不同动物模型对高山美利奴羊早期生长性状遗传参数估计的比较
2018-04-03张剑搏袁超岳耀敬郭健牛春娥王喜军王丽娟吕会芹杨博辉
张剑搏,袁超,岳耀敬,郭健,牛春娥,王喜军,王丽娟,吕会芹,杨博辉
(1中国农业科学院兰州畜牧与兽药研究所/中国农业科学院羊育种工程技术研究中心,兰州 730050;2甘肃省绵羊繁育技术推广站,甘肃肃南 734031)
0 引言
【研究意义】家畜育种的目的是为了不断改进畜群遗传素质,提高生产性能,培育新的品种或品系,而育种方案的科学制定必须以准确、可靠的遗传参数估计为前提条件。为了获得更加准确的估计遗传参数,需要建立一个合理的分析模型,该模型不仅要在生产实践中具有可操作性,还要能准确反映出遗传和环境因素对性状的影响[1]。在数量性状的遗传分析中,育种科研人员常用的分析模型有公畜模型、公畜母畜模型、外祖父模型以及动物模型。无论从遗传学还是统计学的角度看,随着计算机技术和计算方法的日益完善,考虑了所有血缘关系的动物模型都要优于其他分析模型,由于其他模型的使用都有着极其严格的条件限制,不利于在育种实践操作而被逐渐淘汰[2]。同时,随着生物统计理论和方法在家畜育种中的应用,遗传参数估计的理论方法也得到了很大进步,特别是线性混合模型理论和方法与家畜育种的结合,使得遗传参数的估计更加准确。目前,最为流行的遗传参数估计方法为贝叶斯估计法和REML法,其中REML法根据迭代方法的不同分为EMREMK、DFREML和AIREML算法,而AIREML算法是将Fisher’s scoring方法和Newton-Raphson算法有机结合起来的一种数学算法,它可以大大节省计算时间和存储空间[3]。高山美利奴羊是以澳洲美利奴羊为父本、甘肃高山细毛羊为母本,历经 20年育成的中国第一个拥有自主知识产权的高山型美利奴羊新品种。该品种常年生活在海拔 2 400—4 070 m的青藏高原高山寒旱草原严酷生态自然条件下,具有抗逆性强、生产性能高、羊毛品质优等特点[4-5]。由于生存条件的特殊性,其生长性状及遗传特性受高原寒旱环境的影响程度均与其他细毛羊品种不同。因此,利用不同动物模型估计高山美利奴羊的早期生长性状的遗传参数,为高山美利奴羊早期阶段的选育提高提供理论依据;同时,对于研究该品种早期生长性状的遗传特性具有重要意义。【前人研究进展】在 1986年后,美国国家绵羊改良计划(NSIP)选择塔基绵羊进行遗传改良,在很大程度上改进了其遗传进展和生产性能[6]。HANFORD 等[7]估计了塔基绵羊的初生重遗传力为 0.25,断奶重遗传力为 0.22。CLOETE等[8]估计美利奴羔羊的初生重遗传力为 0.16—0.17,母体效应遗传力为 0.28—0.37。断奶重遗传力为0.16,断奶毛长遗传力为0.27。DI等[9]利用多性状动物模型估计了超细型中国美利奴羊的初生重、断奶重和断奶毛长的遗传力分别是 0.15、0.16、0.27。王立贤等[10]采用公畜外祖父模型和公畜母畜模型估计了高加索细毛羊的初生重遗传力为0.164,母体效应遗传力为0.101;断奶重遗传力为0.076、母体效应的遗传力为0.108。王鹏宇等[11]采用不同动物模型估计了青海细毛羊初生重遗传力为0.1995,断奶重遗传力为0.2552。ABEGAZ等[12]采用单性状动物模型估计了霍罗羊的初生重、断奶重和断奶前平均日增重的遗传力分别是 0.14、0.12、0.13。HANFORD 等[13]利用单性状动物模型估计了哥伦比亚绵羊的初生重遗传力为 0.27,断奶重遗传力为0.16,毛长遗传力为0.55。李文辉等[14]利用半同胞组内相关法估计了甘肃高山细毛羊的初生重遗传力为0.178,断奶重遗传力为0.228,断奶毛长的遗传力为0.252。【本研究切入点】高山美利奴羊新品种是世界首例适应海拔2 400—4 070 m的高山寒旱生态区和羊毛纤维直径以 19.1—21.5 µm为主体的毛肉兼用型美利奴羊新品种,为提高细毛羊综合品质提供了优秀种质资源[4-5]。为了更好的发挥高山美利奴羊的优秀品质,迫切需要对不同生长发育阶段重要经济性状的遗传参数和育种值进行全面评估,以便优化和调整现行育种方案,进行群体的选育提高。目前未全面开展有关高山美利奴羊早期生长性状遗传参数估计等方面的研究。【拟解决的关键问题】本文运用 ASReml软件采用不同动物模型估计高山美利奴羊早期生长性状的遗传参数,研究各模型的方差组分和遗传参数间的差异性,筛选出适合高山美利奴羊早期生长性状遗传参数估计的最佳动物模型,同时估计早期生长性状的遗传参数。并从数量遗传学的角度探讨高山美利奴羊早期生长性状的遗传特性,以期为高山美利奴羊早期阶段群体的育种方案优化、选育提高及科学选种提供科学依据。
1 材料与方法
试验于2016年8月至2017年7月期间在中国农业科学院羊育种工程技术研究中心进行。
1.1试验材料
本研究所用数据资料来自高山美利奴羊培育核心场甘肃省绵羊繁殖技术推广站(原甘肃省皇城绵羊育种试验场)2003—2015年核心群早期生长性状鉴定记录,各性状值均在初生和断奶时测定。通过数据处理,最终筛选出了20 057只高山美利奴羔羊早期生长性状的数据资料,其中早期生长性状包括羔羊初生重(birth weight,BWT)、4 月龄断奶重(weaning weight,WWT)、断奶前平均日增重(average daily gain,ADG)、4月龄断奶毛长(weaning staple length,WSL)、妊娠期(gestation length,GL)。有关早期生长性状的描述性统计见表1。
1.2非遗传因素
根据早期生长性状的原始数据,应考虑的非遗传因素(固定效应)有血统类型、出生年份、配种月份、初生月份、群别、性别以及出生类型。
1.3参数估计模型
建立高山美利奴羊早期生长性状的单性状动物模型如下:


表1 早期生长性状的描述性统计量Table 1 Descriptive statistics of early growth traits
式中,y:各性状的观测值向量;β:固定效应;a:个体加性遗传效应;m:母体遗传效应;p:个体永久环境效应;e:残差效应向量;X、Z、V、W 分别表示固定效应、个体加性遗传效应、个体永久环境效应、母体遗传效应的结构矩阵。
1.4不同模型比较
为了准确估计高山美利奴羊早期生长性状的遗传参数,本研究对高山美利奴羊早期生长性状方差组分估计建立了4个不同动物模型,通过模型估计效果的检验,筛出估计早期生长性状的最优动物模型。
1.4.1利用赤池信息准则(AIC)比较不同模型不同模型方差组分估计准确度的检验可以采用赤池信息准则(AIC)作为评价标准,AIC信息指数的计算公式为:AIC=2k-2lnL,式中L为最大似然函数,k为需要估计的参数个数[15]。AIC信息准则可反映模型中需要估计的参数个数对估计效果的影响,当从一组模型中选择最佳模型时,选择AIC最小的模型,此时模型方差组分估计的效果越好[16]。
1.4.2利用似然比检验(LRT)比较模型利用似然比检验[17]比较不同模型的优劣,其检验统计量为:
其中LR为似然比值,LMAX模型1和模型2分别是两个不同模型下的最大似然函数值,模型1是模型2的子模型,而且LR服从卡方分布,自由度为模型2中估计的参数个数与模型1中估计的参数个数之差。如果卡方检验结果差异显著,说明增加的参数对该性状具有显著影响,否则没有[11]。
1.5方差组分与遗传参数估计
本试验采用ASReml软件进行方差组分和遗传参数的估计,ASReml是一个基于混合线性模型(mixed linear model)分析的统计软件,可做复杂的遗传分析,包括估算随机效应的方差组分、固定效应值和预测随机效应值。ASReml利用限制性极大似然法(restricted maximum likelihood, REML)来估算随机效应的方差组分,并通过混合线性模型方程组(mixed model equations, MME)的求解来估算固定效应值和预测随机效应值。其最突出的优点是它的算法及其误差方差结构和随机效应方差结构的多样性,所采用的平均信息算法(average information, AI)可大大节省计算时间和存储空间[18]。与其他的软件系统相比,在分析遗传学数据时,ASReml内建了一系列方差模型,十分适合分析动物和植物遗传学数据,而且它可以快速、有效地处理大型数据集,比其他通用数据分析软件执行速度更快。同时,在R软件中加载ASReml-R包后,可以结合R强大的数据处理和分析功能,使ASReml可以快速、有效地处理大型数据集,进行遗传参数估计和育种值预测[18-19]。
2 结果
2.1固定效应的显著性判断
本研究考虑的固定效应(非遗传因素)包括血统类型、出生年份、配种月份、初生月份、群别、性别以及出生类型。通过F统计检验,判断固定效应中各因子对高山美利奴羊早期生长性状的影响是否显著。由表2可知,除了性别和出生类型对妊娠期影响不显著外,其余各固定效应对高山美利奴羊早期生长性状的影响均为极显著(P<0.001)。
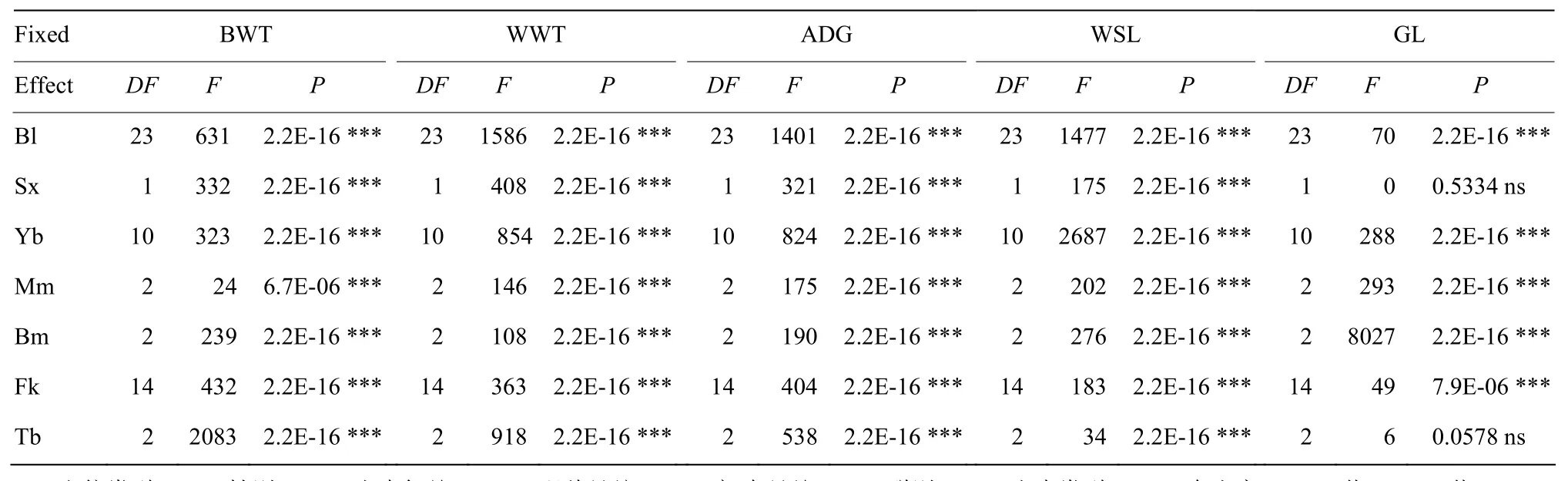
表2 非遗传因素对早期生长性状的影响Table 2 Effects of non-genetic factors on early growth traits
2.2不同动物模型对高山美利奴羊早期生长性状方差组分的估计
2.2.1不同动物模型对初生重方差组分的估计不同动物模型估计的初生重方差组分值见表 3。残差效应(σe2/σy2)在各模型中差别较大,最大的是模型 1和3,为 0.7927;最小的是模型 2和4,为 0.7453。个体加性遗传效应(σa2/σy2)在各模型中差别明显,最大的是模型1和3,为 0.2073±0.0226;最小是模型2 和 4,为 0.0924±0.0160。母体遗传效应(σm2/σy2)在模型2和4中没有差别,为0.1623±0.0113。个体永久环境效应(σs2/σy2)在模型3和4中差别不大。

表3 不同动物模型估计的初生重方差组分Table 3 Variance components estimation of birth weight by different animal models

表4 不同动物模型估计的断奶重方差组分Table 4 Variance components estimation of weaning weight by different animal models
2.2.2不同动物模型对断奶重方差组分的估计不同动物模型估计的断奶重方差组分值见表 4。残差效应(σe2/σy2)在各模型中差异较大,最大的是模型1,为 0.8973;最小的是模型 4,为0.8251。个体加性遗传效应(σa2/σy2)在各模型中差别明显,最大的是模型1和模型3,为 0.1027±0.0159;最小的是模型2,为0.0651±0.0126。母体遗传效应(σm2/σy2)在模型2和模型4差别不大,模型2为0.1098±0.0112,模型4为0.1097±0.0407。个体永久环境效应(σs2/σy2)在模型中差别明显,最大的是模型4为0.0001。
2.2.3不同动物模型对断奶前平均日增重方差组分的估计不同动物模型估计的断奶前平均日增重方差组分值见表 5。残差效应(σe2/σy2)为 0.8422—0.8998。个体加性遗传效应(σa2/σy2)模型 2和 4相等,为0.0681±0.0130;模型 1 为 0.1002±0.0157,模型 3 为0.1001±0.1061,各模型间存在差别。母体遗传效应(σm2/σy2)在模型2和4中没有差别,为0.0898±0.0112。个体永久环境效应(σs2/σy2)在模型中差别明显,最大的是模型3,为0.0001。
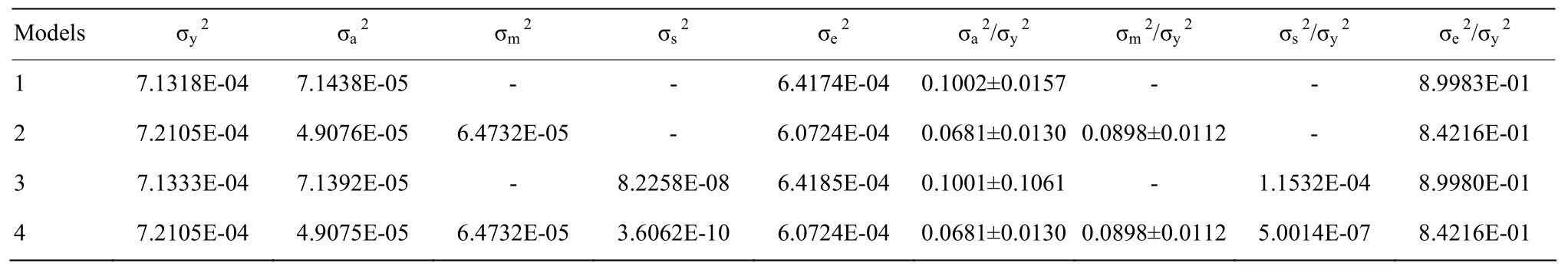
表5 不同动物模型估计的平均日增重方差组分Table 5 Variance components estimation of average daily gain by different animal models
2.2.4不同动物模型对断奶毛长方差组分的估计不同动物模型估计的断奶毛长方差组分值见表 6。残差效应(σe2/σy2)在各模型中差别较大,最大的是模型1,为0.9065;最小的是模型2和4,为0.8962。个体加性遗传效应(σa2/σy2)在各模型中差别较明显,最大的是模型1为0.0937±0.0149;最小是模型 2和4,为0.0865±0.0148。母体遗传效应(σm2/σy2)在模型2和4中没有差别,为0.0173±0.0107。个体永久环境效应(σs2/σy2)在模型3和4中差别不大。
2.2.5不同动物模型对妊娠期组分的估计表 7列出了不同动物模型下妊娠期的方差组分值。各模型的残差效应(σe2/σy2)为0.8621—0.8881。个体加性遗传效应(σa2/σy2)在各模型中差别明显,最大的是模型1和3,为 0.1119±0.0189;最小的是模型2和模型4,为 0.0902±0.0174。母体遗传效应(σm2/σy2)在模型2和4中没有差别,为0.0477±0.0146。个体永久环境效应(σs2/σy2)在各模型中差异不大。

表6 不同动物模型估计的断奶毛长方差组分Table 6 Variance components estimation of weaning staple length by different animal models

表7 不同动物模型估计的妊娠期方差组分Table 7 Variance components estimation of gestation length by different animal models
2.3不同模型的比较
2.3.1利用赤池信息准则(AIC)比较不同模型各早期生长性状不同动物模型的-2lnL值和AIC信息标准值的计算结果列于表8。结果表明,模型2对高山美利奴羊早期生长性状初生重、断奶重、断奶前平均日增重、断奶毛长及妊娠期遗传参数估计的效果最优,说明母体遗传效应对早期生长性状具有重要的影响作用。
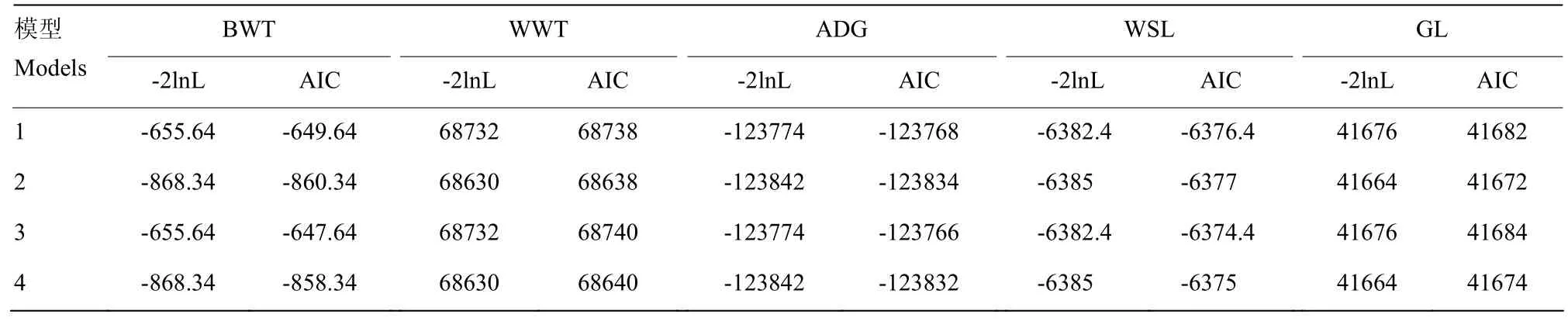
表8 早期生长性状不同动物模型-2lgL值和AIC信息标准值Table 8 Standard values of -2lgL and AIC information for early growth traits in different animal models
2.3.2利用似然比检验(LRT)比较不同模型4种模型相互比较所得到的似然比值及卡方检验的差异显著性列于表 9,结合表 8中的-2lnL值。各模型与模型1进行似然比检验;同时,模型4分别与模型2、3进行似然比检验。结果表明,对于高山美利奴羊早期生长性状的初生重、断奶重和断奶前平均日增重,模型2和4与模型1似然比检验显示彼此间差异极显著(P<0.001),模型4与模型3卡方检验差异极显著(P<0.001),模型4与模型 2卡方检验差异不显著(P>0.05)。对于妊娠期,除模型3与模型1以及模型4与2之间的差异不显著外,其他模型间的差异均极显著(P<0.01)。而对于断奶毛长,各模型彼此间差异不显著(P>0.05)。

表9 早期生长性状不同动物模型卡方检验结果Table 9 Chi-square test results of early growth traits by different animal models
3 讨论
3.1不同动物模型估计效果的比较
从表3—7可知,残差效应在不同动物模型中的差异较大,且在各模型中所占比重最大,这可能与模型所用数据、固定效应数以及各模型中不同随机效应所占比重有关。本研究所用数据为高山美利奴羊早期生产鉴定的原始数据,由于受生产管理水平和生产记录时的误差等条件限制,记录中难免会出现缺失和错误个体的记录;同时,记录中所记载的固定效应有限,使模型中包含的有效固定效应数量减少,而且模型并未考虑固定效应间的二级互作效应,使其隐藏于残差效应中,致使残差效应变大。个体永久环境效应在模型3、4中所占比重很小,对早期生长性状的影响可以忽略不计。王鹏宇等[11]的研究也表明个体环境效应对早期生长性状的影响可以忽略。母体遗传效应在模型2和4中所占比重比早期生长性状中的初生重、断奶重以及断奶前平均日增重的个体加性遗传效应在模型中所占的比重大,在早期生长性状断奶毛长及妊娠期中所占比重较大,这表明早期生长性状遗传参数估计的准确性受母体遗传效应的影响很大。王鹏宇等[20]的研究结果也表明了母体遗传效应对早期生长性状遗传参数估计影响很大。HATCHER等[21]对澳洲美利奴羔羊存活率的研究中也考虑了母体遗传效应的作用,而且羔羊的存活率受母体遗传效应的影响很大。由此可见,母体遗传效应对高山美利奴羊早期生长性状遗传参数估计的准确性起着重要作用。
由表8可知,初生重考虑了个体加性遗传效应和母体遗传效应的模型2,其AIC信息标准值相对于其他各模型更低,表明考虑了个体加性遗传效应和母体遗传效应的模型 2对初生重遗传参数的估计效果最优。由表9结果可知,初生重的模型2和4与模型1彼此间差异极显著,表明母体遗传效应和个体永久环境效应对初生重的遗传参数估计具有显著影响;模型4和模型3比较差异显著,模型4与模型2卡方检验差异不显著,表明母体遗传效应对初生重的影响显著,而个体永久环境效应对初生重的影响不显著。由不同动物模型的-2lnL值和AIC信息标准值综合表明,模型2对断奶重和断奶前平均日增重的遗传参数估计效果最优,说明母体遗传效应对这两个性状的影响显著,而个体永久环境效应对这两个性状影响可以忽略。表9中似然比检验结果显示,对于断奶重和平均日增重两个性状,模型2和模型4与模型1的似然比检验差异显著,模型4与模型3卡方检验差异极显著,模型4与模型2卡方检验差异不显著,这也表明母体遗传效应对断奶重和日增重两个性状有显著影响,个体永久环境效应的影响并不显著。断奶毛长的各模型-2lnL值和AIC信息标准值差异并不是很明显,相比之下,模型2的AIC信息标准值相对于其他各模型更低,表明考虑了个体加性遗传效应和母体遗传效应的模型 2对断奶毛长遗传参数估计效果最佳。但由表9似然比检验结果可知,断奶毛长的各模型彼此间差异不显著,表明母体遗传效应和个体永久环境对断奶毛长性状在0.05显著水平上的影响不显著。对于妊娠期,考虑了个体加性遗传效应和母体遗传效应的模型 2,其 AIC信息标准值相对于其他各模型更低,表明模型2对妊娠期性状的遗传参数估计效果最优。通过似然比检验结果可知,对于妊娠期,除模型3与模型1、模型4与2之间的差异不显著外,其他模型间的差异均极显著,这表明羔羊妊娠期受母体遗传效应的影响显著,而受个体永久环境效应的影响并不显著。由此可见,母体遗传效应对早期生长性状具有显著影响,而个体永久环境效应对早期生长性状的影响可以忽略。王鹏宇等[11,22]同样证明母体遗传效应对早期生长性状有显著影响,而个体永久环境效应对早期生长性状的影响不显著。MANIATIS等[23]的研究结果表明,对于早期生长性状遗传参数估计中,若模型中不包含母体遗传效应时,所估计的遗传参数会偏高。KUSHWAHA等[24]的研究中同样表明了这一点。
综上所述以及由各模型的AIC值和似然比检验结果可知,模型2更适用于高山美利奴羊早期生长性状遗传参数的估计。
3.2不同动物模型对高山美利奴羊早期生长性状遗传参数的估计
从表3—7可知,各模型中不同随机效应对高山美利奴羊早期生长性状遗传参数估计具有不同程度的影响,其中母体遗传效应在初生重、断奶重和断奶前平均日增重性状的遗传参数估计中所占比重比加性遗传效应都大,在断奶毛长和妊娠期中所占比重比加性遗传效应小;个体永久环境效应在早期各生长性状遗传参数估计中所占比重极小。ZHANG等[25]研究结果表明,当模型中所考虑的随机效应对性状的影响可以忽略时,所估计性状的遗传力均成上升趋势。HOSSEINZADEH等[26]研究表明,不同模型中考虑的母体遗传效应、母体永久环境效应、个体永久环境效应或其组合对生长性状遗传参数估计具有不同程度的影响。OSORIO-AVALOS等[27]在研究西班牙美利奴羔羊早期生长性状也考虑了母体遗传效应对性状的影响。王鹏宇等[11,20,22]在研究青海细毛羊不同阶段的生长性状时,也考虑到了母体遗传效应、母体永久环境效应、个体永久环境效应及其相互组合间效应对不同性状的影响。由此说明,在高山美利奴羊早期生长性状遗传参数估计过程中,母体遗传效应对遗传参数估计的准确性具有重要影响,而个体永久环境效应对早期各生长性状遗传参数估计的影响可以忽略。
由表 3—7可知,各模型估计的初生重遗传力为(0.0924±0.0160)—(0.2073±0.0226),母体效应的遗传力为 0.1623±0.0113;断奶重遗传力为(0.0651±0.0126)—(0.1027±0.0159),母体效应的遗传力为(0.1097±0.0407)—(0.1098±0.0112);断奶前平均日增重的遗传力为(0.0681±0.0130)—(0.1001±0.1061),母体效应的遗传力为 0.0898±0.0112。JALIL-SARGHALE等[28]估计了Baluchi羊的早期生长性状的遗传参数,其初生重遗传力为 0.062±0.02,母体遗传效应为0.09±0.02;断奶重遗传力为0.12±0.02,母体遗传效应为0.04±0.01;断奶前平均日增重遗传力为 0.08±0.02,母体遗传效应为 0.03±0.01。DI等[9]估计的中国超细型美利奴羊的初生重为0.15,断奶重遗传力为0.16,断奶毛长遗传力为0.27。 BOUJENANE等[29]估计了萨尔迪羊的初生重遗传力为0.07±0.02,母体遗传效应为0.13±0.03;断奶重遗传力为0.05±0.02,断奶前平均日增重遗传力为 0.01±0.01。SINGH等[30]估计了马尔瓦羊的初生重遗传力为 0.28±0.058,断奶重遗传力为 0.27±0.050,断奶前平均日增重遗传力为0.26±0.050。JAFAROGHLI等[31]估计了莫加尼羊的初生重遗传力为0.07,母体遗传效应为0.18;断奶重遗传力为0.09,母体遗传效应为0.06;断奶前平均日增重遗传力为0.08。BOUJENANE等[32]采用动物模型估计了D’man羊早期生长性状的遗传参数,其初生重遗传力为 0.05±0.02,母体遗传效应为 0.10±0.02。王立贤等[10]估计了高加索细毛羊初生重遗传力为 0.164、母体效应遗传力为0.101,断奶重遗传力为0.076、母体效应的遗传力为0.108;WYK等[33]估计的德美羔羊的初生体重的遗传力是 0.13—0.23,断奶重的遗传力为 0.07—0.09。SAFARI等[34]估计的澳大利亚美利奴羊的初生重遗传力为0.18,母体效应的遗传力为0.19;断奶重的遗传力为0.29,母体效应的遗传力为0.12。断奶毛长的遗传力为(0.0865±0.0148)—(0.0937±0.0149),母体效应的遗传力为 0.0173±0.0107。DI等[9]估计的超细型中国美利奴羊断奶毛长的遗传力为0.27,杨平贵[35]估计的凉山半细毛羊早期性状断奶毛长的遗传力为0.13,李文辉等[14]估计了甘肃高山细毛羊的断奶毛长遗传力为 0.252。妊娠期的遗传力为(0.0902±0.0174)—(0.1119±0.0189),母体效应的遗传力为 0.0477±0.0146。LI等[36]估计了澳大利亚羊的妊娠期遗传力为0.52±0.02,母体遗传力为0.13±0.02,初生重遗传力为 0.15±0.01,母体遗传效应为 0.15±0.01;OSINOWO等[37]估计了Yankasa羊的初生重遗传力为0.05,妊娠期的遗传力为0.02。
上述研究结果表明,本研究与部分的研究结果存在一定的差异性,但还是与一些研究结果较一致,造成这种差异的原因可能与本研究的品种特性及所使用的数据有关。虽然本研究利用了20 057只高山美利奴羔羊的早期生长性状数据,同时也考虑了多个非遗传因素对早期生长性状的影响,但数据量和记录的非遗传因素仍较少,使得估计出的遗传参数与真实的遗传参数仍存在一定偏差。MANIATIS等[23]研究表明,不同数据记录量、数据内容和结构对早期生长性状遗传参数的准确估计具有重要作用。同时,这可能与高山美利奴羊的特殊生活环境、饲养管理、品种特性、数据结构和遗传结构有关。BRADFORD[38]的研究表明,品种、营养水平、母羊年龄、每胎产羔数、季节等因素通过母羊这一途径会不同程度的作用在羔羊上。而本研究中各模型所考虑的随机效应不同,使各模型的数据结构存在一定的差异性,从对各生长性状遗传参数的准确估计也产生了一定的影响。
4 结论
4.1通过不同模型的比较分析,确定模型2为高山美利奴羊早期生长性状遗传参数估计的最佳动物模型;高山美利奴羊早期生长性状受母体遗传效应的影响显著,而受个体永久环境效应的影响可以忽略不计。
4.2基于最佳动物模型估计出的高山美利奴羊早期生长性状的遗传参数分别是初生重的遗传力为0.0924±0.0160,母体效应的遗传力为0.1623±0.0113;断奶重的遗传力为0.0651±0.0126,母体效应的遗传力为 0.1098±0.0112;断奶前平均日增重的遗传力为0.0681±0.0130,母体效应的遗传力为0.0898±0.0112;断奶毛长的遗传力为0.0865±0.0148,母体效应的遗传力为 0.0173±0.0107;妊娠期的遗传力为 0.0902±0.0174,母体效应的遗传力为0.0477±0.0146。
致谢:本研究所用数据资料由甘肃省绵羊繁殖技术推广站提供,在此深表感谢。
[1]白俊艳, 李金泉, 贾小平, 张勤, 道尔吉. 母体遗传效应对绒山羊生产性状遗传参数估计的影响. 遗传, 2006, 28(9): 1083-1086.BAI J Y, LI J Q, JIA X P, ZHANG Q, DAO E J. Influence of maternal genetic effect on genetic parameter estimates of production traits of cashmere goat. Hereditas, 2006, 28(9): 1083-1086. (in Chinese)
[2]张沅, 张勤. 畜禽育种中的线性模型. 北京: 北京农业大学出版社,1993.ZHANG Y, ZHANG Q. Linear Models in Animal Breeding. Beijing:Beijing Agricultural University Press, 1993. (in Chinese)
[3]魏永龙. 白绒山羊生长性状遗传评估模型及遗传参数估计的研究[D]. 呼和浩特:内蒙古农业大学, 2014.WEI Y L. Study of genetic evaluation model and estimation of genetic parameters of growth traits in White Cashmere Goats[D]. Huhhot:Inner Mongolia Agricultural University, 2014. (in Chinese)
[4]岳耀敬, 王天翔, 刘建斌, 郭健, 李桂英, 孙晓萍, 李文辉, 冯瑞林,牛春娥, 郭婷婷, 李范文, 杨博辉. 高山美利奴羊新品种种质特性初步研究. 中国畜牧杂志, 2014, 50(21): 16-19.YUE Y J, WANG T X, LIU J B, GUO J, LI G Y, SUN X P, LI W H,FENG R L, NIU C E, GUO T T, LI F W, YANG B H. Preliminary study on Gansu Alpine Merino sheep new varieties of germplasm characteristics. Chinese Journal of Animal Science, 2014, 50(21):16-19. (in Chinese)
[5]杨博辉. 论高山美利奴羊新品种的价值和意义. 甘肃畜牧兽医,2017, 47(04): 55-56.YANG B H. On the value and significance of new varieties of alpine merino. Gansu Animal Husbandry and Veterinary, 2017, 47(04):55-56. (in Chinese)
[6]BORG R C, NOTTER D R, KOTT R W. Phenotypic and genetic associations between lamb growth traits and adult ewe body weights in western range sheep. Journal of Animal Science, 2009, 87(11):3506-3514. doi: 10. 2527/jas. 2008-1622.
[7]HANFORD K J, VAN VLECK L D, SNOWDER G D. Estimates of genetic parameters and genetic change for reproduction, weight, and wool characteristics of Targhee sheep. Journal of Animal Science,2003, 81(3): 630-640. doi: 10. 2527/2003. 813630x.
[8]CLOETE S W P, MISZTAL I, OLIVIER J J. Genetic parameters and trends for lamb survival and birth weight in a Merino flock divergently selected for multiple rearing ability. Journal of Animal Science, 2009, 87(7): 2196-208. doi: 10. 2527/jas. 2008-1065.
[9]DI J, ZHANG Y, TIAN K C, LAZATE, LIU J F, XU X M, ZHANG Y J, ZHANG T H. Estimation of (co)variance components and genetic parameters for growth and wool traits of Chinese superfine Merino sheep with the use of a multi-trait animal model. Livestock Science,2011, 138(1/3): 278-288. doi: 10. 1016/j. livsci. 2011. 01. 005.
[10]王立贤, 张沅, 潘君乾. 绵羊生长性状母本效应方差组分、遗传参数估计的研究. 畜牧兽医学报, 1993, 24(4): 289-293.WANG L X, ZHANG Y, PAN J Q. Study of estimating variance components and genetic parameters of maternal influenced sheep growth trait. Chinese Journal of Animal and Veterinary Sciences, 1993,24(4): 289-293. (in Chinese)
[11]王鹏宇, 官却扎西, 祁全青, 德毛, 张燕军, 王瑞军, 张文广, 李金泉. 不同模型对青海细毛羊生长性状遗传参数估计的比较. 中国农业科学, 2012(11): 2280-2287. doi: 10. 3864/j. issn. 0578-1752.2012. 11. 018.WANG P Y, GUANQUE Z X, QI Q Q, DE M, ZHANG Y J, WANG R J, ZHANG W G, LI J Q. Comparison and analysis of genetic parameters of growth traits of Qinghai Fine-Wool Sheep estimated by different models. Scientia Agricultura Sinica, 2012(11): 2280-2287.doi: 10. 3864/j. issn. 0578-1752. 2012. 11. 018. (in Chinese)
[12]ABEGAZ S, WYK J V, OLIVIER J J. Model comparisons and genetic and environmental parameter estimates of growth and the Kleiber ratio in Horro sheep. South African Journal of Animal Science, 2005,35(1): 30-40. doi: 10. 4314/sajas. v35i1. 4046.
[13]HANFORD K J, SNOWDER G D, VAN VLECK L D. Models with nuclear, cytoplasmic, and environmental effects for production traits of Columbia sheep. Journal of Animal Science, 2003, 81(8): 1926. doi:10. 2527/2003. 8181926x.
[14]李文辉, 苏文娟, 保国俊. 甘肃高山细毛羊遗传参数估计. 中国畜牧杂志, 1997(06): 24-25+31.LI W H, SU W J, BAO G J. Estimation of genetic parameters of Gansu alpine fine wool sheep. Chinese Journal of Animal Science,1997 (06): 24-25+31. (in Chinese)
[15]GOWANE G R, CHOPRA A, PRINCE L L, PASWAN, ARORA A L.Estimates of (co)variance components and genetic parameters for body weights and first greasy fleece weight in Bharat Merino sheep.Animal an International Journal of Animal Bioscience, 2010,4(3):425-31. doi: 10. 1017/S1751731109991157.
[16]李丽娟, 杨莉, 申小云, 宋德荣. 不同模型对贵州半细毛羊的遗传参数估计比较. 畜牧与兽医, 2014(07): 37-40.LI L J, YANG L, SHEN X Y, SONG D R. Comparison of different models for estimating genetic parameters of Guizhou semi - fine wool sheep. Animal Husbandry & Veterinary Medicine, 2014(07): 37-40.(in Chinese)
[17]白俊艳, 李金泉, 道尔吉, 张勤. 用不同模型估计绒山羊早期生长性状遗传参数的比较. 遗传学报, 2004, 31(6): 578-581.BAI J Y, LI J Q, DAOERJI, ZHANG Q. Comparison of different models for estimating genetic parameters of early growth traits in Cashmere goat. Journal of Genetics and Genomics, 2004, 31(6):578-581. (in Chinese)
[18]林元震, 陈晓阳. R与ASReml—R统计分析教程. 北京:中国林业出版社, 2014: 198-199.LIN Y Z, CHEN X Y. R & ASReml-R Statistical Analysis Tutorial.Beijing: China Forestry Publishing House, 2014: 198-199. (in Chinese)
[19]边黎明, 施季森, 童春发, 郑仁华. ASReml及其在林木遗传分析中的应用前景. 林业科技开发, 2012(04): 9-12.BIAN L M, SHI J S, TONG C F, ZHENG R H. ASReml and its application prospect in forest genetic analysis. Journal of Forestry Engineering, 2012(04): 9-12. (in Chinese)
[20]王鹏宇, 官却扎西, 祁全青, 德毛, 张文广, 李金泉. 母体遗传效应对青海细毛羊生产性能遗传参数估计的影响. 遗传, 2012(05):584-590. doi: 10. 3724/SP. J. 1005. 2012. 00584.WANG P Y, GUANQUE Z X, QI Q Q, DE M, ZHANG W G, LI J Q.Impact of maternal genetic effect on genetic parameter estimation of production traits for Qinghai fine-wool sheep. Hereditas, 2012 (05):584-590. doi: 10. 3724/SP. J. 1005. 2012. 00584. (in Chinese)
[21]HATCHER S, ATKINS K D, Safari E. Lamb survival in Australian Merino sheep: A genetic analysis. Journal of Animal Science, 2010,88(10): 3198. doi: 10. 2527/jas. 2009-2461.
[22]王鹏宇. 青海细毛羊封闭与开放核心群育种规划效果评估[D]. 呼和浩特:内蒙古农业大学, 2012.WANG P Y. Evaluating the breeding efficiency of closed and open nucleus breeding scheme for Qinghai Fine-Wool Sheep[D]. Huhhot:Inner Mongolia Agricultural University, 2012. (in Chinese)
[23]MANIATIS N, POLLOTT G E. The impact of data structure on genetic (co)variance components of early growth in sheep, estimated using an animal model with maternal effects. Journal of Animal Science, 2003, 81(1): 101-108. doi: 10. 2527/2003. 811101x.
[24]KUSHWAHA B P, MANDAL A, ARORA A L, KUMAR R, KUMAR S, NOTTER D R. Direct and maternal (co)variance components and heritability estimates for body weights in Chokla sheep. Journal of Animal Breeding and Genetics, 2009, 126(4): 278-287. doi: 10. 1111/ j.1439-0388. 2008. 00771. x.
[25]ZHANG C, YANG L, ZHONG S. Variance components and genetic parameters for weight and size at birth in the Boer goat. Livestock Science, 2008, 115(1): 73-79. doi: 10. 1016/j. livsci. 2007. 06. 008.
[26]HOSSEINZADEH N G, ARDALAN M. Comparison of different models for the estimation of genetic parameters of body weight traits in Moghani sheep. Agricultural & Food Science, 2010, 19(3):207-213. doi: 10. 2137/145960610792912639.
[27]OSORIO-AVALOS J, MENENDEZ-BUXADERA A, SERRADILLA J M, ALCALA A M. Comparison of genetic parameters of weight traits estimated using cross sectional and longitudinal analyses of animal models with two types of contemporary groups in Spanish Merino lambs. Small Ruminant Research, 2017, 151: 45-51. doi: 10.1016/j. smallrumres. 2017. 04. 007.
[28]JALIL-SARGHALE A, KHOLGHI M, SHAHREBABAK M M,SHAHREBABAK H M, MOHAMMADI H, ABDOLLAHIARPANAHI R. Model comparisons and genetic parameter estimates of growth traits in Baluchi sheep. Slovak Journal of Animal Science,2014, 47(1): 12-18.
[29]BOUJENANE I, CHIKHI A, IBNELBACHYR M, MOUH F Z.Estimation of genetic parameters and maternal effects for body weight at different ages in D’man sheep. Small Ruminant Research, 2015,130: 27-35. doi: 10. 1016/j. smallrumres. 2015. 07. 025.
[30]SINGH H, PANNU U, NARULA H K, CHOPRA A, NAHARWARA V, BHAKAR S K. Estimates of (co)variance components and genetic parameters of growth traits in Marwari sheep. Journal of Applied Animal Research, 2016, 44(1): 27-35. doi: 10. 1080/09712119. 2014.987291.
[31]JAFAROGHLI M, RASHIDI A. (Co)variance components and genetic parameter estimates for growth traits in Moghani sheep. Small Ruminant Research, 2010, 91(2): 170-177. doi: 10. 1016/j.smallrumres. 2010. 03. 010.
[32]BOUJENANE I, DIALLO I T. Estimates of genetic parameters and genetic trends for pre-weaning growth traits in Sardi sheep. Small Ruminant Research, 2017, 146(1): 61-68. doi: 10. 1016/j. smallrumres.2016. 12. 002.
[33]WYK J V, FAIR M D, CLOETE S. Revised models and genetic parameter estimates for production and reproduction traits in the Elsenburg Dormer sheep stud. South African Journal of Animal Science, 2003, 33(4): 213-222.
[34]SAFARI E, FOGARTY N M, GILMOUR A R, ATKINS K D,MORTIMER S I, SWAN A A, BRIEN F D, GREEFF J C, VAN DER WERF J H J. Across population genetic parameters for wool, growth,and reproduction traits in Australian Merino sheep. 2. Estimates of heritability and variance components. Crop & Pasture Science, 2007,58(2): 177-184.
[35]杨平贵. 凉山半细毛羊早期性状遗传参数的估测及羔羊产肉性能研究[D]. 雅安:四川农业大学, 2003.YANG P G. Estimation of genetic parameters of early traits in Liangshan semi-fine wool and study on performance of lamb[D]. Yaan:Sichuan Agricultural University , 2003. (in Chinese)
[36]LI L, BROWN D J. Estimation of genetic parameters for lambing ease, birthweight and gestation length in Australian sheep. Animal Production Science, 2016, 56: 934-940. doi: 10. 1071/AN14129
[37]OSINOWO O A, ABUBAKAR B Y, TRIMNELL A R. Genetic and phenotypic relationships between gestation length, litter size and litter birth weight in Yankasa sheep. Animal Reproduction Science, 1993,34(2): 111-118. doi: 10. 1016/0378-4320(93)90069-4.
[38]BRADFORD G E. The role of maternal effects in animal breeding.VII. Maternal effects in sheep. Journal of Animal Science, 1972, 35(6):1324-1334. doi: 10. 2527/jas1972. 3561324x
