磷脂二酰甘油酰基转移酶(PDAT)基因的克隆与表达分析
2018-04-16潘丽娟陈明娜禹山林梁成伟迟晓元
徐 赫,潘丽娟,陈 娜,陈明娜,王 冕,王 通,禹山林,梁成伟,迟晓元*
(1.青岛科技大学海洋科学与生物工程学院,山东 青岛 266042; 2.山东省花生研究所,山东 青岛 266100)
花生是我国重要的油料作物和经济作物,是重要的植物油脂和蛋白质来源,在农业生产和国民经济中的地位日益显著[1-2]。花生中的三酰甘油(triacylgycerol,TAG)主要由两条途径合成,其中一条便是PDAT途径。二酰甘油(diacylglycerol, DAG)在磷脂二酰甘油酰基转移酶(phospholipids:diacylglycerol acyltransferase,PDAT)的催化条件下与磷脂(phospholipids, PC)反应生成TAG和溶血磷脂(lyso-phospholipids, LPC)[3-5]。
PDAT基因已经在拟南芥[6]、水稻[7-8]、玉米[9]、亚麻、花生[10-11]等植物中被克隆出来。然而PDAT在不同物种中的功能存在差异性。Pan[12]等从亚麻的种子中克隆出一组PDAT基因,在酵母和拟南芥中高度表达,可使其中的亚麻酸成分快速升高。在绿色衣藻中,其PDAT拥有酰基转移酶及脂肪酶的两种活性,体内膜脂分解的过程中,伴随TAG的产生,在各种胁迫条件下得以更好地生存[13]。
为探索PDAT蛋白在花生油脂合成与表达的意义,本文克隆得到2个蛋白的完整基因,分别命名为AhPDAT1和AhPDAT2。其基因登录号分别为KJ704777,MH790267。
1 材料与方法
1.1 植物材料
山东省花生研究所提供的花育33号。花生生长到三叶期后进行几种激素与非生物胁迫处理。具体参照郝翠翠等[14]的方法。
1.2 总RNA提取与cDNA合成、目的基因的扩增
拟南芥两个蛋白序列是从NCBI上搜索到的,分别是AtPDAT1(AT5G13640)和AtPDAT2(AT3G44830)。从花生cDNA文库里发现了2个编码磷脂二酰甘油酰基转移酶的基因的全长序列:AhPDAT1和AhPDAT2。根据上述序列,设计出基因的完整引物。引物序列AhPDAT1-F:5’-ATGTCGTTTCTGCGCCGC-3’;AhPDAT1-R:5’-CTAGAGCTTTAAATTAATATTTTC-3’;AhPDAT2-F:5’-ATGTCTTCAGTTCGACGGAG-3’;AhPDAT2-R:5’-TCACAGGCGTAACTTAATTTTC-3’。具体参照陈娜等[15-16]的方法。模板采用cDNA,通过PCR扩增得到目的基因,具体参照郝翠翠等[14]的方法。
1.3 氨基酸序列分析
蛋白质的基本参数-ProtParam;蛋白跨膜结构-TMHMM 2.0 Server;蛋白的二级结构-SCRATCH Protein Predicter Tool。
1.4 多序列比对与系统发育分析
研究PDAT所存在的进化关系,建立一个多基因组的数据库。这12个基因组分别为:拟南芥(Arabidopsisthaliana)基因组,花生野生种(Arachisduranensis)基因组,花生野生种(Arachisipaensis)基因组,花生栽培种(Arachishypogaeacv. Tifrunner)基因组,大豆(Glycinemax)基因组,水稻(Oryzasativa)基因组,短柄草(Brachypodiumdistachyon)基因组,狗尾草(Setariaitalica)基因组,小立碗藓(Physcomitrellapatens)基因组,江南卷柏(Selaginellamoellendorffii)基因组,莱茵衣藻(Chlamydomonasreinhardtii)和团藻(Volvoxcarteri)基因组数据。基因组数据从peanutbase数据库下载获得。设定E值<1e-10[17]。构建系统发育树采用MEGA 6.0软件的Neighbour-Joining法。
1.5 荧光定量PCR
PCR反应按陈娜等[15-16]的方法。采用Actin11为内参基因,实验数据使用2-△△Cp方法来分析[18]。荧光定量的引物:
qAhPDAT1-F:5’- GAGAAGGAGAAGGAGGAGGAAGAG -3’,
qAhPDAT1-R:5’- ACCACCAGAGGGAGCATATCAC-3’;
qAhPDAT2-F:5’- GCCGAAGAGAACAACAAAGAAGAAG -3’,
qAhPDAT2-R:5’-AGAACAGCATCAACCACCACAC -3’;
qACT11-F:5’- TTGGAATGGGTCAGAAGGATGC -3’;
qACT11-R:5’- AGTGGTGCCTCAGTAAGAAGC -3’。
2 结果与分析
2.1 AhPDAT1和AhPDAT2基因的克隆
编码磷脂二酰甘油酰基转移酶的基因的完整序列是从花生cDNA文库中克隆获得的,它们与拟南芥AtPDAT1和AtPDAT2基因序列相似性很高。其中,AhPDAT1基因全长2103 bp,编码700个氨基酸;AhPDAT2基因全长2046 bp,编码681个氨基酸 (图1、图2)。

图1 AhPDAT1基因的核苷酸及氨基酸序列Fig.1 Nucleotide sequence and deduced amino acid sequence of AhPDAT1
2.2 氨基酸序列分析
对AhPDAT1和AhPDAT2基因编码的蛋白质的理化性质进行了分析。结果表明,AhPDAT1蛋白的理论分子量为78.72 kD,理论等电点为8.66。AhPDAT1包含的700个氨基酸残基中,Gly(G)含量最高,占总氨基酸的9.1%,其次为Lys(K),Ala(A),Leu(L),分别占总氨基酸的8.0%,7.3%,7.3%。带负电的氨基酸残基有88个(Asp + Glu),带正电的氨基酸残基有97个(Arg + Lys)。亲水指数为0.465,不稳定系数是40.74,属于不稳定蛋白。TMHMM在线工具预测发现,AhPDAT1有1个跨膜结构,推测AhPDAT1属于跨膜类蛋白。SCRATCH Protein Predicter Tool结果显示:AhPDAT1含α-螺旋197个,占28.14 %;β-折叠63个,占9.00 %;无规则卷曲440个,占62.86 %。
AhPDAT2蛋白的理论分子量为76.32 kD,理论等电点为6.17。AhPDAT2包含的681个氨基酸残基中,Gly(G)含量最高,占总氨基酸的9.3%,其次为Glu(E),Ala(A),Lys(K),分别占总氨基酸的7.8%,7.6%,6.8%。带负电的氨基酸有88个(Asp + Glu),带正电的氨基酸有81个(Arg + Lys)。亲水指数0.409,不稳定系数44.48,属于不稳定蛋白。TMHMM在线工具预测发现,AhPDAT2有1个跨膜结构,推测AhPDAT2属于跨膜类蛋白。SCRATCH Protein Predicter Tool结果显示:AhPDAT2含α-螺旋191个,占28.05 %;β-折叠65个,占9.54 %;无规则卷曲425个,占62.41 %。
采用拟南芥、野生种和栽培种花生中PDATs蛋白家族氨基酸序列进行多序列比对分析(图3)。结果显示,AhPDAT1氨基酸序列与Arahy.F78IM4.1、Aradu.UA9D8.1、Araip.WVH6X.1氨基酸序列相似性较高,分别为100%、86.6%、88.2%;与AT3G44830.1,AT5G13640.1氨基酸序列相似性较低,分别为55.1%和69.3%。AhPDAT2氨基酸序列与Araip.I19GZ.1、Aradu.S9XBY.1、Arahy.91WX57.1的氨基酸序列相似性较高,分别为99.7%、94.0%、92.45%;与AT3G44830.1,AT5G13640.1氨基酸序列的相似性较低,分别为54.5%和73.2%。因此,AhPDAT1与AhPDAT2编码的蛋白与野生种花生及拟南芥同源性较大,可能来自于同一家族。

图2 AhDPAT2基因的核苷酸及氨基酸序列 Fig.2 Nucleotide sequence and deduced amino acid sequence of AhPDAT2

图3 花生AhPDAT1和AhPDAT2蛋白的多序列比对结果Fig.3 Comparison of AhPDAT1 and AhPDAT2 amino acid sequence among peanut and other species 注:保守结构域用方框标出。 Note: The conserved domain was shownwith boxes.

图4 花生AhPDAT1和AhPDAT2的基因结构 Fig.4 The schematic diagram of the structure of AhPDAT1 and AhPDAT2
AhPDAT1和AhPDAT2基因组全长分别为3918 bp和5630 bp。GSDS工具作基因结构分析,发现AhPDAT1和AhPDAT2的基因结构都含有5个内含子和6个外显子。
2.3 系统发育树分析
如图5所示,从系统发育树可以看出,花生AhPDAT1、AhPDAT2基因与拟南芥AT5G13640基因聚在一起,拟南芥AT3G44830基因与大豆基因聚在一起。真核绿藻莱茵衣藻和团藻的PDAT基因位于树的基部,它们可能是高等植物PDAT基因的来源。

图5 植物PDAT基因系统发育树 Fig.5 Phylogenetic tree of plant PDAT isoforms

图6 AhPDAT1和AhPDAT2基因在花生不同组织和种子不同发育时期的表达分析Fig.6 Expression analysis of AhPDAT1 and AhPDAT2 in seven peanut tissues and at five stages of seed development

图7 AhPDAT1基因在不同非生物逆境胁迫和激素处理下的表达分析 Fig.7 Expression analysis of AhPDAT1 under abiotic stresses and hormone treatments
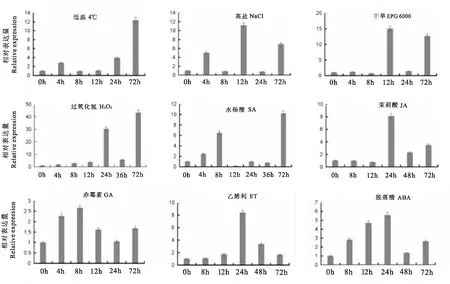
图8 AhPDAT2基因在不同非生物逆境胁迫和激素处理下的表达分析Fig.8 Expression analysis of AhPDAT2 under abiotic stresses and hormone treatments
2.4 基因的表达特性分析
对AhPDAT1和AhPDAT2基因采用实时荧光定量PCR技术分析其特性。内参基因选用花生Actin11基因。结果显示(图6),AhPDAT1基因在种子中的表达量最高;在种子生长周期中,最后一个发育时期有着明显的表达量。AhPDAT2基因在花生的花中表达量最高,其次为下胚轴和叶,在种子中表达量最低。
结果显示(图7),以0h为参考,在各因子影响下,花生叶片中AhPDAT1基因表达量有不同表达,表明该基因可能参与了花生逆境胁迫抗性途径。其中,低温处理后24 h时,表达量大幅上调,为0 h对照的11倍。高盐和干旱处理后12 h时基因表达量最高,分别为0 h对照的40倍和8倍。过氧化氢处理24 h和72h时,表达量明显,其余时期表达量极低。水杨酸处理8 h表达量最高,是0 h对照的16倍。茉莉酸处理24 h时表达量最高,为0 h对照的5.5倍。赤霉素处理72h后表达量达到最大,为0 h对照的6倍。乙烯利处理48 h时表达量最大。脱落酸处理后,基因表达量逐渐上调,12h后显著下降。
结果显示(图8),以0h为参考,在各因子影响下,花生叶片中AhPDAT2基因表达量不同。其中,低温处理后72 h时表达量最高,远高于其他时期。高盐在12h表达量最高为0 h对照的11倍,干旱在12h和72h表达量更明显。过氧化氢和水杨酸处理后72 h时基因表达量最高,分别为0 h对照的43倍和10倍。茉莉酸处理24 h时表达量最高,为0 h对照的8倍。赤霉素处理8h后表达量达到最大,为0 h对照的2.6倍。乙烯利处理和脱落酸处理表达量均在24h最大,分别为0 h对照的8倍和6倍。
3 讨 论
PDAT基因起源很早,在植物和细菌中分布广泛,所编码的蛋白与DGAT基因所编码的蛋白在功能上近似,但目前对于PDAT的研究远滞后于DGAT。前期研究发现,PDAT不仅具有TAG合成的功能[19-20],还在应对非生物胁迫中也有重要作用[21-22]。
本实验克隆得到的AhPDAT1和AhPDAT2基因都属于PDAT家族,通过进化树分析可以看出二者都与大豆PDAT基因有着比较近的亲缘关系。莱茵衣藻和团藻等真核绿藻位于系统发育树的根部,因此推测可能为进化的来源。这两个PDAT基因均含有6个外显子,内含子相位高度保守,说明它们在功能上都具有保守性。根据资料显示一些保守基序在一少部分植物丢失了,而高度保守的特点在被子植物PDAT蛋白序列更为普遍。PDAT基因功能多样性和底物特异性的有效证据之一就是不同物种PDAT基因的表达方式与结构特性差异明显[23-24]。
荧光定量结果显示AhPDAT1基因在种子中的表达量最高,其余组织表达量都很少;而AhPDAT2基因在花生下胚轴中的表达量最高。AhPDAT1和AhPDAT2基因分别在果针下地后60d和36d的表达量远高于其余各时期。此外干旱、高盐、低温等因素制约植物的产量和生长发育[23-24]。本研究发现AhPDAT1和AhPDAT2两基因对9类胁迫均有响应,但是响应模式有所不同。总之,本研究有助于阐明PDAT基因在花生油脂合成途径、抗逆等方面的功能,可以为花生育种提供新的基因资源。
