注意缺陷多动障碍患儿生物运动感知能力的研究
2018-03-30叶小飞胡学会胡盼盼汪晓翠沈丽伟
叶小飞,胡学会,胡盼盼,杨 斌,汪晓翠,沈丽伟,汪 凯
生物运动是指生命体(人类或动物)在空间上整体移动的行为,如步行、奔跑等[1]。生物运动感知包含了对运动刺激物本身、他或她的动作、意图甚至情绪的识别,帮助人类洞悉生物运动所要表达的真正意图,继而增强个体的社会适应能力。脑功能成像研究发现,颞上沟及额下回、中央前回及额中回等是生物运动感知的关键脑区[2]。注意缺陷多动障碍(attention deficit hyperactivity disorder,ADHD)是学龄期儿童常见的一类心理障碍性疾病,主要症状为与年龄不相符的注意缺陷、多动及冲动行为,常合并学习障碍。既往研究表明ADHD患儿在大脑结构及功能上存在损害,Jackson et al[3]应用磁共振成像发现ADHD患儿的前额叶皮质、基底节及胼胝体均存在异常。该研究应用生物运动的时距辨别任务,对ADHD患儿的生物运动感知能力进行测试,旨在探讨其是否存在生物运动感知能力障碍。
1 材料与方法
1.1病例资料
1.1.1ADHD组 2015年12月~2016年12月在安徽省儿童医院就诊的ADHD患儿46例,其中男41例,女5例,发病年龄6~15(9.54±2.09)岁,病程3月~4年,平均(1.91±0.99)年,平均受教育年限(3.34±2.01)年,均为右手利。ADHD患儿均为来本院初诊患儿。纳入标准:① 符合DSM-IV中有关ADHD诊断标准;② 无先天性疾病及相关精神疾病;③ 所有ADHD儿童未服用任何药物治疗,也未采用其他手段进行过干预;④ 年龄6岁以上,视听功能正常,可理解并执行整个实验过程。排除标准:① 精神发育迟滞,采用中国修订版联合型瑞文智力测试标准型(CRT-S1),智商<70分;② 包括已知的神经系统疾病或广泛性发育障碍;③ 合并其他慢性疾病如哮喘、糖尿病等。
1.1.2健康对照组 同期在本院体检的健康儿童46例,均正常入学,发育正常,右手利,视、听能力正常,既往无神经、精神疾病史。在性别、年龄、受教育年限与ADHD患儿相匹配。
所有入选儿童在试验前被告知本次试验的目的及意义等,同时征得受试儿童及其法定监护人同意,签署知情同意书,本研究通过安徽省儿童医院医学研究与伦理委员会的审核批准。
1.2方法
1.2.1神经心理学背景测试 包括瑞文智力测试、数字广度评分。
1.2.2时距辨别任务 试验应用一台14英寸的联想笔记本电脑,在Matlab程序运行下,电脑屏幕呈现刺激(刷新频率为60 Hz),受试儿童双眼距显示屏50 cm。
1.2.2.1 生物运动序列 即正立的光点运动序列,通过采用Vanrie和 Verfaillie三维运动捕捉系统改进的光点序列[4]。该序列是将行走过程中人的关节的位置数字化编码为可运动的光点而获得的。
1.2.2.2 非生物运动序列 即倒置的光点运动序列,是将经典的正立生物运动序列旋转180°倒置后获得。
1.2.2.3 试验程序 试验范式为白色的光点刺激物在黑色的背景呈现,每试次中生物运动刺激和非生物运动刺激相继呈现。一组序列(生物运动和非生物运动)呈现结束后,要求受试儿童判断哪一个刺激的持续时距更长,告知被试不能数数或默念计时,且只有在完成上一组选择后,方可呈现下一组序列,试验由70个试次组成,每20~30个试次中间有一个休息间歇。试验总共约30 min。所有受试儿童完成该测试的依从性均较好,且本研究中涉及的所有测试任务均经过专业培训的人员进行操作,见图1。
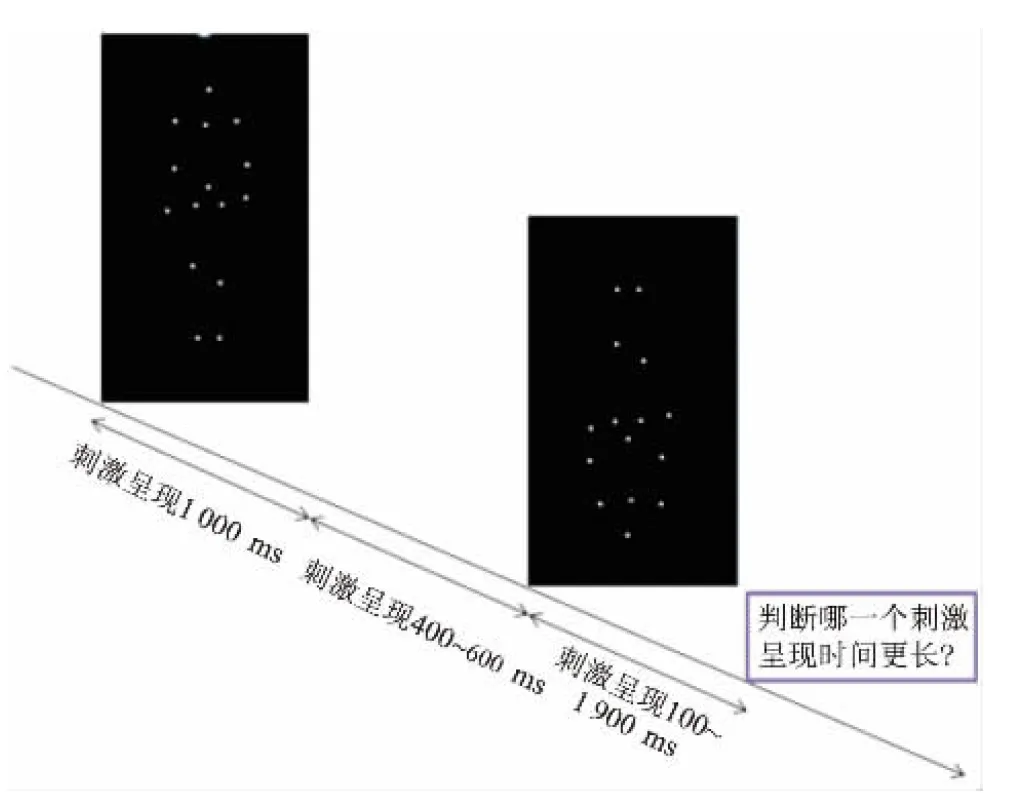
图1 时距辨别任务示意图
1.2.3数据分析 所有受试儿童的测试结果均被导入Boltzmann S型函数中,横坐标代表生物运动序列与非生物运动序列的时距差,共7个点,赋值-3~3,即表示两种刺激的时距差从-900~ +900 ms,每个单位长度为300 ms。纵坐标代表选择生物运动序列为“长时距”的反应百分比。主观相等点(the point of subjective equality,PSE)则表示受试在该点感知到的生物运动刺激与非生物运动刺激的持续时距相等。PSE值由Boltzmann方程的中点计算而来f(x)=1/(1+exp[(x-x0)/ω]),PSE值为负值时代表生物运动具有时距延长效应,即正立的生物运动序列比倒置的非生物运动序列被感知到的时距更长,时距延长的具体数值则为负值的绝对值乘以 300,单位为ms。正的PSE值则与之相反,即时距压缩[5]。
1.3统计学处理采用SPSS 17.0 进行统计学分析。性别属于定性资料,采用χ2检验;年龄、教育年限、瑞文智商数及PSE值属于计量资料,且符合正态分布,采用独立样本t检验进行组间比较,对照组PSE值与检验值“0”的比较则应用单样本t检验;正序及倒序数字广度属于计量资料,但为非正态分布,采用Mann-Whitney U检验进行组间比较。ADHD患儿的PSE值与年龄、教育年限、病程、数字广度、瑞文智商及发病年龄的相关关系采用Pearson相关分析,P<0.05表示差异有统计学意义。
2 结果
2.1两组儿童一般资料比较与同龄的健康儿童相比,ADHD患儿在性别、年龄、教育年限及瑞文智商方面差异均无统计学意义,但两组间的正序数字广度评分(z=-4.04,P<0.05)和倒序数字广度评分(z=-3.71,P<0.05)差异均有统计学意义,见表1。
2.2两组儿童时距辨别结果比较与检验值“0”相比,健康对照组的PSE为负值(-0.30±0.35),差异有统计学意义(t=-5.83,P<0.05)。ADHD患儿的PSE为正值(0.06±0.63),与健康对照组相比,其值明显增大,两组间的独立样本t检验,差异有统计学意义(t=0.34,P<0.05)。
2.3ADHD患儿时距辨别的相关分析Pearson相关分析结果表明,ADHD患儿的PSE值与患儿的年龄、教育年限、病程、数字广度及瑞文智商均无明显相关性,但与患儿发病年龄相关(r=-0.91,P<0.05),即发病年龄越小,患儿生物运动感知能力越差,见表2。
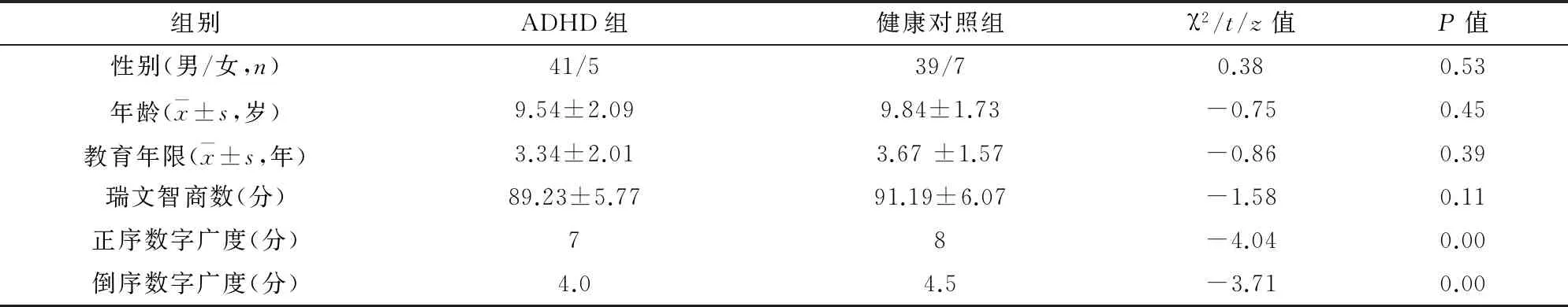
表1 ADHD组与健康对照组的一般资料比较 (n=46)

表2 ADHD患儿的PSE值与年龄、教育年限、病程、数字广度、瑞文智商及发病年龄相关分析
3 讨论
生物运动能够提供非常丰富的社会信息,如身份、性别、情绪等,同时这种信息获取的方式对于远距离的沟通、交流、模仿学习各种复杂的运动行为至关重要,特别是当形状、颜色等属性特征模糊不清的时候就显的更为重要[1,6]。心理学家 Johansson[7]通过在人体的某些重要关节处(如肩、肘、踝等)标以信号,进而拍摄记录人的运动过程中这些关节的运动轨迹,继而得到光点运动序列(生物运动),受试者通过光点运动序列识别出运动人物的重要信息。然而,当光点运动序列倒置过来后(非生物运动),这种加工优势明显受损,也就是说在呈现时间相同的情况下,被试感知到正立的含有生物运动信息的生物运动序列的呈现时间要长于倒置的不含有生物运动信息的非生物运动序列,称为“倒置效应”[8]。
本研究显示,ADHD患儿的生物运动感知能力受损,表现为生物运动的时距延长效应消失。ADHD是一类与遗传、家庭社会环境、脑结构与功能异常、神经递质代谢异常等有关的疾病。Cortese et al[9]发现,与同龄正常儿童相比,ADHD患儿的全脑体积减小,主要表现在额叶、顶叶、枕叶、颞叶。Silk et al[10]的研究发现,ADHD患儿的全脑皮层灰质厚度变薄,且枕、顶叶和额、颞叶的纤维联络减少,存在脑发育成熟障碍。这与国内学者卢又燃等[11]应用fMRI发现ADHD患儿额叶一基底神经节环路功能异常的结果相一致。上述的研究证实,ADHD患儿存在额叶结构与功能的缺失。
众多研究[12]表明,生物运动感知是由一个广泛的神经网络负责,其中镜像神经元系统较多的用于生物运动感知的神经机制研究。与非生物运动刺激相比,受试者观察生物运动刺激时,其在颞上沟、中央前回、额中回及额下回诱发的神经活动更强,fMRI的同步研究也证实,受试者观察生物运动时,其下颞皮质、颞叶侧部及额下回活性明显增加[2,13]。综上研究,ADHD的发病机制、生物运动均与大脑额叶功能关系密切,据此推测,ADHD患儿的生物运动感知功能受损,可能与额叶功能异常有关。
相关分析表明,ADHD患儿生物运动感知能力与年龄、教育年限、病程、数字广度及瑞文智商均无显著相关,但与发病年龄呈负相关性。众所周知,额叶、扣带回多巴胺系统参与认知功能。哌甲酯能阻断多巴胺转运体重吸收、扩大多巴胺受体响应时间从而改善ADHD症状[14],支持AHDH的发病与多巴胺能系统异常有关。而多巴胺受体密度与儿童发育关系密切。因此,本研究表明ADHD患儿生物运动感知能力障碍可能与额叶-纹状体中的多巴胺系统异常有关,且发病年龄越小,其生物运动感知能力越差。
在神经心理学背景测试中,ADHD患儿的正、倒序数字广度得分低于正常同龄儿童。而数字广度试验属于执行功能的一部分,需要额叶参与完成,提示ADHD患儿数字广度表现差可能与额叶功能异常有关,同时既往研究[15]显示额叶功能在生物运动感知能力表现上至关重要。因此,背景测试的结果更进一步支持了我们的推测,即ADHD患儿生物运动感知功能障碍可能与额叶受损有关。
综上所述,ADHD患儿存在生物运动感知能力障碍,发病年龄越小,生物运动感知能力表现越差,且可能与额叶受损有关。由于此次研究对象均为新确诊且未进行任何干预的ADHD患儿,因此对其是否进行早期干预以及何种干预有待进一步长期随访、研究。
[1] 蒋 毅,王 莉. 生物运动加工特异性:整体结构和局部运动的作用[J]. 心理科学进展,2011,19(3):301-11.
[2] Saygin A P, Wilson S M, Hagler D J, et al. Point-light biological motion perception activates human premotor cortex[J]. J Neurosci,2004,24(27):6181-8.
[3] Jackson J N, MacKillop J. Attention-deficit/hyperactivity disorder and monetary delay discounting: a meta-analysis of case-control studies[J]. Biol Psychiatry Cogn Neurosci Neuroimaging,2016,1(4):316-25.
[4] Vanrie J,Verfaillie K. Perception of biological motion:A stimulus set of human point-light actions[J]. Behav Res Methods Instrum Comput,2004,36(4):625-9.
[5] Wearden J H,Ferrara A.Stimulus range effects in temporal bisection by humans[J].Q J Exp Psychol B,1996,49(1):24-44.
[6] Henry J D,Thompson C,Rendell P G, et al. Perception of biological motion and emotion in mild cognitive impairment and dementia[J].J Int Neuropsychol Soc, 2012,18(5):866-73.
[7] Johansson G.Visual perception of biological motion and a model for its analysis[J]. Percept Psychophys,1973,14(2):201-11.
[8] Chang D H. Acceleration carries the local inversion effect in biological motion perception[J]. J Vis,2009,9(1):1-17.
[9] Cortese S,Castellanos F X.The relationship between ADHD and obesity: implications for therapy[J].Expert Rev Neurother,2014,14(5):473-9.
[10] Silk T J, Vilgis V, Adamson C, et al. Abnormal asymmetry in frontostriatal white matter in children with attention deficit hyperactivity disorder[J]. Brain Imaging Behav, 2016, 10(4): 1080-9.
[11] 卢又燃,耿道颖,冯晓源,等. 注意缺陷多动障碍儿童语义工作记忆的功能MRI研究[J]. 中华放射学杂志,2006,40(11):1129-33.
[12] 操瑞花,叶 星,田仰华,等. 帕金森病患者生物运动感知能力研究[J]. 中华行为医学与脑科学杂志,2014,23(3):193-5.
[13] Pelphrey K A,Carter E J. Brain mechanisms for social perception:lessons from autism and typical development[J]. Ann N Y Acad Sci,2008,1145:283-99.
[14] Castellanos F X,Aeosta M T.Towards an understanding of the molecular mechanisms underlying the pharmacological treatments of attention deficit hyperactivity disorder[J]. Rev Neurol,2011,52 Suppl 1:S155-60.
[15] 刘婷婷,操瑞花,胡盼盼,等. 早中期帕金森患者的杂乱生物运动感知障碍[J]. 中国神经精神疾病杂志,2015,41(5):257-61.
