棉花种子老化过程中生理生化指标变化比较研究
2018-04-09,,,,
, , , ,
(1.辽宁省经济作物研究所, 辽宁 辽阳 111000; 2.沈阳农业大学农学院, 沈阳 110161)
种子作为农业生产的关键材料,通常具有一个生命体、一个营养供应体和一个保护结构[1],生命体的活性、自身营养供应、保护结构的完整性,三者共同决定了种子的发芽、出苗和成苗能力。1977年,国际种子检验协会(ISTA)将种子在发芽和出苗期间的活性强度及特征的综合表现定义为种子活力[2-3]。
种子生理成熟后便进入贮藏或休眠阶段,该阶段种子会发生老化或劣变,发芽率和活力等随之降低。Gove等[4]将种子老化和劣变定义为:种子的生活力、品质及其性能从一个较高水平下降至较低水平的不可逆的变化过程。种子的老化或劣变包含一系列的变化。从外表看,包括种皮变色,光泽度降低,油脂种子“走油”等[5]。从内部结构看,主要表现为细胞显微结构的变化,突出表现为膜系统受损,膜结构和功能发生转变[5-6],渗漏加速。从生理方面看,丙二醛(MDA)、抗坏血酸(AsA )、过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)等物质含量或酶活性升高或较低,呼吸作用与ATP含量变化,核酸与蛋白质等大分子物质合成能力明显下降,甚至染色体结构发生畸变、DNA断裂等[4]。从萌发情况上看,发芽率、发芽指数和整齐率下降,生长缓慢[6],抗逆性下降,弱苗、白化苗、畸形苗增多,严重时会导致生活力丧失[7],从而造成难以估计的经济损失[5,8]。
种子劣变受多方面因素影响,包括内部因素和外部因素。种子劣变的机理目前认为与膜损伤、大分子物质变化、有毒物质积累和内源激素不平衡有关[9]。影响种子老化或劣变的因素以及种子老化过程中的生理生化变化是前人研究所关注的问题,研究范围包括玉米[10]、大豆[11]、小麦[12]、花生[13]、大麦[14]等主要农作物,也包括苜蓿[15,25]、罗布麻[16,23]、中草药[17]等,但迄今为止,尚无棉花种子此类研究的报道。
棉花作为我国主要的农作物,其产业发展关乎国计民生。棉花种子分2类:一类为毛籽,即经过初步轧花后,表面附有短绒的种子;一类为光籽,即脱去短绒后的光滑种子。经验认为,毛籽耐贮藏,耐播种期的不利条件(如低温等);而光籽出苗快,易于操作,尤其适于机械播种。棉籽含油量较高,为探讨光籽和毛籽的老化过程差异以及对种子活力的影响,2016年进行了棉花种子人工老化试验,以期为棉花种子的贮藏和利用提供理论参考。
1 材料与方法
1.1 试材及处理
以2015年收获的棉花品种辽棉23号和辽棉31的种子为试材。将所选试材分成2部分,一部分不脱绒为毛籽,另一部分利用硫酸脱绒成光籽。选取适量的饱满种子,分别装在网袋中,于恒温培养箱内进行老化处理。条件设置为温度T=40 ℃、湿度R=100%,避光,处理时间分别为2,4,6,8 d,以未经老化处理的种子为对照(ck)。处理结束后,将样本置于室温下1~2 d,恢复正常温湿度。为避免短绒对试验结果造成影响,将毛籽用硫酸脱绒成光籽。
1.2 种子生理指标的测定
将各处理种子在室温条件下用蒸馏水浸泡24 h,种子激活后,即可进行各项指标的测定。
采用DDBJ-350型电导率仪测定并计算种子外渗液电导率值,采用TBA比色法测定种子丙二醛(MDA)含量,采用愈创木酚法测定种子过氧化物酶(POD)含量,采用紫外吸收法测定种子过氧化氢酶(CAT)和超氧化物歧化酶(SOD)含量。具体操作方法参考李玲等编写的《植物生理学模块实验技术》。因为本试验试材为种子材料,样品含水量较少,因此在研磨时,将研磨缓冲液加倍。
所有指标的测定均重复3次,最终结果取平均值。
1.3 种子发芽率测定
参照标准发芽试验,将经过老化处理的光籽及经过脱绒的毛籽用5%硼酸消毒5 min,选取50粒,放在垫好滤纸的培养皿中,添加足量蒸馏水;将培养皿置于人工气候箱内,温度设置为28 ℃,每天光照12 h。第4天和第7天检查发芽情况并计算发芽率。
2 结果与分析
2.1 种子渗出液电导率比较分析
由图1可见,随着种子老化程度加深,种子渗出液电导率值也在提高。说明种子的细胞膜结构在老化过程中被破坏,渗出量增加,这一点与前人的研究结果相符合。
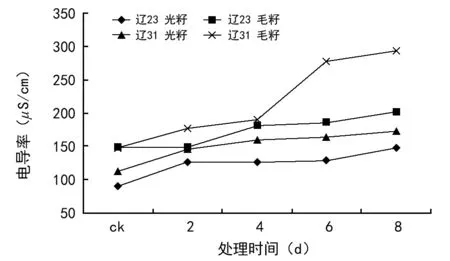
图1 种子渗出液电导率
辽棉23号种子渗出液电导率值低于辽棉31种子,说明其细胞膜受破坏程度要轻一些。以往的经验认为,毛籽由于有短绒保护,比光籽更耐贮存,但本试验中毛籽的渗出液电导率却高于光籽,这个现象有待进一步探讨。
2.2 种子丙二醛(MDA)含量比较分析
由图2可见,种子丙二醛(MDA)含量的变化表现比较复杂,曲线呈波动状态。
光籽的MDA含量呈上升—下降—再上升的趋势,而且辽棉31的表现要平稳一些,辽棉23号在老化4 d时有一个明显的飞跃,之后再进入平稳状态。毛籽的MDA含量首先明显下降,之后辽棉23号持续下降,到老化8 d时有所上升;而辽棉31号呈上升—下降—再上升的波动状态。辽棉23号光籽的MDA含量要高于辽棉31,但毛籽的MDA含量由于波动较大,不宜进行比较。
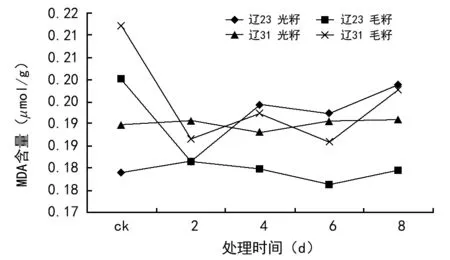
图2 种子丙二醛(MDA)含量
2.3 种子过氧化物酶(POD)活性比较分析
由图3可见,种子过氧化物酶(POD)活性的变化表现比较复杂,但是除辽棉31光籽呈现较强烈波动状态外,总体下降—上升—再下降的趋势。
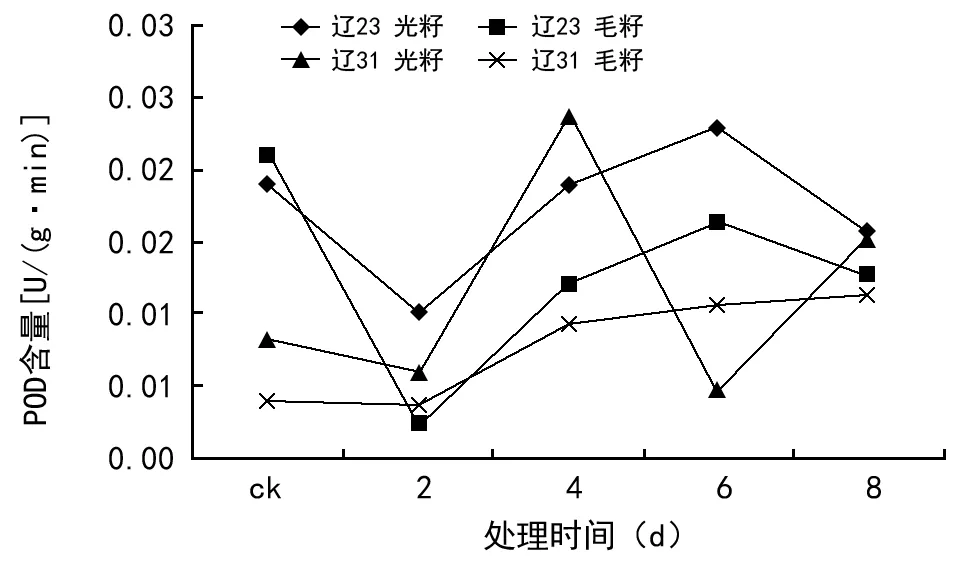
图3 种子过氧化物酶(POD)活性
辽棉23号无论是光籽还是毛籽,其种子内POD活性都呈先下降—后上升—再下降的趋势,光籽的波动较大。辽棉31号光籽的POD活性总体曾上升趋势,但在老化4 d时出现一个峰值;而毛籽的POD活性呈现平稳上升趋势。
2.4 种子过氧化氢酶(CAT)活性比较分析
由于具体原因,本试验只得到了辽棉23号种子的过氧化氢酶(CAT)活性值。由图4可见,种子CAT活性表现为先升高、后降低的趋势。毛籽的CAT活性明显高于光籽。可以认为,毛籽有短绒保护,受老化影响较小,所以激活后,其CAT活性较高。
2.5 种子超氧化物歧化酶(SOD)活性比较分析
由于具体原因,本试验只得到了辽棉31种子超氧化物歧化酶(SOD)活性值。由图5可见,光籽的SOD活性表现为先明显升高、后迅速降低、到处理6 d时趋于平稳的趋势。毛籽的SOD活性表现轻微下降、再缓慢上升、到老化6 d时明显下降的趋势。光籽的SOD活性高于毛籽,这一点与预期结果相悖,需要进一步试验加以印证。
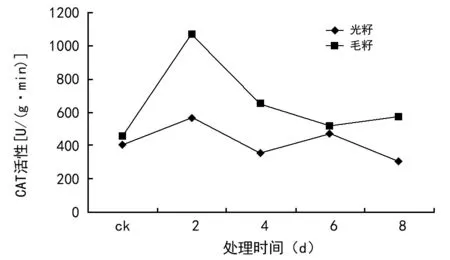
图4 辽棉23号种子过氧化氢酶(CAT)活性

图5 辽棉31种子超氧化物歧化酶(SOD)活性
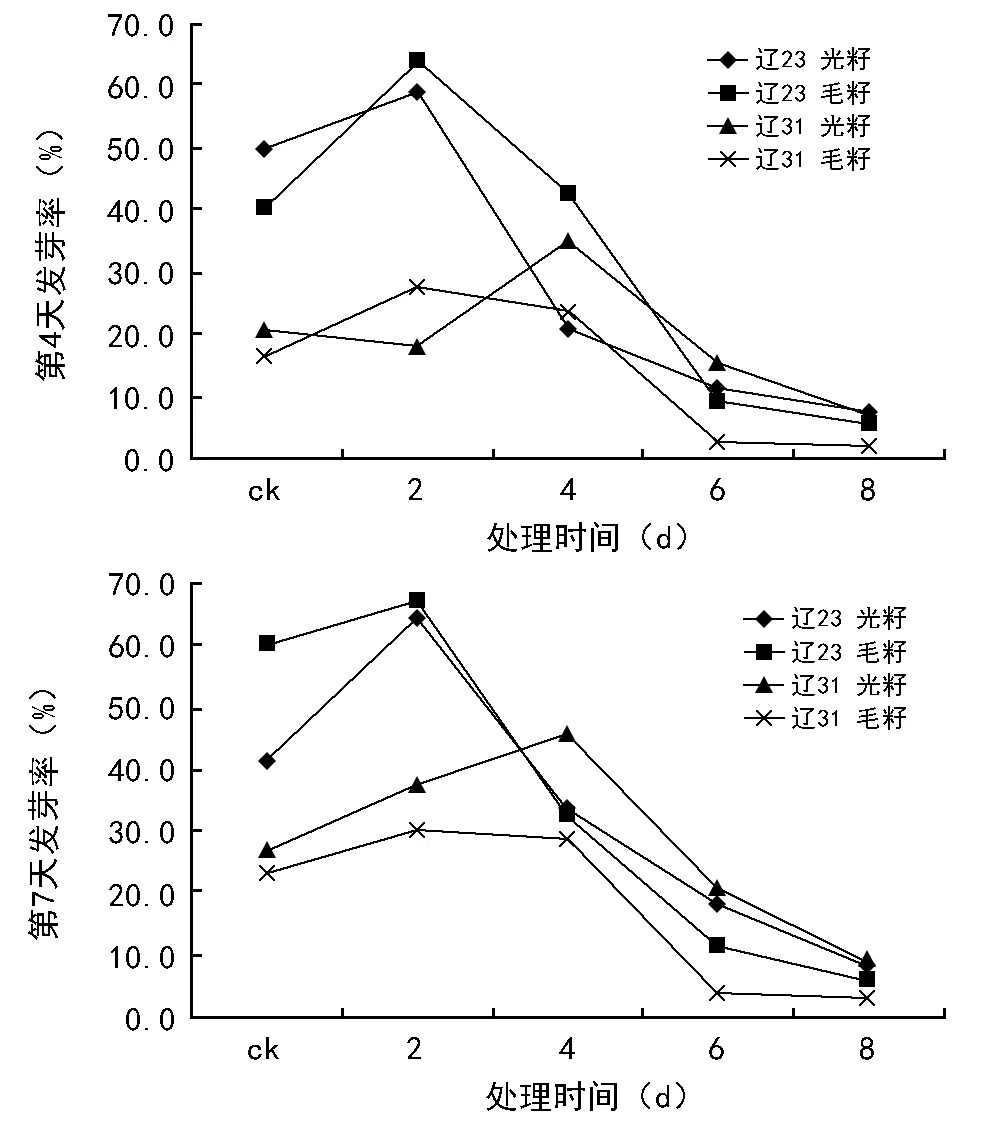
图6 种子发芽率(第4天,第7天)
2.6 种子发芽率比较分析
由图6可见,发芽第7天时的发芽率稍高于第4天时的发芽率。对于经过老化处理6 d和老化处理8 d的种子,光籽的发芽率低于20%,毛籽的发芽率低于10%,并且第4天和第7天时差别不大,已经基本丧失发芽能力。比较来说,辽棉23号基本表现为毛籽的发芽率高于光籽的发芽率,而辽棉31在发芽第7天时明显表现光籽的发芽率高于毛籽的发芽率,发芽第4天时这个趋势不明显。
2个品种的发芽率都表现出先升高后降低的趋势。辽棉23号表现活跃一些,其对照处理发芽率较高,老化处理2 d的发芽率最高,老化处理4 d的发芽率急剧下降,到老化处理6 d和老化处理8 d,其发芽能力已经基本丧失。辽棉31的表现要迟钝一些,其发芽率的峰值出现在老化处理4 d,总体来说,发芽率低于辽棉23。
3 结论与讨论
3.1 种子老化过程中的一系列生理反应
关于种子老化过程中的生理生化指标变化,前人进行了大量的研究。周国栋等关于芒麦种子的研究表明,老化后种子细胞膜的透性增加,浸出液电导率也随之增加,有毒害作用的丙二醛(MDA)含量增加[18]。李雪峰等关于辣椒种子的研究得出同样的结果,即随着种子老化程度加深,种子浸出液电导率升高,丙二醛(MDA)含量增加[19]。同样的结论也出现在麻黄[20]、实葶葱[21]、结球白菜[22]等种子的研究上。在酶活性方面,刘霞等关于蓝萼香茶菜种子的研究表明,种子老化会导致抗氧化系统能力降低,过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)的活性都会降低[23]。张永娟等在罗布麻种子老化机理研究的过程中也发现,随着种子老化程度的加深,过氧化氢酶(CAT)和脱氢酶活性随之降低,并且这2个比较敏感的指标可以作为测定种子活力的生化指标来检验种子的劣变程度[24]。
本试验中,随着种子老化程度的加深,种子渗出液电导率呈升高趋势,这一点与前人的研究结果相符;丙二醛(MAD)含量方面,除毛籽老化处理2 d的有明显下降外,其余处理呈缓慢上升趋势;过氧化物酶(POD)活性也呈波动状态,但波动幅度较小,基本呈平稳上升趋势;过氧化氢酶(CAT)活性在老化处理2 d时有一个峰值,之后微幅波动,总体呈下降趋势;超氧化物歧化酶(SOD)活性与其类似,只是毛籽的峰值出现在老化处理6 d。总体来说,该试验在种子渗出液电导率和丙二醛(MAD)含量2个指标上所得到的结论与前人的研究结果基本相符;在酶活性方面,过氧化氢酶(CAT)活性和超氧化物歧化酶(SOD)活性所得结论基本相符,但过氧化物酶(POD)活性变化趋势还有待进一步印证。
3.2 不同棉花品种之间、光籽与毛籽之间对老化处理的反应
本试验结果表明,2个品种对老化处理的反应有明显的区别。总体来说,辽棉31的耐贮藏能力相对较差,因为其渗出液电导率、丙二醛(MAD)含量都较高,而过氧化物酶(POD)活性却较低,而且,随着老化程度加深,差距也会越来越大。光籽和毛籽对老化处理的反应也有区别,渗出液电导率、过氧化氢酶(CAT)活性2个指标,毛籽要高于光籽;而过氧化物酶(POD)活性、超氧化物歧化酶(SOD)活性2个指标,光籽要高于毛籽;丙二醛(MAD)含量本试验中趋势不明显。由此可见,棉花种子贮藏过程中,不同品种、不同处理的种子要分别对待,以保证种子的实际应用价值。
3.3 种子渗出液电导率变化情况与预期相反
有研究表明,随着老化程度的加深,细胞膜质结构受损程度会加深,种子渗出液电导率也会随之升高[18-22],这一点在本试验中得到了证实。但是,以往的经验认为,毛籽由于有短绒保护,比光籽更耐贮存。但本试验中毛籽的渗出液电导率高于光籽,说明其受破坏程度较重,这个现象与经验相悖。
本试验中,毛籽老化后要经过硫酸脱绒才能进行试验,而光籽是约3个月前脱绒处理的。硫酸脱绒的过程中,种皮会遭到破坏(包括机械损伤和化学损伤),但种子作为活体,有一定的自愈功能,通过油脂外渗、细胞修复等,种皮的损伤会得到一部分修复,从而抗老化程度有所恢复。推论认为这是本试验中毛籽的渗出液电导率高于光籽的原因,还需要进一步证实,尤其是种子对于损伤的修复机理,还有待研究。
3.4 老化处理对种子发芽率的影响
本试验中,无论是毛籽还是光籽、第4天还是第7天,其发芽率都表现为先上升、后下降的趋势,即:与对照相比,老化处理2 d的种子发芽率最高,其后下降。老化处理6 d时,种子的发芽率已经在10%以下,老化处理8 d的种子发芽率更低,根据通常概念,此时的种子已经没有应用价值。
温汤浸种是生产中常用的催芽方法,是种子热击处理的方法之一。该方法一般是在40~60 ℃的条件下将种子处理30 min,从而增强种皮的通透性,使氧气顺利进入种子内部,增强种子的代谢活动,为种子萌发提供充足的物质和能量。王旭明等将番茄种子在40 ℃条件下处理20 min,结果发现,种子发芽率得到显著提高[25]。孔令琪等在研究老化对苜蓿硬实种子的影响时也发现,老化处理后种子的发芽率、浸出液电导率较处理前均有所提高[26]。刘旭欢等研究老化处理对春小麦种子活力的影响也认为,从老化4 d开始,随着老化时间增长,发芽相关指标均呈明显下降趋势,认为种子在短时间内受到高温高湿胁迫伤害较轻,老化的时间越长,种子受到的伤害越深,劣变越快[12]。
本试验结果表明,老化处理2 d时,种子在较高温度和湿度条件下被激活,因此发芽率较高。随着老化处理时间延长,种子内含物发生变化,有毒有害物质积累增加,影响了种子活力,降低了发芽率。这一点与前人的研究结论相符。但是,关于种子内含物及生理生化指标的变化与种子发芽率之间的关系,本试验并没有得到可供参考的结论,还需要继续深入研究。
总之,通过本试验得出结论,不同品种、不同处理的棉花种子抗老化能力有较明显的差别,因此在加工贮藏过程中应该区别对待。另一方面,老化处理2 d的种子发芽率提高,证明适当的高温高湿处理有利于种子的激活和萌发。
参考文献:
[1]颜启传,李国忠,伍斌.种子学[M].北京:中国农业出版社,2001(6):1-2.
[2]孙群,王建华,孙宝启.种子活力的生理和遗传机理研究进展[J].中国农业科学,2007,40(1):48-53.
[3]王显国,韩建国,陈志红.新麦草种子成熟过程中活力变化的研究[J].草地学报,2002,8(4):306-311.
[4]闰慧芳,夏方山,毛培胜.种子老化及活力修复研究进展[J].中国农学通报,2014,30(3):20-26.
[5]王维婷,单成钢,朱京斌,等.中药种子成熟度、劣变机制及贮藏方法研究进展[J].种子科技,2010(2):26-30.
[6]李培英,孙宗玖,阿不来提,等.常温贮藏下新农1号狗牙根种子劣变的研究[J].草业科学,2007,10(10):63-67.
[7]沈玉忠.种子劣变因素的分析[J].教育教学论坛,2011(20):243-244.
[8]杨亚平.水稻种子劣变的生理生化机制及其相关蛋白研究[M].长沙:湖南师范大学出版社,2008:1-10.
[9]黄情,李云霞,魏先杰,等.种子劣变与修复[J].种子,2013,32(4):40-44.
[10]李春雷,马世骏,彭滨,等.人工老化对玉米种子活力指标、内含物质含量及生理指标的影响[J].吉林农业大学学报2014,36(5):505-509,514.
[11]高华伟,满强,潘晶,等.大豆人工老化与自然老化的种子活力差异研究[J].种子,2015,34(1):14-18.
[12]刘旭欢,加得拉·努尔兰,童婷,等.不同老化时间和收获期对春小麦种子活力的影响[J].新疆农业科学,2014,51(8):1 394-1 399.
[13]沈一,刘永惠,陈志德.人工老化对花生种子营养成分的影响[J].江苏农业科学,2013,41(10):79-81.
[14]廖乐,齐军仓,吕小兰,等.人工老化对大麦种子活力的影响[J].种子,2014,33(8):71-74.
[15]李春杰,王彦荣,朱廷恒,等.紫花苜蓿种子对逆境贮藏条件的反应[J].应用生态学报,2002,13(8):957-961.
[16]刘志华,王妮,陈立宇,等.罗布麻种子自然状态老化过程中生理生化指标测定与分析[J].种子,2015,34(3):43-46.
[17]赵冬,李明.白木香种子贮藏过程的生理生化指标变化[J].湖北农业科学,2013,52(22):5 556-5 560.
[18]周国栋.种子老化对老芒麦种质生理特性及遗传完整性变化的影响[D].北京:中国农业科学院,2012.
[19]李雪峰,邹学校,刘志敏.辣椒种子人工老化及劣变的生理生化变化[J].湖南农业大学学报(自然科学版),2005,31(3):265-268.
[20]李胜,张真,杨广兴,等.不同处理对麻黄种子生理生化特性的影响[J].草业科学,2007,24(1):59-64.
[21]帕提曼·阿布都热合曼,林辰壹,艾克怕尔·吾买尔.实葶葱种子老化过程生理生化变化的研究[J].新疆农业大学学报,2011,34(2):125-128.
[22]薄丽萍,吴震,蒋芳玲,等.不结球白菜种子活力及抗氧化特性在人工老化过程中的变化[J].西北植物学报,2011,31(4):724-730.
[23]张永娟,韩蕊,原烨,等.罗布麻种子老化过程中的生理生化特性[J].草业科学,2011,28(12):2 130-2 135.
[24]刘霞,刘菲.蓝萼香茶菜种子人工老化过程中生理生化特性的研究[J].种子,2012,31(4):27-30.
[25]王旭明,洪法水,靳英.热击处理对番茄种子萌发及幼苗生长的影响[J].长江蔬菜,1998(2):28-29.
[26]孔令琪,王俊峰,刘国庆,等.苜蓿硬实种子对人工老化时间处理的生理响应[J].草地学报,2011,19(1):81-85.
