盐度和pH对长肋日月贝耗氧率和排氨率的影响
2018-03-27严俊贤陈明强吴开畅李有宁马振华
严俊贤,陈明强,吴开畅,李有宁,胡 静,赵 旺,马振华
(1.中国水产科学研究院 南海水产研究所,热带水产研究开发中心,海南 三亚,572018;2.中国水产科学研究院 南海水产研究所,农业部南海渔业资源开发利用重点实验室,广东 广州,510300)
长肋日月贝(Amusiumpleuronectes)是我国南海海域的食用贝类,俗称飞螺,属扇贝科、日月贝属,该贝富含谷氨酸、天冬氨酸、丙氨酸等呈味氨基酸,味道鲜美,具有很高的食用价值[1-2]。目前,国内对长肋日月贝的繁育与养殖开发正处于初级阶段,相关的研究报道较少,王雨等[3]对长肋日月贝的个体发育及其人工育苗技术进行了初步研究,付玉等[4]对广西省北部湾的野生长肋日月贝的资源分布进行了调查评估,并研究了其性状间的相关关系。国外则较早就对长肋日月贝展开研究,在人工繁育、遗传多样性、生态资源等方面均有报道[5-7]。
能量学研究是研究生物新陈代谢水平的一个重要手段,其主要指标包括耗氧率和排氨率,能够反映研究对象对环境的适应性能和养殖特性等,对其人工养殖和生态研究具有重要意义。国内外对不同水产动物的能量学研究均有报道,包括虾夷扇贝(Patinopectenyessoensis)[8]、泥蚶(Tegillarcagranosa)[9]、圣保罗对虾(Farfantepenaeuspaulensis)[10]、大弹涂鱼(Boleophthalmuspectinirostris)[11]、瓦氏黄颡鱼(Pelteobagrusvachelli)[12]、花尾胡椒鲷(Plectorhinchuscinctus)[13]和大菱鲆(Scophalmusmaximus)[14]等,然而目前有关长肋日月贝的能量学研究尚未见报道。人工养殖的近海区域环境容易受到雨水和人为因素的影响,其理化因子的变化相对较多,笔者利用不同盐度和pH胁迫下对长肋日月贝呼吸排泄的影响进行研究,旨在为长肋日月贝的养殖管理提供可靠数据,对该贝的养殖可行性进行评估,并为其生理生态学研究提供参考资料。
1 材料与方法
1.1 材料
试验用长肋日月贝采自海南三亚海域,运至陵水县新村镇试验基地并暂养。挑选活力好、无损伤的个体作为试验对象。用毛刷清洗贝壳表面的污垢,分成壳长约5.0 cm和6.0 cm两组。暂养期间日换水1次,换水量为30%,投喂饵料为亚心形扁藻(Platymanassubcordiformis)和球等鞭金藻(Isochrysisgalbana),24 h连续充气,暂养用海水为经过沉淀和砂滤及棉袋过滤的干净海水,盐度为33,pH为8.58,暂养时间为4 d。
1.2 试验方法
盐度试验:设置5个盐度梯度,分别为16、21、26、31、36。利用经过曝气的地下井水和海盐对海水进行调节,在盐度调节期间,每天盐度调节量为2,待调节到所需盐度并稳定3 d后进行试验。
pH试验:设置5个pH梯度,分别为6、7、8、9、10。分别利用NaOH溶液和HCl溶液对海水进行pH调节,先将海水pH调试至相应梯度,然后将试验贝放入并开始试验。
试验海水与暂养海水相同,温度为25.5 ℃。将长肋日月贝放入2.5 L呼吸瓶内进行呼吸排泄并密封;壳长约5.0 cm的为A组,每瓶放4个长肋日月贝,约6.0 cm的为B组,每瓶放3个长肋日月贝,并设无贝的空白对照组,以上均设置3个平行;呼吸持续时间为4 h。采用HACH-HQ30d精密仪器测量溶解氧,利用靛酚蓝分光光度法[15-16]测定海水中氨氮含量。
呼吸时间以所有试验贝放入试验水体张壳后开始计算,试验结束后测量长肋日月贝的壳长、壳高、壳宽、体质量,然后将软体部取出并放于80 ℃烘箱中烘至干质量不再减少,最后称量软体部干质量,两种规格长肋日月贝的生物学数据见表1。

表1 两种规格长肋日月贝的生物学数据
1.3 计算方法
长肋日月贝的耗氧率和排氨率公式如下:
耗氧率/mg·(h·g)-1=(D0-Dt)·V/(t·m)
排氨率/mg·(h·g)-1=(N0-Nt)·V/(t·m)
氧氮比=耗氧率/排氨率
式中,D0、Dt和N0、Nt分别为试验结束时空白对照组和试验组的溶解氧质量浓度和氨氮质量浓度(mg/L),V为试验水体的体积(L),t为呼吸持续时间(h),m为试验贝的软体部干质量(g)。
1.4 数据处理
用SPSS 19.0和Excel 2010统计软件对试验所得数据进行统计分析并作图。
2 结 果
2.1 盐度对长肋日月贝耗氧率和排氨率的影响
5个盐度梯度中,盐度为16~21时两种规格长肋日月贝的耗氧率均随盐度升高而升高;盐度为21~36时则随盐度升高而下降,A、B两组的最高耗氧率分别为3.315 mg/(h·g)和2.501 mg/(h·g),方差分析表明,盐度对长肋日月贝耗氧率的影响不显著(P>0.05),当盐度变化较大时,对长肋日月贝耗氧率的影响显著(P<0.05)(图1)。
各盐度梯度下长肋日月贝的排氨率见图2,其变化趋势与耗氧率的变化趋势一致,A、B组的单位质量排氨率最高点为盐度21时,排氨率分别为0.217 mg/(h·g)和0.166 mg/(h·g),盐度对长肋日月贝排氨率的影响不显著(P>0.05),但当盐度变化较大时影响显著(P<0.05)。A组的排氨率高于B组,规格对排氨率的影响显著(P<0.05)。

图1 不同盐度对长肋日月贝耗氧率的影响

图2 不同盐度对长肋日月贝排氨率的影响
2.2 pH对长肋日月贝耗氧率和排氨率的影响
当pH为6~9时,耗氧量呈上升趋势,pH为9~10时则表现为下降趋势,两组长肋日月贝的最高耗氧量分别为3.435 mg/(h·g)和2.624 mg/(h·g),方差分析得出pH对长肋日月贝的耗氧率影响不显著(P>0.05),pH变化较大的情况下则影响显著(P<0.05)(图3)。

图3 不同pH对长肋日月贝耗氧率的影响
pH对两种规格长肋日月贝的排氨率的影响见图4,当pH为6~9时,其排氨率与pH为正相关,而pH为9~10时则相反,A组和B组的最高排氨率分别为241.561 mg/(h·g),和200.765 mg/(h·g),方差分析表明,pH对A组长肋日月贝的排氨率影响不显著(P>0.05),但对B组影响显著(P<0.05)。同一pH梯度下,A组耗氧率比B组高,方差分析表明规格对长肋日月贝的耗氧率呈显著影响(P<0.05)。
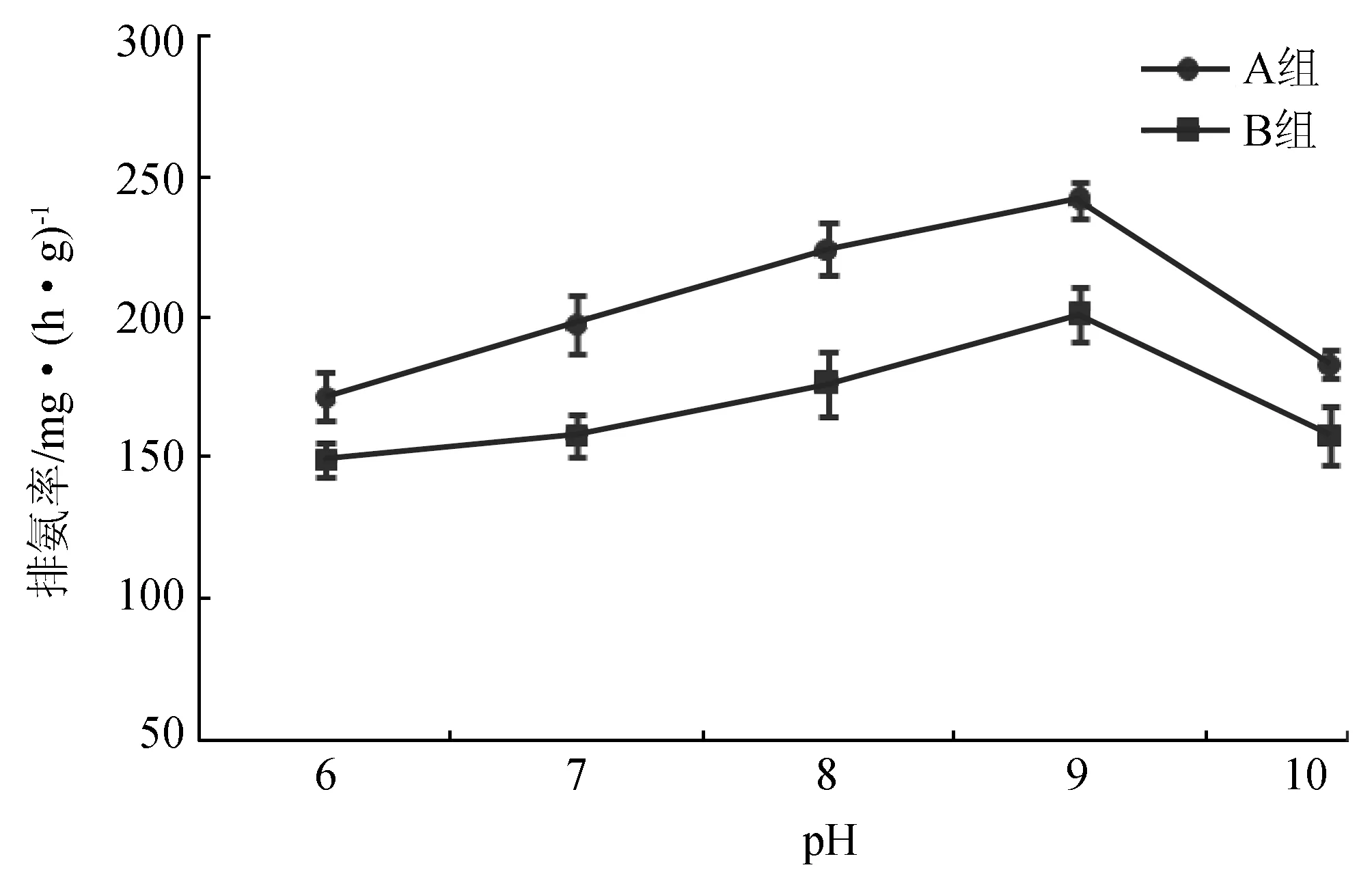
图4 不同pH对长肋日月贝排氨率的影响
2.3 盐度和pH对不同规格长肋日月贝的氧氮比
随着盐度的升高,两组长肋日月贝的氧氮比均为缓慢变大的趋势(表2)。当pH为6~9时,B组表现为随pH升高而增大,A组则不明显,至pH为10时则出现骤降。长肋日月贝的个体规格对氧氮比的影响不明显。

表2 两种规格的长肋日月贝在不同盐度和pH下的氧氮比
3 讨 论
3.1 不同氨氮测试方法的差异
水中的氨氮含量是指NH4+、NH3·H2O和NH3的总含量,均为无机氮[17],目前对水样中进行氨氮检测的常用方法主要次溴酸盐氧化法、纳氏试剂法和本研究中采用的靛酚蓝分光光度法等。次溴酸盐氧化法是最常用的水质检测方法,但对海水中氨氮的测定存在较多问题,如干扰因素多、标准系列相关性不好、测定样品中出现混浊等[17];纳氏试剂法是一种经典的氨氮检测方法,同样容易出现水体变混浊的情况,影响检测的准确度[18]。因此,以上两种方法对准确检测海水中氨氮含量有一定的难度。靛酚蓝分光光度法的出现及不断改进为海水中氨氮含量的测定提供了一个新的途径,目前采用该方法的相关能量学报道也逐渐出现[19-20]。经过改良的靛酚蓝分光光度法其回收率可超过96%,相对标准偏差为3.45%~5.1%,灵敏度与精度均较高,且操作方便,反应迅速,是适用于海水中氨氮含量测定的一种稳定方法[15-16]。
3.2 盐度对长肋日月贝耗氧率和排氨率的影响
盐度是海水养殖中的重要理化指标,对海洋生物的生长与存活有巨大影响,盐度过高或过低均对其产生消极影响,其耗氧率和排氨率会出现规律性变化。Pillai等[21]发现将新对虾(Metapenaeusmonoceros)从其适应的盐度中转移到另一低盐度水体,排氨率显著升高,而高盐度(35)的耗氧率则比中等盐度(20~25)低;而栉孔扇贝(Chlamysfarreri)在海水盐度变低后,其耗氧率和排氨率呈现为上升趋势[22]。这可能与机体在调节渗透压时需要消耗能量有关,当海洋生物生活在其适应的盐度条件中,该盐度正处于其等渗点,机体即消耗较少能量便可维持其正常的生理活动,当环境中盐度发生变化,则需要消耗更多的能量来适应,从而引起耗氧率和排氨率的升高[23]。合浦珠母贝(Pinctadafucata)在盐度为21时,其耗氧率和排氨率达到最高值[19],这与本试验的结果一致;同时,青蛤(Cyclinasinensis)在盐度为20时耗氧率和排氨率也达到最高[24],与本试验结果相近,说明以上贝类在该盐度范围需要较多的能量去维持其正常生理活动。不同物种其最高耗氧率和排氨率出现的盐度也不同,如毛蚶(Scapharcasubcrenata)稚贝[25]、皱肋文蛤(Meretrixlyrata)[26]的分别为30和23。长肋日月贝在盐度为16时耗氧率和排氨率最低,观察发现其双壳紧闭,将机体处于低强度生理活动的状态,该现象在其他贝类也有出现[27-30]。
3.3 pH对长肋日月贝耗氧量和排氨率的影响
水生生物的生理活动受pH的影响,黑唇鲍(Haliotisrubra)在pH低于7的水体中,耗氧率和排氨率明显偏低[31]。同样,欧洲食用牡蛎(Ostreaedulis)、长牡蛎(Crassostreagigas)和紫贻贝(Mytilusedulis)在pH低于7时,表型为生长速度慢,死亡率高[32]。本试验中pH为6时,长肋日月贝的耗氧率和排氨率显著低于pH为9时,可见低pH极不利于贝类的生长和存活。一般贝类耗氧率和排氨率峰值会出现在pH为8~9,如尖紫蛤(Soletellinaacuta)[33]的峰值出现在pH为8.4,缢蛏(Sinonovaculaconstricta)[34]的为pH 8和9,而长肋日月贝的峰值出现9,这表明其峰值可能处于pH为8.5~9.0。同时,pH过高对贝类的正常生理活动也不利,尖紫蛤在pH为9.4时其耗氧率和排氨率出现突然骤降[33],这与本研究结果相似。
3.4 体质量对长肋日月贝耗氧量和排氨率的影响
通常贝类的体质量越大,相同环境条件下其耗氧率和排氨率则越低,该规律在多数贝类如波纹巴非蛤(Paphiaundulate)[35]、泥螺(Bullactaexarata)[36]和方斑东风螺(Babyloniaareolata)[37]等均有出现。贝类单位体质量耗氧率或排氨率(R)与肉干质量(m)之间的关系可用以下公式表示:R=am-b[38](其中,a为单位肉干质量的耗氧率或排氨率,b为体质量指数),可见体质量与单位体质量的耗氧率和排氨率呈负相关。本研究发现在梯度水平相同的情况下,长肋日月贝体质量越大其耗氧率和排氨率越小,与该规律一致。
3.5 长肋日月贝氧氮比变化与基础代谢
氧氮比即氧和氮的比值,是动物基础代谢的重要生理指标,与蛋白质、脂肪和碳水化合物的消耗分解比例相关[39]。本研究发现氧氮比随盐度的上升而增大,说明在盐度较高的情况下,长肋日月贝主要消耗脂肪与碳水化合物来维持正常的生理代谢,而低盐度情况下则需消耗更多的蛋白质来进行渗透压的调节。长肋日月贝在pH为6~9时氧氮比为缓慢上升的趋势,然而pH为10时突然下降,表明需要消耗更多的能量来适应环境,这与机体在受到外界环境胁迫有关[40]。
总之,一定范围内的盐度、pH对长肋日月贝的耗氧率和排氨率总体影响较小,说明长肋日月贝能够较好地适应浅海或海湾等容易受雨水和人为因素影响的养殖环境,但需要作进一步的验证。
[1] 王祯瑞.中国动物志[M].北京:科学出版社,2002:166-173.
[2] 朱彩艳,王雨,张殿昌,等.长肋日月贝的营养成分分析及评价[J].海洋科学,2011,35(3):87-91.
[3] 王雨,叶乐,杨其彬,等.长肋日月贝个体发生观察及人工育苗初步试验[J].南方水产,2009,5(1):37-41.
[4] 付玉,颜云榕,卢伙胜,等.北部湾长肋日月贝的生物学性状与资源时空分布[J].水产学报,2012,36(11):1694-1705.
[5] Belda C A,Del Norte A G C.Notes on the induced spawning and larval rearing of the Asian moon scallop,Amusiumpleuronectes(Linne),in the laboratory [J].Aquaculture,1988,72(1/2):173-179.
[6] Mahidol C,Na-Nakorn U,Sukmanomon S,et al.Mitochondrial DNA diversity of the Asian moon scallop,Amusiumpleuronectes(Pectinidae),in Thailand[J].Marine Biotechnology,2007,9(3):352-359.
[7] Del Norte A G C.Aspects of the growth,recruitment,mortality and reproduction of the scallopAmusiumpleuronectes(Linne) in the Lingayen Gulf,Philippines [J].Ophelia,1988,29(2):153-168.
[8] Hao Z L,Tang X J,Ding J,et al.Effect of high temperature on survival,oxygen consumption,behavior,ammonia-N excretion,and related immune indicators of the Japanese scallopMizuhopectenyessoensis[J].Aquaculture International,2014,22(6):1863-1876.
[9] 冀德伟,陈琛,李婷婷,等.盐度胁迫对泥蚶能量代谢的影响[J].水产科学,2016,35(5):504-510.
[10] Barbieri E.Effects of zinc and cadmium on oxygen consumption and ammonium excretion in pink shrimp (Farfantepenaeuspaulensis,Pérez-Farfante,1967,Crustacea) [J].Ecotoxicology,2009,18(3):312-318.
[11] Cao F,Wang H.Effects of salinity and body mass on oxygen consumption and ammonia excretion of mudskipperBoleophthalmuspectinirostris[J].Chinese Journal of Oceanology and Limnology,2015,33(1):92-98.
[12] 杨莹,李二超,姜雪芹,等.瓦氏黄颡鱼幼鱼标准代谢的初步研究[J].水产科学,2011,30(4):192-197.
[13] 李加儿,刘士瑞,区又君,等.花尾胡椒鲷幼鱼的呼吸和排泄代谢[J].南方水产,2009,5(2):34-38.
[14] 唐贤明,田景波,隋曌,等.盐度对大菱鲆幼鱼耗氧率和排氨率的影响[J].南方水产,2006,2(4):54-58.
[15] 魏海峰,刘长发,张俊新.靛酚蓝法测定水中氨氮方法的改进[J].实验室研究与探索,2013,32(7):17-19.
[16] 王锋,李玉环.靛酚蓝光度法测定海水中的氨型氮[J].光谱实验室,2002,19(5):631-633.
[17] 吕晓洁,张志刚.海水次溴酸盐氧化法测定氨氮存在的几个问题及解决[G]//中国环境科学学会.中国环境科学学会学术年会优秀论文集(上卷),北京:中国环境科学学会,2007:474-475.
[18] 陈佳荣.水化学实验[M].北京:中国农业出版社,1996.
[19] 刘建业,喻达辉,李金碧.盐度和pH对合浦珠母贝(Pinctadafucata)耗氧率和排氨率的影响[J].海洋与湖沼,2011,42(4):603-607.
[20] 成书营,黄桂菊,潘俐玲.盐度对企鹅珍珠贝耗氧率和排氨率的影响[J].广东农业科学,2012,39(16):135-137.
[21] Pillai B R,Diwan A D.Effects of acute salinity stress on oxygen consumption and ammonia excretion rates of the marine shrimpMetapenaeusmonoceros[J].Journal of Crustacean Biology,2002,22(1):45-52.
[22] Yang H,Wang P,Zhang T,et al.Effects of reduced salinity on oxygen consumption and ammonia excretion ofChlamysfarreri[J].Chinese Journal of Oceanology and Limnology,1998,17(3):207-211.
[23] 张媛,方建光,毛玉泽,等.温度和盐度对橄榄蚶耗氧率和排氨率的影响[J].中国水产科学,2007,14(4):690-694.
[24] 金春华.温度和盐度对青蛤耗氧率和排氨率的影响[J].丽水学院学报,2005,27(2):46-51.
[25] 沈伟良,尤仲杰,施祥元.不同规格及不同盐度下毛蚶稚贝耗氧率和排氨率的研究[J].海洋水产研究,2008,29(2):53-56.
[26] 栗志民,刘志刚,徐法军,等.体重、温度和盐度对皱肋文蛤耗氧率和排氨率的影响[J].海洋科学进展,2011,29(4):512-520.
[27] 唐保军,尹飞,归从时.海水盐度对紫血蛤耗氧率和排氨率的影响[J].台湾海峡,2012,31(1):89-94.
[28] Navarro J M.The effects of salinity on the physiological ecology ofChoromytiluschorus(Molina,1782) (Bivalvia:Mytilidae) [J].Journal of Experimental Marine Biology and Ecology,1988,122(1):19-33.
[29] Djangmah J S,Shumway S E,Davenport J.Effects of fluctuating salinity on the behaviour of the west African blood clamAnadarasenilisand on the osmotic pressure and ionic concentrations of the haemolymph [J].Marine Biology,1979,50(3):209-213.
[30] Shumway S E.Oxygen consumption in oysters:an overview [J].Marine Biology Letters,1982,3(1):1-23.
[31] Harris J O,Maguire G B,Edwards S J,et al.Effects of pH on growth rate,oxygen consumption rate,and histopathology of gill and kidney tissue for juvenile greenlip abalone,HaliotislaevigataDonovan and blacklip abalone,Haliotisrubraleach[J].Journal of Shellfish Research,1999(18):611-619.
[32] Bamber R N.The effects of acidic seawater on three species of lamellibranch mollusc[J].Ecology,1990,143(3):181-191.
[33] 黄洋,黄海立,林国游,等.盐度、pH 和规格对尖紫蛤(Soletellinaacuta)耗氧率和排氨率的影响[J].海洋与湖沼,2013,44(1):120-125.
[34] 范德朋,潘鲁青,董双林,等.盐度和pH 对缢蛏耗氧率及排氨率的影响[J].中国水产科学,2002,9(3):234-238.
[35] 李俊辉,刘红礼,杜晓东,等.盐度和规格对波纹巴非蛤Paphiaundulate耗氧率和排氨率的影响[J].海洋学研究,2011,29(4):95-99.
[36] 吴文广,张继红,方建光,等.盐度对不同规格泥螺耗氧率和排氨率的影响[J].生态学杂志,2013,32(9):2457-2461.
[37] 刘建勇,邓兴朝,绍杰.体重和温度对方斑东风螺耗氧率和排氨率的影响[J].中国水产科学,2005,12(3):239-244.
[38] Luo J,Liu C,Li F, et al.Effects of salinity and body weight on oxygen consumption rate and ammonia excretion rate ofHemifusustuba[J].Marine Sciences,2008,32(5):46-50.
[39] Mayzaud P.Respiration and nitrogen excretion of zooplankton:Ⅱ.studies of the metabolic characteristics of starved animials[J].Marine Biology,1973,21(1):19-28.
[40] Widdows J.Combined effect of body size,food concentration and season on the physiology ofMytillusedulis[J].Journal of Marine Biology Assoc UK,1978,58(2):109-124.
