基于数字化标本和种群形态变异特征试论块茎堇菜的分类地位
2018-04-10侯云云王亚莉
苏 雪 侯云云,2 高 婷 王亚莉 孙 坤*
(1.西北师范大学生命科学学院,兰州 730070; 2.富平中学,渭南 711700)
块茎堇菜(ViolatuberiferaFranchet.)和鳞茎堇菜(V.bulbosaMaxim.)为堇菜科(Violaceae)堇菜属(ViolaL.)合生托叶组(Sect. Adnatae)的两种多年生草本植物,主要分布于青藏高原及其毗邻的高山地区,也是该地区两种特有植物[1]。块茎堇菜和鳞茎堇菜不仅具有相似的地理分布,而且在形态结构上也较为相似,尤其是它们具有相同的花部特征和繁殖特性,均可通过形成开放花、地上闭锁花以及着生于地下匍匐茎上的闭锁花和鳞茎进行繁殖[1~2],这也致使Stapf[3]在比较两者模式标本后认为它们是同一种植物。然而,由于块茎堇菜和鳞茎堇菜的叶片形态存在差异,块茎堇菜叶片通常为肾形、基部心形、先端钝,而鳞茎堇菜叶片为长圆状卵形、基部楔形或浅心形、先端急尖,因此Becker W[4]和王庆瑞[1]均将块茎堇菜和鳞茎堇菜作为两个不同的种处理。叶片作为最可塑的器官,易受环境饰变的影响[5],因此,陈又生等[6]在编写《Flora of China》时,将块茎堇菜重新归入鳞茎堇菜。可见,块茎堇菜的分类地位一直以来都存在较大争议。
尽管叶片形态的可塑性使其在分类上存在一定局限性,但叶片形态作为植物形态结构的重要组成部分,仍广泛应用于传统植物分类学中[7~9]。虽然,孙坤等[10]在堇菜属数量分类研究中认为块茎堇菜和鳞茎堇菜的叶片形态性状存在差异,应作为两个不同的种处理,但迄今对于块茎堇菜和鳞茎堇菜叶片形态特征的变异规律并不清楚。因此,本文拟通过对数字化标本和同域分布的鳞茎堇菜和块茎堇菜种群叶片形态变异的深入研究,探讨块茎堇菜和鳞茎堇菜叶片形态的分类学价值,以期为揭示块茎堇菜的分类地位提供证据。
1 材料和方法
1.1 材料
研究材料分为两部分:(1)对中国科学院植物研究所标本馆(PE)、中国科学院昆明植物所标本馆(KUN)、中国科学院西北高原生物研究所标本馆(QTPMB)和中国科学院成都生物研究所标本馆(CDBI)等标本馆内现存块茎堇菜和鳞茎堇菜所有叶形完整的数字化标本植株进行测量和统计;(2)在块茎堇菜和鳞茎堇菜同域分布的青藏高原东缘甘肃省合作市,分别选取块茎堇菜和鳞茎堇菜各3个种群,包括合作草原站附近(各80株)、合作扎油沟(各30株)和合作完冒乡(各30株),每个种群选取大小基本一致的个体,每个个体选取4~6片叶测量。
1.2 方法
利用Adobe photoshop软件中的标尺对数字标本馆下载获得的标本图片进行叶片形态参数的测量和统计。由于块茎堇菜和鳞茎堇菜叶形差异主要集中于叶片基部和先端形状处,故我们选取以下反映叶片形态性状的特征作为统计指标:最大叶片长度(L)、最大叶片宽度(W)、叶柄长(PL)和长宽比(LWR)、叶片最宽处到顶部距离(AL)、叶片最宽处到基部距离(BL)。块茎堇菜和鳞茎堇菜自然种群的叶片形态参数采用游标卡尺测量。为更好地反映叶片形态特征,在数字化标本测量指标基础上多选取以下参数:1/5最大叶片长度处叶片宽度(W′)、△L(L-L′)、△W(W-W′)。块茎堇菜和鳞茎堇菜的基本形态及测量参数见图1。
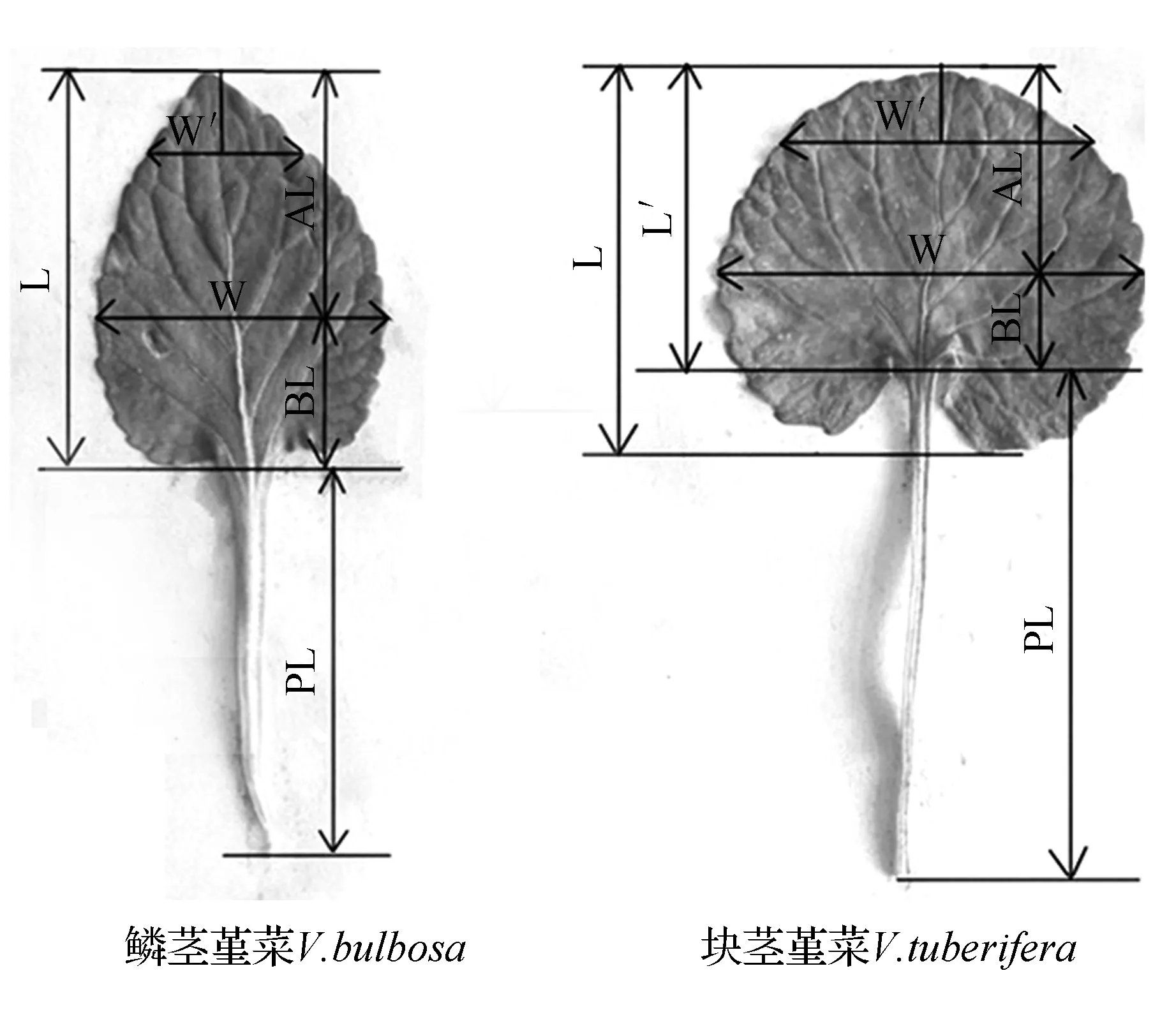
图1 块茎堇菜和鳞茎堇菜叶片形态及测量参数L.最大叶片长度;L′.从基部到顶部的叶片长度;W.最大叶片宽度;W′.1/5最大叶片长度处的叶片宽度;BL.叶片最宽处到基部的距离;AL.叶片最宽处到顶部的距离;PL.叶柄长度Fig.1 Leaf morphology and measurement parameters of V.bulbosa and V.tuberiferaL. Maximum blade length; L′. Blade length from base to top; W. Maximum blade width; W′. Blade width from the base to the point of 1/5 maximum blade length; BL. Blade length form the base to the point of maximum width; AL. Apex length from the base to the point of maximum width; PL. Petiole length
1.3 数据处理
测量所得叶片形态数据利用Excel统计分析其最大值、最小值、平均值、标准差和变异系数,用SPSS20.0进行单因素方差分析(ANOVA),在9个叶片形态参数基础上进行主成分分析(PCA),用Origin 9.0软件作图。
2 结果与分析
2.1 基于数字化标本叶片形态特征对块茎堇菜和鳞茎堇菜的主成分分析
主成分分析结果表明,所有数字化标本按照叶形特征明显分为两大叶形不同的集群,一个集群为叶片肾形,基部心形,先端钝,符合块茎堇菜的叶片形态;另一个集群为叶片长圆状卵形,基部楔形或浅心形,先端急尖,符合鳞茎堇菜的叶形特征(图2)。第一主成分贡献的叶片特征主要是L、W、BL和LWR值,贡献率为73.375%;第二主成分贡献的叶片特征主要是AL和PL值,贡献率为19.298%,这些叶片形态参数可基本反映块茎堇菜和鳞茎堇菜的叶片形态差异。
2.2 基于数字化标本块茎堇菜和鳞茎堇菜的叶片形态变异特征
对不同地区块茎堇菜和鳞茎堇菜数字化标本叶片形态变异的分析发现,各地区块茎堇菜和鳞茎堇菜的叶片形态存在一定变异,反映叶片大小的形态参数,如L、W、AL、BL、PL等在各个地区变异系数均较大(21.66%~76.85%),但反映叶形的参数LWR值在各个地区的变异系数均相对较小(7.99%~17.80%)。叶片形态特征参数差异性分析表明,W、BL和LWR在不同地区的同种堇菜间没有显著性差异,但在块茎堇菜和鳞茎堇菜间却存在显著性差异,其中LWR在两种堇菜间差异极显著(P<0.01)(表1)。因此,块茎堇菜和鳞茎堇菜的叶形特征是一个相对稳定且受环境影响较小的性状,可用作其分类的依据,其中W、BL和LWR的分类价值较大,特别是LWR作为反映叶片的基本形状的叶形参数,能反映出块茎堇菜和鳞茎堇菜的叶形差异。
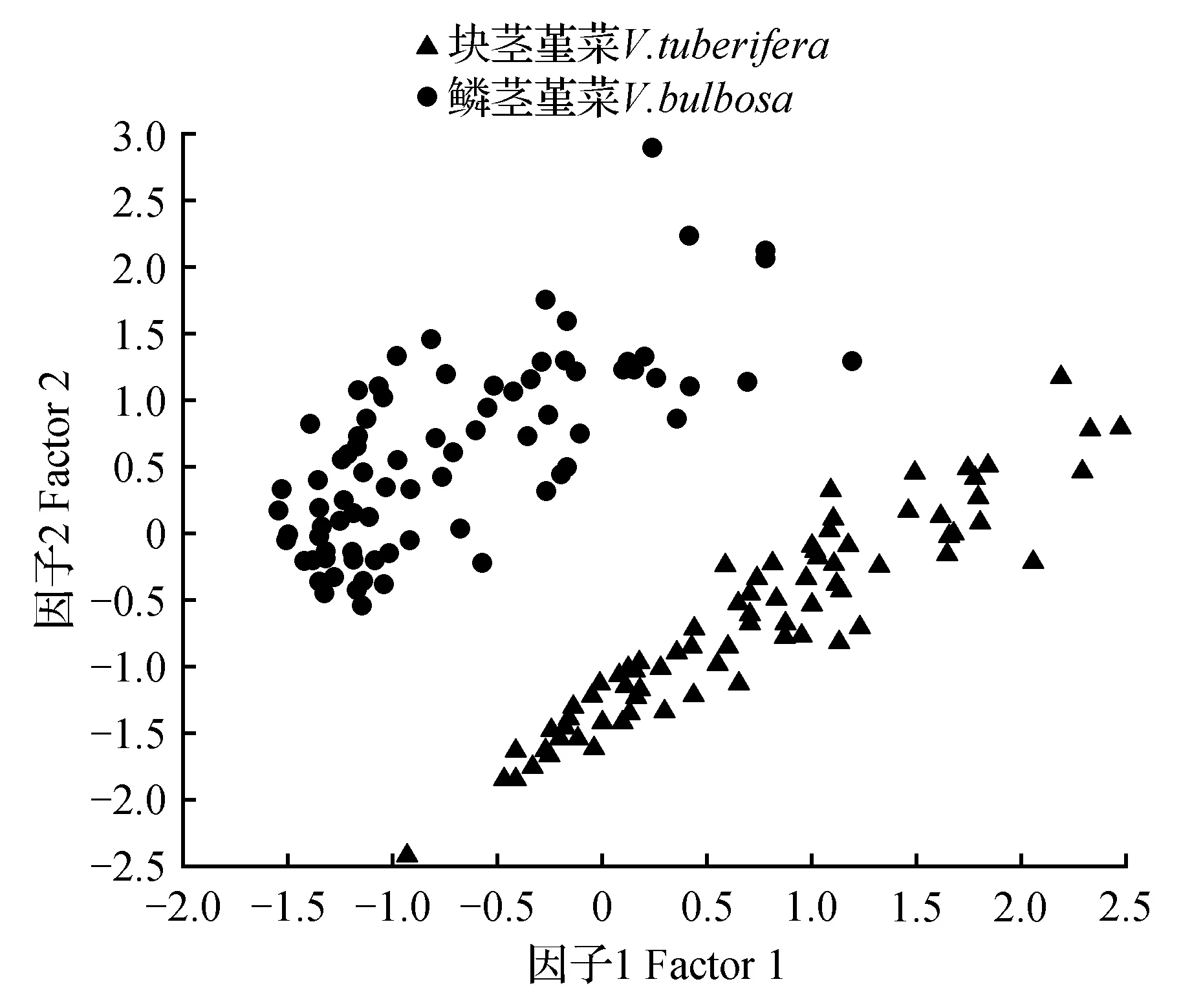
图2 基于数字化标本叶片形态对块茎堇菜和鳞茎堇菜的主成分分析Fig.2 The PCA of V.bulbosa and V.tuberifera based on leaf morphology of digital specimen
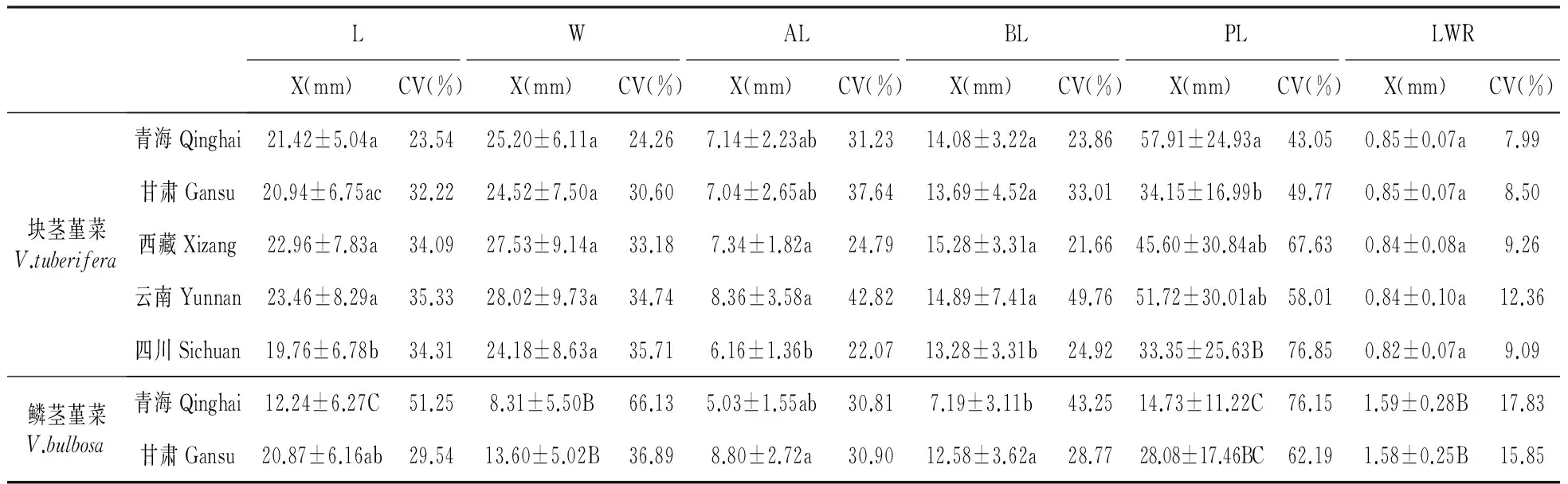
LWALBLPLLWRX(mm)CV(%)X(mm)CV(%)X(mm)CV(%)X(mm)CV(%)X(mm)CV(%)X(mm)CV(%)块茎堇菜V.tuberifera青海Qinghai21.42±5.04a23.5425.20±6.11a24.267.14±2.23ab31.2314.08±3.22a23.8657.91±24.93a43.050.85±0.07a7.99甘肃Gansu20.94±6.75ac32.2224.52±7.50a30.607.04±2.65ab37.6413.69±4.52a33.0134.15±16.99b49.770.85±0.07a8.50西藏Xizang22.96±7.83a34.0927.53±9.14a33.187.34±1.82a24.7915.28±3.31a21.6645.60±30.84ab67.630.84±0.08a9.26云南Yunnan23.46±8.29a35.3328.02±9.73a34.748.36±3.58a42.8214.89±7.41a49.7651.72±30.01ab58.010.84±0.10a12.36四川Sichuan19.76±6.78b34.3124.18±8.63a35.716.16±1.36b22.0713.28±3.31b24.9233.35±25.63B76.850.82±0.07a9.09鳞茎堇菜V.bulbosa青海Qinghai12.24±6.27C51.258.31±5.50B66.135.03±1.55ab30.817.19±3.11b43.2514.73±11.22C76.151.59±0.28B17.83甘肃Gansu20.87±6.16ab29.5413.60±5.02B36.898.80±2.72a30.9012.58±3.62a28.7728.08±17.46BC62.191.58±0.25B15.85
注:同一列同一字母代表差异不显著;不同小写字母代表差异显著(P<0.05);不同大写字母代表差异极显著(P<0.01)X.平均值±标准差;CV.变异系数下同。
Note:The same column of the same letter represents no significant difference,different lowercase letters represent significant differences(P<0.05) different uppercase letter represent extremely significant differences(P<0.01)X. Mean±Standard deviation; CV. Coefficient of variationThe same as below.
2.3 种群水平上块茎堇菜和鳞茎堇菜叶片形态变异
为进一步了解块茎堇菜和鳞茎堇菜间的叶片形态变异特征,我们以小地理尺度的几个同域种群为例,分析了种群水平的叶片形态变异特征(表2)。结果表明,块茎堇菜和鳞茎堇菜种群内的叶片形态存在一定变异,其中△L和PL变异系数相对较高,但反映叶片基部和先端形状的叶形参数△W、AL和BL的变异系数较低,特别是反映叶片基本形状的参数LWR,其种群内变异系数均小于7%。这一结果说明两种堇菜的叶片大小虽存在一定变异,但其叶片形态却没有明显变化。种群间的变异分析表明,鳞茎堇菜或块茎堇菜不同种群间的W、△W、AL、BL和LWR差异不显著,而且在种群内或种群间相对稳定的叶形参数在两种堇菜间也差异显著,如合作扎油沟地区的块茎堇菜和鳞茎堇菜间的叶片形态参数L、W、W′、△L、AL、BL和LWR值在两种堇菜间均是差异极显著(P<0.01),△W差异显著(P<0.05),完冒乡和草原站结果也与扎油沟类似。可见,两种堇菜叶片形态的主要差异是存在于种间而并非种群间或种群内部。
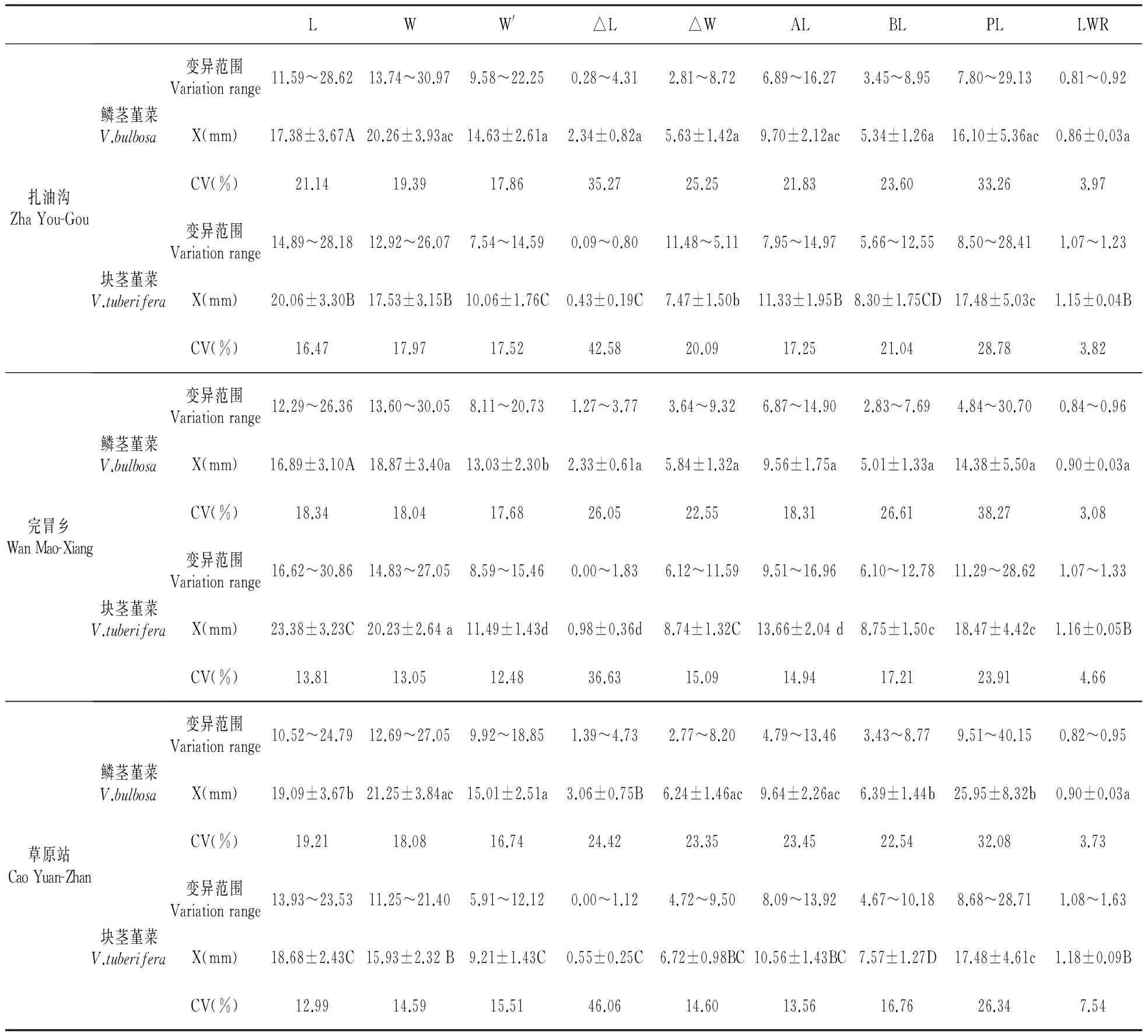
表2 种群水平上块茎堇菜和鳞茎堇菜叶片形态变异
2.4 种群水平上基于叶片形态的块茎堇菜和鳞茎堇菜的主成分分析
块茎堇菜和鳞茎堇菜几个同域自然种群叶片形态变异特征主成分分析结果显示,所有自然种群个体根据叶片形态分为两个集群,块茎堇菜聚集在一处,鳞茎堇菜聚集在另一处,没有混杂现象(图3),说明没有中间叶形的存在,这与数字化标本的分析结果完全一致。可见,同域生长的块茎堇菜和鳞茎堇菜其叶片形态在种内不同种群间的分化不大,主要表现为种间的差异。
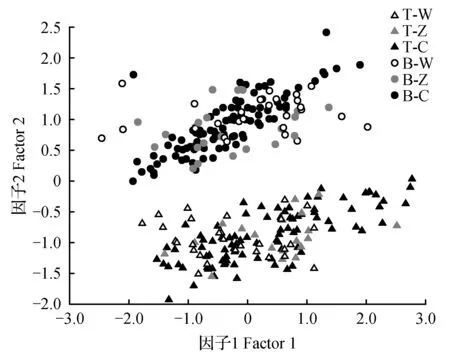
图3 种群水平上基于叶片形态的块茎堇菜和鳞茎堇菜的主成分分析 T-W.完冒乡块茎堇菜;T-Z.扎油沟块茎堇菜;T-C.草原站块茎堇菜;B-W.完冒乡鳞茎堇菜;B-Z.扎油沟鳞茎堇菜;B-C.草原站块茎堇菜Fig.3 The PCA of the populations of V.bulbosa and V.tuberifera based on leaf morphological characters T-W. Represents V.tuberifera in Wan Mao-Xiang; T-Z. Represents V.tuberifera in Zha You-Gou; T-C. Represents V.tuberifera in Cao Yuan-Zan; B-W. Represents V.bulbosa in Wan Mao-Xiang; B-Z. Represents V.bulbosa in Zha You-Gou; B-C. Represents V.bulbosa in Cao Yuan-Zan
3 讨论
叶片形态,包括形状和大小,叶缘、叶基和先端形状,叶片分裂程度,叶脉分布式样等因植物种类不同而不同,这些形态特征在植物分类学上具有较大意义[11],系统研究叶片形态特征差异可为植物分类鉴定提供证据[12~13]。然而,叶片作为植物进化过程中最具可塑性器官,其形态也易受环境的影响[14],如Hovenden & Schoor[15]对山毛榉叶形态研究表明海拔是影响叶形态的重要因素。因此,深入研究植物的叶片形态变异特征及其与环境的关系可以为揭示叶片性状的分类学价值提供依据。数字化标本馆为研究不同地区植物形态变异提供了一个重要的便利平台[16]。我们利用数字化标本对块茎堇菜和鳞茎堇菜整个分布区标本叶片形态变异的主成分分析结果表明,所有标本按叶形分为两个集群,分别对应不同的叶形。对块茎堇菜和鳞茎堇菜叶片形态变异特征的比较分析发现,同一种内反映叶形的W、BL和LWR值在各地区内或不同地区间均相对稳定、但在两种堇菜间却存在显著性差异,这说明两种堇菜间的叶形差异可能并非由于环境条件导致的环境饰变,而是由遗传因素决定的。
植物受相同环境因子的驱动会表现出相似的形态特征[17~18]。本文的数字标本信息虽然来自块茎堇菜和鳞茎堇菜的整个分布区,但因标本数量有限,因而在种群水平分析同域分布两种堇菜的叶片形态变异可以进一步揭示其叶片变异特征和叶片形态作为其分类依据的可靠性。我们对小地理尺度同域分布的两种堇菜自然种群叶片形态变异的研究表明,反映叶片基部和先端特征及叶形的参数△W、AL、BL以及LWR值在种群内的变异系数较低。两种堇菜种内种群间在叶片形态上虽存在一定变异,但存在于种间的叶形差异更加显著,如在种群内或种群间相对稳定的叶形参数W、△W、AL、BL和LWR值,在块茎堇菜和鳞茎堇菜间却表现出了极显著差异。主成分分析结果也表明两种堇菜的叶片形态在种内分化不大,主要表现为种间的差异。这与孙坤等[10]基于数量分类学的研究和苏雪[19]对同一地区块茎堇菜和鳞茎堇菜遗传结构的研究结果一致。综上可见,块茎堇菜和鳞茎堇菜的叶片形态差异在种间相对稳定,W、△W、AL、BL和LWR等反映其叶片形态特征的参数有一定分类价值。分析表明,块茎堇菜叶片通常为肾形,先端钝、基部心形,而鳞茎堇菜叶片则通常为长圆状卵形,先端急尖、基部楔形或浅心形。因此,建议将块茎堇菜作为鳞茎堇菜的亚种或独立的种处理。
1.王庆瑞.中国植物志[M].北京:科学出版社,1991.
Wang Q R.Flora of China[M].Beijing:Science Press,1991.
2.Maximowicz C J.Diagnoses plantarum novarum asiaticarum[J].Melanges Biologiques Tirés du Bulletin(Physico-Mhatematique de) l’Académie Impèriale des Sciences de St.-Pétersbourg,1876,(9):707-831.
3.Stapf O.Bulbous violet in the Himalayas[J].Bulletin of miscellaneous information(Royal Botanic Gardens,Kew),1894,(94):368-370.
4.Becker W.Violae Asiaticae et Australenses III[J].Beihefte zum Botanischen Centralblatt,1918,36:15-59.
5.Picotte J J,Rhode J M,Cruzan M B.Leaf morphological responses to variation in water availability for plants in thePiriquetacarolinianacomplex[J].Plant Ecology,2009,200(2):267-275.
6.Chen Y S,Yang Q E,Ohba H,et al.Violaceae[M].//Wu Z Y,Raven P H.Flora of China,Volume 13.Beijing:Science Press,St.Louis:Missouri Botanical Garden Press,2007:72-111.
7.Tsukaya H.Leaf shape:genetic controls and environmental factors[J].The International Journal of Developmental Biology,2005,49(5-6):547-555.
8.李永华,卢琦,吴波,等.干旱区叶片形态特征与植物响应和适应的关系[J].植物生态学报,2012,36(1):88-98.
Li Y H,Lu Q,Wu B,et al.A review of leaf morphology plasticity linked to plant response and adaptation characteristics in arid ecosystems[J].Chinese Journal of Plant Ecology,2012,36(1):88-98.
9.Hodálová I,Mereda P Jr,Mártonfi P,et al.Morphological characters useful for the delimitation of taxa withinViolasubsect.Viola(Violaceae):a morphometric study from the west Carpathians[J].Folia Geobotanica,2008,43(1):83-117.
10.孙坤,王庆瑞,王漪.国产堇菜属(Viola)植物的数量分类学研究[J].西北师范大学学报:自然科学版,1997,33(3):69-73.
Sun K,Wang Q R,Wang Y.Numerical taxonomic study ofViolain China[J].Journal of Northwest Normal University:Natural Science,1997,33(3):69-73.
11.Beak C B.An introduction to plant structure and development[M].Cambridge:Cambridge University Press,2005.
12.Lang X D,Su J R,Lu S G.CephalotaxustalonensisCheng et Feng ex S.G.Lu et X.D.Lang,a new name of the family Cephalotaxaceae and its taxonomic status[J].Bulletin of Botanical Research,2013,33(1):4-6.
13.Wu J F,Zhang C Y,Chen J Z.Morphological diversity within litchi(LitchichinensisSonn.) based on leaf and branch traits[J].Scientia Horticulturae,2016,207:21-27.
14.Vogel S.Leaves in the lowest and highest winds:temperature,force and shape[J].New Phytologist,2009,183(1):13-26.
15.Hovenden M J,Vander Schoor J K.Nature vs nurture in the leaf morphology of Southern beech,Nothofaguscunninghamii(Nothofagaceae)[J].New Phytologist,2004,161(2):585-594.
16.王林林,赵明富,王赟,等.利用腊叶标本初探椭圆叶花锚(龙胆科)花特征的地理变异[J].植物分类与资源学报,2011,33(5):503-513.
Wang L L,Zhao M F,Wang Y,et al.A preliminary study on geographical variations in floral traits ofHaleniaelliptica(Gentianaceae) based on Herbaria[J].Plant Diversity and Resources,2011,33(5):503-513.
17.Heredia U L D,Valbuena C M,Cordoba M,et al.Variation components in leaf morphology of recruits of two hybridising oaks[Q.petraea(Matt.) Liebl.andQ.pyrenaicaWilld.] at small spatial scale[J].European Journal of Forest Research,2009,128(6):543-554.
18.吴玉环,罗昊,李微,等.牛角藓(Cratoneuronfilicinum(Hedw.) Spruce)叶片形态特征的生态变异及其分类学意义[J].生态学杂志,2007,26(6):882-885.
Wu Y H,Luo H,Li W,et al.Morphological diversity and its taxonomic significance ofCratoneuronfilicinumspruce(Hedw.) Spruce leaf[J].Chinese Journal of Ecology,2007,26(6):882-885.
19.苏雪.兼具开放花和闭锁花繁殖的堇菜属一些类群的遗传结构[D].兰州:西北师范大学,2005.
Su X.The Genetic structure of three pairs of related species ofViolawith chasmogamous and cleistogamous breeding system[D].Lanzhou:Northwest Normal University,2005.
