岷山地区一群川金丝猴冬季的日活动时间分配
2018-04-09郭家强范元英白洢露黎大勇
郭家强,范元英,2,白洢露,胡 杰,黎大勇
(1.西华师范大学 生命科学学院,四川 南充 637009;2.苍溪县国家森林公园管理局,四川 广元 628400)
动物的日活动节律和活动时间分配与其自身能量的收支和代谢有着直接的关系,并且动物会随环境的变化而进行相应的调整。探讨动物对环境的行为适应策略是动物行为生态学研究的重要内容[1]。
动物的日活动节律是指其每天完成生存所必需的各类活动的一个相对稳定的行为程序。诸多因素对其日活动节律产生影响,如白昼温度、日照强度、栖息地质量、形态特征和气候条件等[2]。动物的活动时间分配是指动物根据自身的需求把时间分配到各类必需的日常活动中的过程[3]。生态因素(如食物资源、气候等)、社会因素(如天敌、猴群大小、性别-年龄差异等)等是影响灵长类动物活动时间分配的重要因素[4-6]。
川金丝猴(Rhinopithecusroxellana)属疣猴亚科(Colobinae)动物,为我国特有种、国家Ⅰ级重点保护动物。2003年被IUCN 列为濒危等级(EN),CITES列为附录I物种。目前主要分布于四川、甘肃、陕西、湖北等地,因地理隔离,该物种已分化为三个亚种,即四川亚种(指名亚种)(R.r.roxellana)、秦岭亚种(R.r.qinlingensis)和湖北亚种(R.r.hubeiensis)[7-8]。其中,指名亚种在四川境内呈明显的不连续性分布,主要分布于岷山山系和邛崃山系。
不同地区川金丝猴的活动时间分配有所不同[9-13],这反映了川金丝猴对不同生态环境的行为适应策略。但迄今为止,相关研究多涉及秦岭亚种和湖北亚种,而对于数量最大的指名亚种的研究尚缺乏。同时,冬季又是一年中环境条件最恶劣,猴群生存最艰难的时期,猴群将采取何种适宜的行为策略来应对严酷的自然环境至关重要。为此,我们以四川唐家河国家级自然保护区的一川金丝猴群为研究对象,对其冬季的活动时间分配和活动节律开展了相关的研究,以期了解岷山地区川金丝猴冬季的行为适应策略。
1 研究方法
1.1 研究地点
四川唐家河国家级自然保护区地处岷山山系,摩天岭南麓,位于四川盆地的北缘青川县境内的西北角,与甘肃的白水江国家级自然保护区相邻,地势西北高东南低,属川西高原与四川盆地边缘接壤的高山峡谷区。总面积约400 km2。区内最高峰为大草坪(海拔3 864 m),最低点为石罐子(海拔1 250 m)。这里属于亚热带季风气候,四季分明,雨量充沛。年平均气温为13.7 ℃,年均降雨量为1 100 mm[14]。研究区域内的阔叶林多分布于海拔2 200m以下,主要的树种有糙皮桦(Betulautilis)、锐齿槲栎(Quercusalienavar.acuteserrata)、山胡椒(Linderaglauca),椴树(Tiliatuan)和卵叶钓樟(Linderalimprichtii)等,林下有猫儿刺(Ilexpernyi)、针刺悬钩子(Rubuspungens)、蕊帽忍冬(Lonicerapileata)、糙花箭竹(Fargesiascabrida)等常见灌木;针阔混交林分布于海拔2 200~2 500 m,主要的树种有华山松(Pinusarmandi)、麦吊云杉(Piceabrachytyla)、岷江冷杉(Abiesfaxoniana)、红桦(Betulaalbo-sinensis)和糙皮桦,林下主要为缺苞箭竹(Fargesiadenudata);针叶林分布于海拔2 500~2 800 m,主要的树种为岷江冷杉,林下以缺苞箭竹为优势种。
1.2 研究对象和方法
于2016年11月到2017年2月,以四川唐家河国家级自然保护区石桥河-文县河区域的一群川金丝猴(约138只)为研究对象,该猴群对人的活动已基本习惯化,使得近距离(20~ 50 m) 跟踪和观察猴群成为可能[15]。观察从早上8∶00开始到下午18∶00结束。对猴群进行连续的跟踪观察,每月4~7d,从最早发现猴群开始观察记录,连续跟踪直至猴群进入夜宿地,第二天则提前到夜宿地从8∶00开始继续观察记录。
野外数据收集借助OLYMPUS(10×42)双筒望远镜,在不打扰猴群的适宜距离外进行观察。发现猴群时,采用15 min瞬时扫描取样法[16],收集相关数据。扫描过程中,从猴群的一侧开始,依次记录所见个体在取样时刻所表现的行为类型以避免重复取样,保证样本间的相对独立性。将行为分为:休息(Resting)、移动(Moving)、觅食(Foraging)和其他 (Other)(见文献[17]),且各种行为类型的动作时间须持续5 s以上,同时,在条件允许的情况下,记录行为发生者的性别和年龄。如果在取样时间内,猴群不在视野范围,则停止此次取样,直到下一次取样时间再重新开始扫描。结合野外观察条件,我们将川金丝猴分为4个不同的性别-年龄组:成年雄猴(Adult male)、成年雌猴(Adult female)、青年猴(Juvenile)和婴幼猴(Infant)[18]。
1.3 数据处理
将每次扫描取样视为一个独立的样本,以每次扫描中某种活动的个体数除以本次扫描观察到的个体总数,得出该活动在这一样本中的时间比[2,19-20]。然后把每小时内多次扫描得到的时间比平均,得到每小时各种活动的时间分配,再以每小时的活动时间分配为基本单位求其均值,得到每月的活动时间分配,再求冬季四个月活动时间分配的均值,得到整个冬季川金丝猴的活动时间分配。以每月不同性别年龄组的个体发生各种行为的个体数和取样的个体总数的比值来表示每个月不同性别年龄组个体的活动时间分配,再求出冬季四个月的平均值,得到不同性别年龄组个体的冬季活动时间分配[21]。
利用One-way ANOVA检验来比较日活动节律中前后两个不同时间段活动时间分配的差异,采用Kruskal-Wallis H检验对不同性别年龄组的各类行为时间分配进行比较,若差异显著,则再利用Mann-Whitney U检验来进行两两比较。以上数据的处理和分析借助Microsoft Excel 2003和SPSS 20.0 for Windows软件。
2 结 果
2.1 日活动节律
研究期间,扫描取样2 597次。冬季,川金丝猴群在清晨天亮后离开夜宿地,开始一天的漫游,在上午和下午各有一个明显的取食高峰,在中午有一个明显的休息高峰,并与傍晚天黑前进入夜宿地。One-way ANOVA检验分析表明猴群每一小时内,平均用于取食的时间存在显著性差异(F9,93=2.648,P<0.01);平均用于休息的时间也存在显著性差异(F9,93=2.473,P<0.05);主要表现为,分别在早上的9∶00~10∶00和下午的13∶00~17∶00各有一个取食高峰,中午的12∶00~13∶00为休息高峰(图1)。
2.2 日活动时间分配
冬季由于环境相对比较恶劣,因此川金丝猴把大部分时间都用于获取食物,平均用于取食的时间最多,占49.92%;其次是移动,移动是为寻找更多的食物,平均用于移动的时间占31.29%;平均用于休息的时间较少,占16.87%,用于理毛等其他行为的时间最少,仅占1.92%。
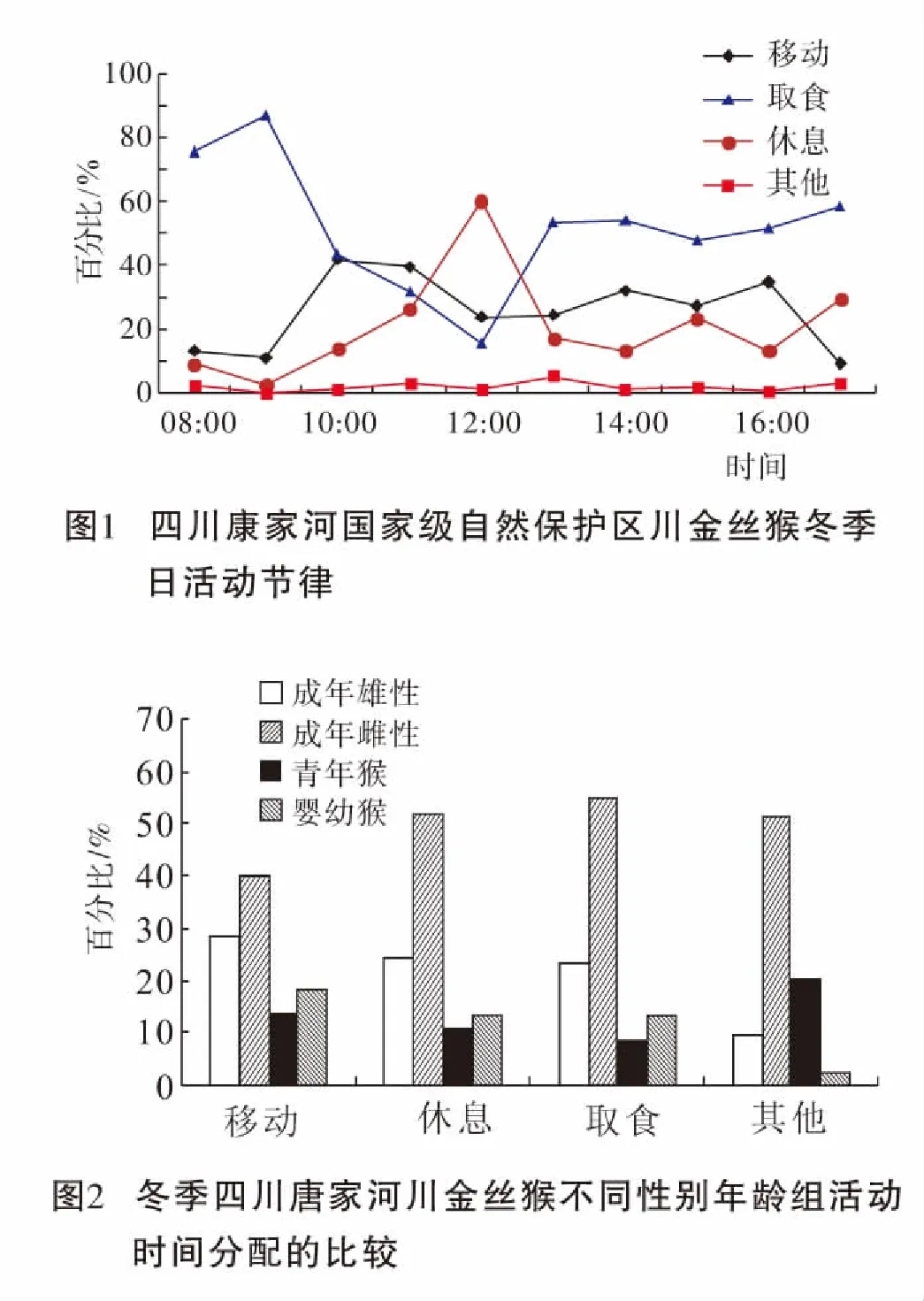
2.3 不同性别年龄组个体的冬季活动时间分配
经Kruskal-Wallis H检验各性别-年龄组在各种行为类型的时间分配上均存在显著差异(P<0.05)(图2)。进一步分析表明,成年雌猴个体花费54.92%的时间用于觅食,显著高于成年雄猴的23.22%(Mann-Whitney U检验,Z=-2.882,P=0.002),青年猴的8.66%(Z=-2.882,P=0.002)和婴幼猴的13.20%(Z=-2.882,P=0.002);成年雄猴用于觅食的时间也显著多于青年猴(Z=-2.722,P=0.004)。同时,成年雌猴用于休息的时间也较多,达51.84%,显著高于成年雄猴的24.5%(Z=-2.402,P=0.015)、青年猴的10.88%(Z=-2.882,P=0.002)和婴幼猴的12.78%(Z=-2.807,P=0.002)。在移动行为上,成年雄猴的移动时间显著较青年猴(Z=-2.882,P=0.002)和婴幼猴多(Z=-2.326,P=0.015),而与成年雌猴的移动时间分配相似(Z=-1.761,P=0.093);青年猴与婴幼猴的移动时间分配无显著差异(Z=-1.363,P=0.180)。此外,在其他行为中,仅成年雌性与婴幼猴的时间分配有显著差异(Z=-2.478,P=0.015)。
3 讨 论
3.1 日活动节律
在本研究中,唐家河川金丝猴群冬季的日活动节律表现为上午和下午各有一个觅食高峰,中午有一个休息高峰。相似的活动节律在众多食叶的疣猴类中均有发现,例如川金丝猴(Rhinopithecusroxellana)[9-10]、黔金丝猴(Rhinopithecusbrelichi)[22]、滇金丝猴(Rhinopithecusbieti)[23-24]、黑叶猴(Trachypithecusfranciosi)[25]及白头叶猴(Trachypitheucsleucocephalus)[26-27]等。这些食叶性疣猴类之所以有这样的日活动节律可能代表了疣猴类对温度的一种适应:冬季气温较低,而中午正好是阳光直射,日间温度最高的时候,此时猴群会选择一个背风向阳的地方进行午休,使阳光能够直射到躯体以维持体温[2]。另外,中午的休息也有利于叶食性的疣猴类消化上午取食高峰摄取的大量纤维素食物[17]。
3.2 日活动时间分配
动物群体如何利用时间反映了该种动物的生存策略[28]。当食物匮乏时,灵长类会采取三种不同的调节措施来应对恶劣的环境:(1)增加维持基本生存的活动时间,即增加觅食和移动时间;(2)仅增加觅食的时间;(3)增加休息时间或减少高能量消耗活动的时间[29-30]。我们的研究表明:冬季唐家河川金丝猴采取了第一种类型的生存策略,即花费更多的时间用于觅食和移动(占81.21%)。
典型的疣猴类动物通常会花费大量的时间(≥40%)来休息[31-32],而金丝猴尚属例外[9,11-12],我们的研究也再次表明川金丝猴用于休息的时间(16.87%)远不及其他的疣猴类,但具体的生物学机制目前仍不清楚。
与其他地区的研究结果相比较(表1),我们发现唐家河川金丝猴冬季花费的觅食时间除了仅低于秦岭玉皇庙的东梁群外[12],高于秦岭的西梁群[9]及其他地区川金丝猴的觅食时间分配[10-11]。其中一个重要的原因或许是由于不同地区川金丝猴冬季食物资源的可获得性差异造成的。冬季其他类型的食物较为匮乏,地衣类植物成为川金丝猴越冬的主要食物,例如,在四川白河地区,川金丝猴在冬季对地衣类的摄取量高达51%[33],秦岭地区川金丝猴冬季摄取地衣类植物的量占41%[34],神农架地区川金丝猴冬季食物中松萝类植物占43.28%[18]。但在唐家河,川金丝猴冬季主要的食物为树(枝)皮,而地衣类植物相对较少,因此这里的川金丝猴将会花费更多的时间来找寻和取食。
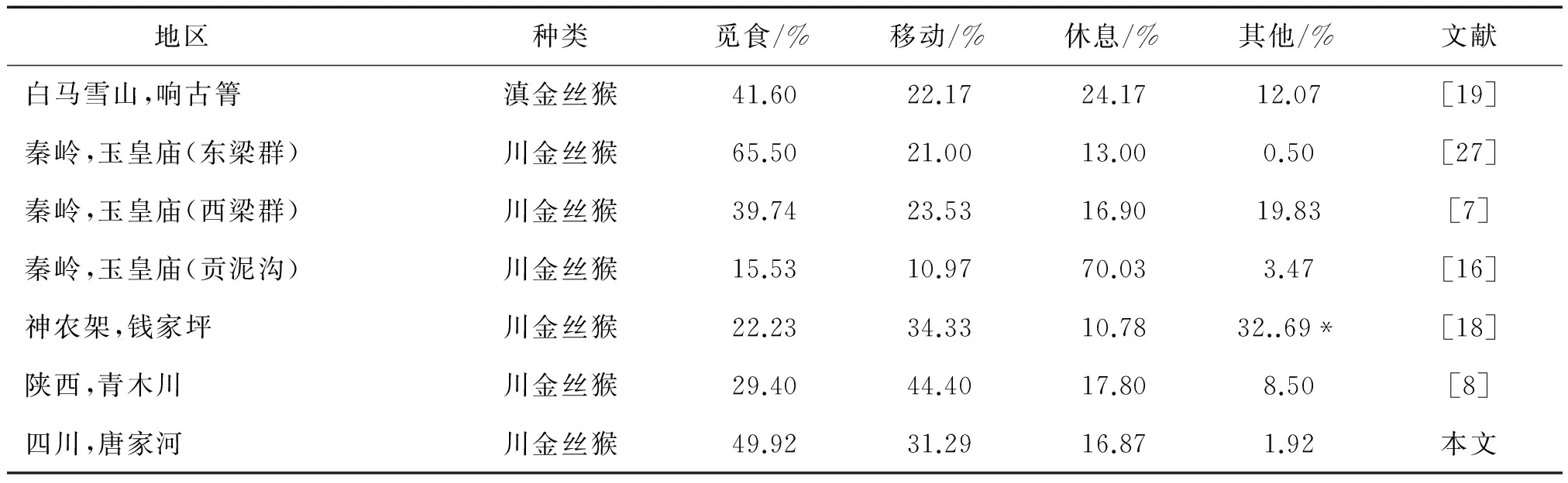
表1 不同地区几种金丝猴冬季活动时间分配
注:*包括社会行为和其他行为。
3.3 不同性别-年龄组个体的活动时间分配
性别、年龄也是影响灵长类活动时间分配的一个重要因素[11,20]。我们的研究表明:冬季川金丝猴成年雌猴平均用于觅食和休息的时间均高于群内的其他成员。这或许是由于成年雌猴的繁殖需求所决定的[35]。在冬季,由于成年雌猴要么正处于孕期,要么还担负着照顾婴幼猴的重任,因此成年雌猴往往会花费更多地时间来觅食,以满足其对于营养和能量的需求,同时增加休息时间,还能降低能量的消耗,这无疑是一种很好的行为适应策略。成年雄猴则不同,它们仅需要满足当前的能量需求,因此,它们的觅食时间相对较短。成年雄猴和成年雌猴平均用于移动的时间高于青年猴和婴幼猴,这可能是因为成年雄猴需要频繁地巡视和移行[36],而成年雌猴需要移动更远的距离来搜寻足够多的食物,婴幼猴则经常依附在母亲的怀里,很少单独活动,因此移动的时间较少。在其他行为方面,成年雌猴与婴幼猴存在显著性差异,成年雌性在理毛等其他行为上花费的时间更多,这是因为成年雌猴更注重个体间的友谊和交往[35],往往会花费较多的时间给群内其他成员理毛,以获得更多的理毛回报,维系与家庭成员之间的关系[37]。婴幼猴则由于行为发育尚不完善,其社会行为相对较少。
致谢:本研究在野外数据收集过程中,唐家河国家级自然保护区的领导和工作人员给予了支持和帮助,在此表示谢意!
参考文献:
[1]HALLE S,STENSETH N C.Activity Patterns Small Mammals,Ecological Approach[M].New York:Springer-Verlag Berlin Heidelberge,2000.
[2]CLUTTON-BROCK T H,HARVEY P H.Species Differences in Feeding and Ranging Behavior in Primates[M]//Clutton-Brock T H Ed.Primate Ecology:Studies of Feeding And Ranging Behaviour in Lemurs,Monkeys and Apes[M].London:Academic Press,1977.
[3]ISBELL L A,YOUNG T P.Social and Ecological Influences on Activity Budgets of Vervet Monkeys and Their Implications For Group Living[J].Behavioral Ecology and Sociobiology,1993,32(6):377-385.
[4]SMITH C C.Feeding Behaviour and Social Organization in Howling Monkeys.in:Clutton-BRock T H ed.P Rimate Ecology:Studies of Feeding and Ranging Behaviour in Lemurs,Monkeys and Apes[M].London:Academic Press,1977.
[5]MARSH C W.Time budget of Tana River Red Colobus[J].Folia primatol,1981,35(1):30-50.
[6]朱荣华,黎大勇,姚刚,等.圈养川金丝猴活动时间分配研究[J].西华师范大学学报(自然科学版),2016,37(3):253-255.
[7]邓其祥,胡锦矗,余志伟.金丝猴的生态生物学特性及其分布[J].南充师院学报(自然科学版),1981,2(3):75-85.
[8]全国强,谢家骅.金丝猴的研究[M].上海:上海科技教育出版社,2002.
[9]吕九全,李保国.秦岭川金丝猴的昼间活动时间分配[J].兽类学报,2006,26(1):26-32.
[10]许鹏,李言阔,缪涛,等.陕西青木川自然保护区川金丝猴的昼间时间分配和活动节律[J].江西科学,2015,33(3):324-329.
[11]LI Y M.Activity Budgets in A Group of Sichuan Snub-Nosed Monkeys in Shennongjia Nature Reserve,China[J].Current Zoology,2009,55(3):173-179.
[12]GUO S T,LI B G.WATANABE K.Diet And Activity Budget of Rhinopithecus Roxellan in The Qinling Mountains,China[J].Primates,2007,48(4):268-276.
[13]LIU X C,STANFORD C B,LI Y M.Effect of Group Size on Time Budgets of Sichuan Snub-Nosed Monkeys (Rhinopithecusroxellana) in Shennongjia National Nature Reserve,China[J].International Journal of Primatology,2013,34(2):349-360.
[14]胡锦矗.四川唐家河自然保护区综合科学考察报告[R].成都:四川科学技术出版社,2005.
[15]范元英,黎大勇,黄小富,等.岷山山系川金丝猴群的社会结构——以唐家河国家级自然保护区川金丝猴为例[J].四川动物,2015,34(6).832-836.
[16]ALTMANN J.Observational Study of Behavior:Sampling Methods[J].Behaviour,1974,49(3):227-267.
[17]黎大勇,任宝平,胡杰,等.白马雪山自然保护区响古箐滇金丝猴活动时间分配[J].兽类学报,2013,33(3):223-231.
[18]LI Y M.Seasonal Variation of Diet and Food Availability in A Group of Sichuan Snub-Nosed Monkeys in Shennongjia Nature Reserve,China [J].Am.J.Primatol,2006,68(3):217-233.
[19]HARRISON M J S.Time Budget of The Green Monkeys,Cercopithecus Sabaecus:Some Optimal Strategies[J].International Journal of Primatology,1985,6(4):351-376.
[20]AGETSUMA N,NAKAGAWA N.Effects of Habitat Differences on Feeding Behaviors of Japanese Monkeys:Comparison Between Yakushima and Kinkazan[J].Primates,1998,39(3):275-289.
[21]唐创斌,蒋建波,黄乘明,等.环境和社会因素对喀斯特石山猕猴日活动节律和活动时间分配的影响[J].兽类学报,2017,37(2):131-138.
[22]杨业勤,雷孝平,杨传东.黔金丝猴的野外生态[M].贵阳:贵州科技出版社,2002.
[23]DING W,ZHAO Q K.Rhinopithecus Bieti at Tacheng,Yunnan:Diet and Daytime Activities[J].Int J Primatol,2004,25(3):583-598.
[24]李沂韦,黎大勇,任宝平,等.白马雪山自然保护区超大滇金丝猴群的日活动节律[J].西华师范大学学报(自然科学版),2015,36(3):221-227.
[25]黄乘明,周岐海,李友邦,等.广西扶绥黑叶猴活动节律和日活动时间分配[J].兽类学报,2006,26(4):380-386.
[26]周岐海,黄恒连,唐小平,等. 白头叶猴日活动时间分配及其季节性变化[J].兽类学报,2010,30(4):449-455.
[27]黄乘明.中国白头叶猴[M].桂林:广西师范大学出版社,2002.
[28]POST W,BAULU J.Time Budgets of Macaca Mulatta[J].Primates,1978,19(1):125-140.
[29]WATANUKI Y,NAKAYAMA Y.Age Difference in Activity Patterns of Japanese Monkeys:Effects of Temperature,Snow,and Diet[J].Primates,1993,34(4):419-430.
[30]SCHOENER T W.Theory of Feeding Strategies[J].Annual Review of Ecological Systems,19712(1):369-404.
[31]HUANG C M.LI Y B.ZHOU Q H.Activity Patterns and Their Occurrence in Day Range of Francois Langur (Trachypithecus Francoisi) in Fusui Reserve.Guangxi[J].Journal of Guangxi Normal University,2003,21(4):78-81.
[32]STANFORD C B.The Capped Langur in Bangladesh:Behavioral Ecology and Reproductive Tactics[J].Contributions To Primatology,1991,26(1):1-179.
[33]KIRKPATRICK R C,GU H J,ZHOU X P.A Preliminary Report on Sichuan Snub-Nosed Monkey (RhinopithecusRoxellana) at Baihe Nature Reserve[J].Folia Primatol,1999,70(2):117-120.
[34]赵海涛,党高弟,王程亮,等.秦岭南坡川金丝猴的食物组成及季节性变化[J].兽类学报,2015,5(2):130-137.
[35]李宏群,张育辉,李保国.秦岭川金丝猴秋冬季节活动时间分配的初步研究[J].陕西师范大学学报(自然科学版),2004,32(2):86-89.
[36]李兆元.白头叶猴(Presbytisleucocephalus)活动时间分配[J].兽类学报,1992,12(1):7-13.
[37]朱紫瑞.秦岭川金丝猴(Rhinopithecusroxellana)成年雌性相互理毛的时间匹配性以及理毛策略的研究[D].西安:西北大学,2008.
