[19]。而肥大细胞是LTC的重要来源之一,提示肥大细胞与ILC2在过敏性哮喘中的交互作用。
有研究显示,过敏原诱导的气道反应,如果暴露于环境污染物,如汽车尾气物质(Diesel exhaust particles,DEPs)等,炎性反应程度将进一步增加。而ILC2及Th2细胞的功能障碍可能是其原因之一。但动物模型研究发现,Rag2缺失时相关疾病完全被阻断,而在ILC2缺失时,疾病仅可见轻微缓解。可见T细胞仍是疾病发生发展不可或缺的中心环节[20]。最近研究揭示,ILC2与神经元存在相互调控作用[21-23]。ILC2选择性表达神经介质U(Neuromedin U,NMU)受体NMUR1,肺组织中可看到神经纤维结构,ILC2的位置相毗邻,提示二者可能有相互作用的组织结构支撑。NMUR1信号阻断可下调ILC2活化所致的炎性改变,揭示了肺部局部效应组织与神经系统的相互关系在过敏性哮喘中的作用[21],在消化道小肠黏膜也有类似发现[22]。这些研究提示,黏膜神经系统与区域免疫系统共同完成对局部组织功能及稳态的调节。
3 表观基因组学与过敏性哮喘
表观遗传学修饰,如DNA甲基化、基因组印记和RNA编辑等,通过对DNA组装入核的方式进行修饰,改变基因被转录元件接近的容易程度,进而影响基因组稳定性。而这些如甲基化(Methylation)和乙酰化(Acetylation)等化学修饰的过程,较易受到环境等外界因素的影响,活化原本沉默的基因,或与之相反,沉默原本应活化的基因,而这最终影响了个体对疾病的易感性。并且改变了的遗传物质有可能传给下一代,从而影响其对疾病的易感性,即由非DNA变异而改变表型的“可遗传的”现象[24,25]。遗传因素和环境因素,常用来解释过敏性疾病如何从一代传到下一代的风险程度。有相当多的证据支持疾病早期,甚至早到胚胎发育时期,即有疾病发生因素的影响。不仅母体暴露,如吸烟等,母亲上一代,甚至父系的环境暴露,均与过敏性哮喘等症状有一定的相关性。而这些现象或者可以从表观遗传学的机制中得到一些解释[26]。但如何从环境作用中将遗传因素剥离出来单独分析,对于很多慢性参与因素来说则仍然是一个难题。
卫生假说(Hygiene hypothesis)提示,诸如家庭生活环境、生育和喂养方式、微生物荷载、抗生素使用等,都对出生后是否发生过敏有作用[27]。这些诱发因素的作用机制之一,可能是通过对表观基因组学的修饰,进而影响了疾病发生发展甚至转归。而其中,相当部分研究显示,关键分子的DNA甲基化增加或减少,占据极大的作用[28,29]。研究发现,儿童哮喘轨迹可能在出生时就已经启动,包括在免疫调节和致炎通路中的表观遗传学修饰变化,而这些修饰均受到母体哮喘的影响[30]。一项追踪哮喘母亲的孩子历时9年的研究发现[30],9岁时发生或未发生哮喘,在其新生儿期的免疫细胞的589个不同的甲基化区域的表现上有着较明显的差异。其中SMAD3的甲基化水平,在哮喘孩子增高尤为明显,且与儿童哮喘风险相关。并且发现,在其母亲患哮喘的新生儿中,SMAD3甲基化水平与其天然炎性介质IL-1,呈现非常强的正相关[30]。6~12岁持发性过敏性哮喘儿童的外周血单个核细胞(Peripheral blood mononuclear cells,PBMCs)的DNA甲基化水平,包括几个和T细胞成熟、Th2免疫相关以及过氧化应激等相关的基因,均处于低甲基化水平[31]。表观遗传对疾病的作用,极有可能是通过对遗传信息的修饰达成的。如果这个假说是正确的,那么在下一代阻断过敏性哮喘的关键就可能是如何在孕期进行干预。有研究者对过敏家庭进行了表观遗传学相关的三代队列研究。结果发现,认为子代可能并不只是仅仅继承了亲代的表观基因组信息,进而传递到下一代,而更多则是遗传中发生的表观遗传放大效应,这可能更能解释表观遗传学参与过敏性疾病家族性发生的机制。提示在今后的研究中,可能更需要研究这些在亲子代间传递的表观遗传特征可能带来的功能性表现,以及建立这些现象的临床相关性对诱导性表观遗传性状转移模型的验证。
4 黏膜系统微生态与过敏性哮喘
许多证据表明,肠道微生态参与免疫耐受的形成及维持,并可能在其中有着重要作用[32]。该微生态可以被多种因素影响,如各种环境因素,包括饮食、抗生素使用、早期微生物暴露情况等。机体黏膜微生态的生物多样性的减少,以及肠道或皮肤菌群组成改变,与多种炎性疾病相关,包括过敏性疾病如哮喘、炎性肠病(Inflammatory bowel diseases,IBD)、Ⅰ型糖尿病等[33]。在一些免疫异常或免疫耐受异常人群,常发现其固有的微生态菌群的改变,或发生常见的菌群失调。而这种风险可能由于随后对肠道微生态有影响的生活方式而增大[34]。
研究显示,包括病原微生物在内的微生态群与哮喘发生和发展有很强的相关性[35]。从健康成人支气管冲刷物的分析发现,下气道也正常存在微生态,而并不是无菌的环境,并且发现哮喘病人或慢性阻塞性肺疾病的病人,其下气道的微生态组成较健康成人有所改变[36]。发生哮喘的儿童,在1周及1个月时,其肠道菌群多样性都远远低于那些后来没有发生哮喘的儿童,提示出生初期肠道菌群的多样性降低与之后发生哮喘有较高的相关性[37]。哮喘病人,特别是非肥胖哮喘病人,其肠道分泌组胺的细菌增多[38]。
肠道和肺部的微生态对哮喘的发生和进展均有影响。用新生小鼠的研究表明,某些肠道微生物与过敏性哮喘的发生有着很强的相关性。这些研究都提示,如何调整肠道及气道的微生态,特别是在生活早期,可能是预防或治疗哮喘的方法[39]。然而,用益生菌联合益生元在这个疾病上治疗的研究,总体结果模棱两可[40]。体外实验显示,益生菌可促进细胞分泌IL-10等下调免疫反应的介质,但人体实验的结果却没有像体外实验一样,提示体内的复杂性,远非体外细胞实验所能代表。例如,有研究显示在过敏性皮炎似乎有一定效果[41],但更多则并不理想,很难得到重复性的研究结果[40,42-44]。当然,这可能与选取的益生菌种类、组合、剂量等有关,并且研究对象的异质性,也对研究结果有一定影响。鉴于没有很好的临床前有效证据,在微生态与过敏性哮喘的相关研究及临床应用上,可能还需要更多系统性的研究数据的支撑[45]。
5 哮喘的表型与内表型
内表型(Endotype)是对重新认识哮喘分子异质性时提出的一个相对于疾病表型(Phenotype)的另一个概念,即由疾病确定的功能或病理机制来定义疾病的亚型[46]。而表型则是由疾病观察到的特征来定义,较少或并不涉及疾病的机制[46]。由于表型的复杂性,内表型的分类界定也尚不清晰。如果说某个内表型与某种特定疾病机制一一相关,则哮喘可能更多是某单一机制却可有多个内表型。尽管如此,这样进行分类无疑仍有助于优化治疗方案,并且强调从根据病人疾病表现为之选择正确的治疗方案(从表型出发),转变为根据特定治疗方案的作用机制来选择更适合的病人(从内表型或治疗型出发)[47]。
哮喘无论从临床或分子表型,都呈现出明显的异质性,而了解这些异质性,对于如何选择针对关键通路的阻断治疗方法,尤为重要。临床发现,如果考虑了哮喘表型特征,再选用相应的生物制剂,如细胞因子等的拮抗剂进行干预治疗,通常能取得明显效果。对于表现为高Th2免疫反应表型的一类哮喘,如一些修饰Th2免疫反应的方案,用于阻断IL-5、IL-13、IgE,或前列腺素D2,或一些其他信号通路干预等,可降低Th2型反应[48]。然而,如何鉴别到那些适合这些昂贵的生物干预治疗的病人,则需要实验室有快速、重复性好、特异性强的生物学指标。例如,对于糖皮质激素有效的难治性嗜酸性粒细胞增多性哮喘这样的表型,由于通常其中心环节是Th2型反应,故而选用针对Th2细胞因子的阻断剂有着很好的应用前景。而采用外周血嗜酸性粒细胞作为一个操作简单而重复性好的生物学指标,可能用来明确那些有特定表型的哮喘病人,指导选择生物治疗方案。
对于低Th2哮喘病人,目前还没有很明确的生物学指标,而仅仅依赖于检测不到高Th2反应指标来诊断。但发现低Th2哮喘可能与Th1细胞或Th17细胞相关[49]。而这些病人往往对激素治疗反应较差,其治疗手段的研究远远落后于高Th2哮喘。近来,研究发现在低Th2哮喘中,存在较高水平的中性粒细胞,或中性粒细胞相关介质[48]。而如何找到针对更多亚表型哮喘的特异性指标,则可能也是研究工作的热点之一[50]。如MeDALL(Mechan-isms of the Development of Allergy)的一项研究显示,在多发性过敏中,仅有不足40%是由于IgE致敏所致,由此提出非IgE致敏的一类多发过敏疾病。认为单因素致敏和多因素致敏可能也代表着两种不同的表型[51]。
6 过敏原特异性免疫治疗
世界卫生组织指出“免疫脱敏治疗是唯一可以彻底治疗过敏性疾病的根本性治疗方法”,即过敏原特异性免疫疗法(Allergen-specific immunoth-erapy,ASIT)。这是用少量过敏原提取物反复多次注射或经黏膜途径给药,让病人脱敏。该疗法的使用虽然已有近百年历史,机理仍不完全清楚,且疗效有很大的个体性差异。对于ASIT的研究和临床应用,2017年我国学者(主要针对过敏性鼻炎)和欧洲学者均发布了相应的指南[52,53]。明确ASIT作用机制,寻找更有效的耐受诱导原,如何进行个性化治疗,如何寻找有效生物标记识别对某种干预,治疗性抗原或复合物的应答性,均是研究热点。
在ASIT 的机制研究中,有发现经过ASIT治疗后,过敏部位的肥大细胞及嗜酸性粒细胞和嗜碱性粒细胞的数量有所下降,同时能增多调节性T细胞(T regulatory cell,Treg)及调节性B细胞(B regulatory cell,Breg)的数量,增加IL-10的分泌,抑制炎性树突细胞分泌细胞因子,炎性效应细胞如Th1、Th2及Th17细胞减少[54],还可能通过抑制CCL20及肺组织IL-33、CCL17、eotaxin水平等,拮抗过敏反应[55]。同时也有实验结果提示,在ASIT过程中,反复多次的过敏原刺激机体能产生多种类型亲合力各异的免疫球蛋白IgG[56],高亲合力IgG可能通过截获过敏原,而减少其被特异性的IgE抗体捕获。低亲合力IgG还可以和其抑制性受体FcγRIIB(免疫球蛋白IgG受体IIB)结合,通过特异性的IgG作为桥梁,FcεRI和抑制性的FcγRIIB得到交联,减弱肥大细胞的活化,降低机体对过敏的易感性[56]。这些研究从不同的角度逐步揭示ASIT作用机制的冰山一角,但ASIT的机理仍然属于探索阶段。例如ASIT治疗后肥大细胞数量减少的机理仍然不明,在ASIT中产生的大量IgG是否和治疗中发现的效应细胞肥大细胞数量减少有联系,细胞凋亡在ASIT的作用等等,都等待从不同角度不同环节的研究来回答和完善。对ASIT的机制更深入更细节的了解,不仅有助于设计更好的方法预防治疗过敏性疾病,其机制还可能对研究其他疾病的有效治疗方法,如自身免疫性疾病、器官移植、慢性感染、肿瘤等疾病、有一定的借鉴作用[57]。
选择预测监测或评估治疗有效性的生物指标,也是ASIT重要的内容。有推荐以特异性IgG4作为治疗敏感性,而特异性IgE/总IgE比率和IgE-FAB作为临床疗效指标[58],并且检测鼻腔液中的特异性IgE、IgG4及一些功能性抗体,与症状有更好的相关性,可能较外周血结果更能显示对治疗的反应性。此外,如嗜碱性粒细胞活化,一些可溶性细胞因子或趋化因子,及其他一些如DC、Treg、Breg等细胞学指标,这些研究结果还需要更多临床验证,并且还需要研究处于ASIT病人的靶器官的其他抗体,如IgA等[47]。
7 对更适合的实验动物的需求
过敏性疾病的研究在相当程度上依赖于实验动物模型,特别是对疾病发生发展机制的深入研究,以便开发有针对性的治疗手段。这些实验性研究无论从技术还是伦理角度,都很难直接在人体上来开展。目前已成功建立的疾病动物模型基本都是依赖于近交系实验小鼠。而其中绝大多数研究是基于常用的两个实验室近交系小鼠品系,即C57BL/6和BALB/c。由于这些近交系小鼠具有相同的遗传背景,大大方便了实验模型的可重复性和稳定性。目前生物医学界对免疫学的认识极大部分是基于对这些实验室小鼠的研究。然而,随着研究的深入,关于实验室小鼠是否能可靠地反映人体的免疫反应状态,日益受到医学科学界关注。对于将基于实验室小鼠的研究结果直接应用于推测人体免疫学反应机理,以及开发治疗手段的研究,都面临许多解释方面的困难。
实验室小鼠生活在相对洁净的屏障内,而环境对动物的免疫系统形成有着重要的影响。野生小鼠的免疫反应,对其进化适应性非常重要,因为他们需要不断适应其生活的环境得以更好生存。目前这个领域的研究较少。但已有的研究显示,野生动物的体内细胞免疫反应性远远高于实验室源动物。而对于体外培养的再刺激的反应,相对于实验室动物而言,则似乎处于免疫抑制状态[59]。比如,血清中免疫球蛋白水平,在不同环境生活的小鼠有着明显区别[60]。实验室小鼠的水平明显低于野生小鼠或者与野生小鼠共生活的小鼠,其中以IgE水平差异为甚,而后两者间则未见明显区别[60]。实验室小鼠和野生小鼠在NK细胞的生物学功能上的表现也有显著区别,如NK细胞数量、NK细胞经IL-2/IL-12刺激后表达IFN-γ的水平以及NK细胞活化状态,野生小鼠都明显高于实验室小鼠[61]。
8 结语
过敏性疾病患病率的增加,以及对生活质量更高要求,使该领域研究日益受到关注。研究技术及临床检测手段的更新带来更多研究思路的可行性,均给该领域研究带来更多机遇,同时也带来更多挑战。一些临床治疗方法的有效性及异质性,机制不十分明确,而一些很好的基础研究,研究成果如何进一步转化,在临床得到应用,需更多细节化探索。本领域的研究需要建立新型动物模型,结合生物信息学研究个性化发病机制及治疗机制、遗传及环境影响及交互作用,以及如何对临床检测及治疗指标进行有效评估。见图1。
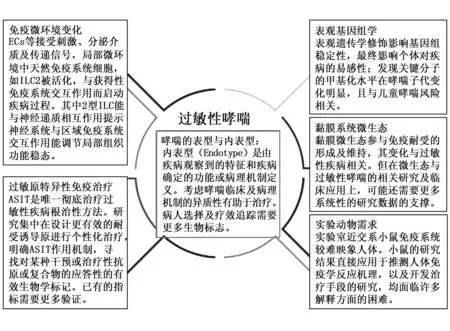
图1 过敏性哮喘研究热点Fig.1 Current research highlights in allergic asthma
参考文献:
[1] Martinez FD,Vercelli D.Asthma[J].Lancet(London,England),2013,382(9901):1360-1372.
[2] Sood S,Castro M.Asthma in 2016:reassured about the old,excited about the new[J].Lancet Respirat Med,2016,4(12):937-939.
[3] Ideker T,Sharan R.Protein networks in disease[J].Genome Res,2008,18(4):644-652.
[4] Lambrecht BN,Hammad H.The airway epithelium in asthma[J].Nat Med,2012,18(5):684-692.
[5] Besnard AG,Togbe D,Guillou N,etal.IL-33-activated dendritic cells are critical for allergic airway inflammation[J].Euro J Immunol,2011,41(6):1675-1686.
[6] Wawrzyniak P,Wawrzyniak M,Wanke K,etal.Regulation of bronchial epithelial barrier integrity by type 2 cytokines and histone deacetylases in asthmatic patients[J].J Allergy Clin Immunol,2017,139(1):93-103.
[7] van Rijt LS,Vos N,Willart M,etal.Persistent activation of dendritic cells after resolution of allergic airway inflammation breaks tolerance to inhaled allergens in mice[J].Am J Res Cri Care Med,2011,184(3):303-311.
[8] Wang P,Zhao W,Sun J,etal.Inflammatory mediators mediate airway smooth muscle contraction through a G protein-coupled receptor-transmembrane protein 16A-voltage-dependent Ca(2+)channel axis and contribute to bronchial hyperresponsiveness in asthma[J].J Allergy Clin Immunol,2017.pii:S0091-6749(17)31167-3.
[9] Halim TY,Krauss RH,Sun AC,etal.Lung natural helper cells are a critical source of Th2 cell-type cytokines in protease allergen-induced airway inflammation[J].Immunity,2012,36(3):451-463.
[10] Price AE,Liang HE,Sullivan BM,etal.Systemically dispersed innate IL-13-expressing cells in type 2 immunity[J].Proc Natl Acad Sci U S A,2010,107(25):11489-11494.
[11] Martinez-Gonzalez I,Steer CA,Takei F.Lung ILC2s link innate and adaptive responses in allergic inflammation[J].Trends Immunol,2015,36(3):189-195.
[12] Licona-Limon P,Kim LK,Palm NW,etal.TH2,allergy and group 2 innate lymphoid cells[J].Nature Immunol,2013,14(6):536-542.
[13] Madouri F,Chenuet P,Beuraud C,etal.Protein kinase Ctheta controls type 2 innate lymphoid cell and TH2 responses to house dust mite allergen[J].J Alle Clin Immunol,2017,139(5):1650-1666.
[14] Halim TY,Steer CA,Matha L,etal.Group 2 innate lymphoid cells are critical for the initiation of adaptive T helper 2 cell-mediated allergic lung inflammation[J].Immunity,2014,40(3):425-435.
[15] Gregory LG,Jones CP,Walker SA,etal.IL-25 drives remodelling in allergic airways disease induced by house dust mite[J].Thorax,2013,68(1):82-90.
[16] Oliphant CJ,Barlow JL,McKenzie AN.Insights into the initiation of type 2 immune responses[J].Immunology,2011,134(4):378-385.
[17] Neill DR,Wong SH,Bellosi A,etal.Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity[J].Nature,2010,464(7293):1367-1370.
[18] Wilhelm C,Hirota K,Stieglitz B,etal.An IL-9 fate reporter demonstrates the induction of an innate IL-9 response in lung inflammation[J].Nature Immunol,2011,12(11):1071-1077.
[19] Lund SJ,Portillo A,Cavagnero K,etal.Leukotriene C4 potentiates IL-33-induced group 2 innate lymphoid cell activation and lung inflammation[J].J Immunol,2017,199(3):1096-1104.
[20] De Grove KC,Provoost S,Hendriks RW,etal.Dysregulation of type 2 innate lymphoid cells and TH2 cells impairs pollutant-induced allergic airway responses[J].J Alle Clin Immunol,2017,139(1):246-257.e4.
[21] Wallrapp A,Riesenfeld SJ,Burkett PR,etal.The neuropeptide NMU amplifies ILC2-driven allergic lung inflammation[J].Nature,2017,549(7672):351-356.
[22] Klose CSN,Mahlakoiv T,Moeller JB,etal.The neuropeptide neuromedin U stimulates innate lymphoid cells and type 2 inflammation[J].Nature,2017,549(7671):282-286.
[23] Cardoso V,Chesne J,Ribeiro H,etal.Neuronal regulation of type 2 innate lymphoid cells via neuromedin U[J].Nature,2017,549(7671):277-281.
[24] Jablonka E,Raz G.Transgenerational epigenetic inheritance:prevalence,mechanisms,and implications for the study of heredity and evolution[J].Quar Rev Biol,2009,84(2):131-176.
[25] Prokopuk L,Western PS,Stringer JM.Transgenerational epigenetic inheritance:adaptation through the germline epigenome?[J].Epigenomics,2015,7(5):829-846.
[26] Arshad SH,Karmaus W,Zhang H,etal.Multigenerational cohorts in patients with asthma and allergy[J].J Allergy Clin Immun,2017,139(2):415-421.
[27] Liu AH.Revisiting the hygiene hypothesis for allergy and asthma[J].J Allergy Clin Immunol,2015,136(4):860-865.
[28] Kabesch M,Michel S,Tost J.Epigenetic mechanisms and the relationship to childhood asthma[J].Eur Res J,2010,36(4):950-961.
[29] Michel S,Busato F,Genuneit J,etal.Farm exposure and time trends in early childhood may influence DNA methylation in genes related to asthma and allergy[J].Allergy,2013,68(3):355-364.
[30] DeVries A,Wlasiuk G,Miller SJ,etal.Epigenome-wide analysis links SMAD3 methylation at birth to asthma in children of asthmatic mothers[J].J Allergy Clin Immunol,2017,140(2):534-542.
[31] Yang IV,Pedersen BS,Liu A,etal.DNA methylation and childhood asthma in the inner city[J].J Alle Clin Immunol,2015,136(1):69-80.
[32] Hooper LV,Littman DR,Macpherson AJ.Interactions between the microbiota and the immune system[J].Science,2012,336(6086):1268-1273.
[33] Gilbert JA,Quinn RA,Debelius J,etal.Microbiome-wide association studies link dynamic microbial consortia to disease[J].Nature,2016,535(7610):94-103.
[34] Haahtela T,Holgate S,Pawankar R,etal.The biodiversity hypothesis and allergic disease:world allergy organization position statement[J].World Allergy Organ J,2013,6(1):3.
[35] Edwards MR,Walton RP,Jackson DJ,etal.The potential of anti-infectives and immunomodulators as therapies for asthma and asthma exacerbations[J].Allergy,2018,73(1):50-63.
[36] Hilty M,Burke C,Pedro H,etal.Disordered microbial communities in asthmatic airways[J].PLoS One,2010,5(1):e8578.
[37] Abrahamsson TR,Jakobsson HE,Andersson AF,etal.Low gut microbiota diversity in early infancy precedes asthma at school age[J].Clin Exp Allergy,2014,44(6):842-850.
[38] Barcik W,Pugin B,Westermann P,etal.Histamine-secreting microbes are increased in the gut of adult asthma patients[J].J Allergy Clin Immunol,2016,138(5):1491-1494.e7.
[39] Chung KF.Airway microbial dysbiosis in asthmatic patients:A target for prevention and treatment?[J].J Allergy Clin Immunol,2017,139(4):1071-1081.
[40] Zuccotti G,Meneghin F,Aceti A,etal.Probiotics for prevention of atopic diseases in infants:systematic review and meta-analysis[J].Ann Allergy Asthma Immuonl,2015,70(11):1356-1371.
[41] Kalliomaki M,Salminen S,Poussa T,etal.Probiotics and prevention of atopic disease:4-year follow-up of a randomised placebo-controlled trial[J].Lancet(London,England),2003,361(9372):1869-1871.
[42] Kuitunen M,Kukkonen K,Juntunen-Backman K,etal.Probiotics prevent IgE-associated allergy until age 5 years in cesarean-delivered children but not in the total cohort[J].J Allergy Clin Immunol,2009,123(2):335-341.
[43] Cuello-Garcia CA,Brozek JL,Fiocchi A,etal.Probiotics for the prevention of allergy:A systematic review and meta-analysis of randomized controlled trials[J].J Allergy Clin Immunol,2015,136(4):952-961.
[44] Azad MB,Coneys JG,Kozyrskyj AL,etal.Probiotic supplemen-tation during pregnancy or infancy for the prevention of asthma and wheeze:systematic review and meta-analysis[J].BMJ(Clinical Researched),2013,347:f6471.
[45] Mennini M,Dahdah L,Artesani MC,etal.Probiotics in asthma and allergy prevention[J].Front Pediatr,2017,5:165.
[46] Green RH,Brightling CE,Woltmann G,etal.Analysis of induced sputum in adults with asthma:identification of subgroup with isolated sputum neutrophilia and poor response to inhaled corticosteroids[J].Thorax,2002,57(10):875-879.
[47] Pfaar O,Bonini S,Cardona V.Perspectives in allergen immuno-therapy:2017 and beyond[J].Ann Allergy Immunol,2018,73(Suppl 104):5-23.
[48] Stokes JR,Casale TB.Characterization of asthma endotypes:implications for therapy[J].Ann Allergy Asthma Immunol,2016,117(2):121-125.
[49] Kuo CS,Pavlidis S,Loza M,etal.T-helper cell type 2(Th2)and non-Th2 molecular phenotypes of asthma using sputum transcriptomics in U-BIOPRED[J].Euro Res J,2017,49(2),pii1602135.
[50] Walsh GM.Biologics targeting IL-5,IL-4 or IL-13 for the treatment of asthma-an update[J].Exp Rev Clin Immunol,2017,13(2):143-149.
[51] Anto JM,Bousquet J,Akdis M,etal.Mechanisms of the development of allergy(MeDALL):introducing novel concepts in allergy phenotypes[J].J Allergy Clin Immunol,2017,139(2):388-399.
[52] Bao Y,Chen J,Cheng L,etal.Chinese Guideline on allergen immunotherapy for allergic rhinitis[J].Journal of thoracic disease,2017,9(11):4607-4650.
[53] Halken S,Larenas-Linnemann D.EAACI guidelines on allergen immunotherapy:prevention of allergy[J].Pediatr Allergy Immunol,2017,28(8):728-745.
[54] Akdis CA.Therapies for allergic inflammation:refining strategies to induce tolerance[J].Nat Med,2012,18(5):736-749.
[55] Hesse L,van Ieperen N,Habraken C,etal.Subcutaneous immunotherapy with purified Der p1 and 2 suppresses type-2 immunity in a murine asthma model[J].Allergy,2018.doi:10.1111/all.13382.
[56] Zha L,Leoratti FMS,He L,etal.An unexpected protective role of low-affinity allergen-specific IgG through the inhibitory receptor FcγRⅡb[J].J Allergy Clin Immunol,2018,doi:10.1016/j.jaci.2017.09.054.
[57] Akdis M,Akdis CA.Mechanisms of allergen-specific immunoth-erapy:multiple suppressor factors at work in immune tolerance to allergens[J].J Allergy Clin Immunol,2014,133(3):621-631.
[58] Shamji MH,Kappen JH,Akdis M,etal.Biomarkers for monitoring clinical efficacy of allergen immunotherapy for allergic rhinoconjunctivitis and allergic asthma:an EAACI Position Paper[J].Allergy,2017,72(8):1156-1173.
[59] Abolins S,King EC,Lazarou L,etal.The comparative immunology of wild and laboratory mice,Mus musculus domesticus[J].Nat Communol,2017,8:14811.
[60] Beura LK,Hamilton SE,Bi K,etal.Normalizing the environment recapitulates adult human immune traits in laboratory mice[J].Nature,2016,532(7600):512-516.
[61] Boysen P,Eide DM,Storset AK.Natural killer cells in free-living Mus musculus have a primed phenotype[J].Molecular Ecology,2011,20(23):5103-5110.