长海县筏养虾夷扇贝大规模死亡原因的初步研究
2018-03-27于佐安李大成王笑月林杉杉付成东刘项峰周遵春
于佐安,李大成,王笑月,张 明,林杉杉,付成东,刘项峰,周遵春
(辽宁省海洋水产科学研究院,应用海洋生物技术开放实验室,水产分子生物学重点实验室,辽宁 大连 116023)
2007年至今,长海县筏养虾夷扇贝(Pationopectenyessoensis)连续多年大规模死亡,尤其在2龄贝养成期的6—9月死亡率最高,导致当前该海区筏养虾夷扇贝产业以一龄贝养殖为主。与此相似,20世纪90年代,栉孔扇贝(chlamysfarreri)爆发大规模死亡[1],国内学者从病害、养殖密度、海区环境等角度对其死亡原因开展研究,张福绥等[2]认为在夏季高温期,海区饵料不足,消耗增大,养殖水体污染和病原体滋生等是直接原因;而宋微波等[3-4]认为一种球形病毒是导致栉孔扇贝大规模死亡的直接原因。另有研究表明,高温是引发养殖贝类大规模死亡的主要环境胁迫因素,高温可以对生物机体产生热胁迫,进而对生物产生多方面影响[5-8]。而夏季海区持续的高温可导致贝类的血细胞浓度、吸附能力以及吞噬活性显著降低,影响存活细胞以及死亡细胞的能力,诱发扇贝疾病生长死亡[9-13]。滕炜鸣等[14]从虾夷扇贝闭壳肌脓胞中分离出一株查氏弧菌(Vibriochagasii),经鉴定确认,查氏弧菌是导致虾夷扇贝闭壳肌出现脓胞的主要致病病原。笔者通过病毒粒子的病原学相关研究和流行病学调查结果也认为高温与虾夷扇贝大规模死亡有一定相关性,而病毒不是导致虾夷扇贝大规模死亡的主要原因[15]。总体看,扇贝大规模死亡是环境条件、病害、养殖密度、种质等多方面因素综合造成的。
长海县筏养虾夷扇贝大规模死亡虽然也和水温、病害、养殖密度、种质等多方面因素相关,但造成虾夷扇贝大规模死亡的真正病因尚未见详细系统的研究报道。笔者从与筏养虾夷扇贝大规模死亡相关的主要因素中选取水温、饥饿和病害三个因素进行初步研究分析,探讨该海域虾夷扇贝大规模死亡的病因,为防治其大规模死亡提供参考。
1 材料与方法
1.1 筏养虾夷扇贝海区调查及样品采集
1.1.1 筏养虾夷扇贝海区浮游植物丰度调查
(1)样品采集
2015年4—9月,在大长山岛小泡子村筏式虾夷扇贝养殖海区(北纬39°15′10.74″~39°15′34.02″,东经122°32′55.50″~122°34′13.26″)设定5个调查站位(图1),每月采样一次,采样水层为4 m(筏式虾夷扇贝养殖水深一般为3~5 m)。
图1 筏养虾夷扇贝海区浮游植物调查站位
(2)按照国家海洋调查规范(GB/T 12763.6-2007)[16]进行采样分析,浮游植物优势度根据其出现的频率及丰度计算[17],计算公式为:
Y=(ni/N)×fi
式中,Y为优势度,ni为第i个种的丰度,N为样品总丰度,fi为该种的站位出现频率。以Y≥0.02确定为优势种类。
1.1.2 筏养虾夷扇贝死亡数量统计和水温实时监测
1.1.2.1 虾夷扇贝死亡数量和脓胞数量统计
2015年4—9月,每月取样一次,统计虾夷扇贝死亡情况,每个站位取3笼养殖扇贝(2015年4月分苗,共1200枚)进行死亡率统计分析。累计死亡率分析参考文献[18]的方法,月平均死亡率的分析处理参考文献[19]的方法。每个站位解剖30枚扇贝,统计出现脓胞的扇贝个数。
1.1.2.2 水温实时监测
2015年4—9月,在筏养虾夷扇贝海区,利用Seabird SBE56水下温度仪进行连续不间断水温实时监测。测量周期2 h一次,测量水深3 m。
1.2 同水温条件不同单细胞饵料密度模拟养殖试验
1.2.1 试验材料和日常管理
1.2.1.1 试验材料和时间
试验用虾夷扇贝来自选育家系,在长海县小长山岛海洋岛集团所属海区进行养成。扇贝壳高5~7 cm,2016年6月21日开始在辽宁省海洋水产科学研究院海水引育种中心暂养,8月2日试验结束,各试验组试验周期约30 d。
1.2.1.2 水温控制和日常管理
2016年6月17日引进试验用虾夷扇贝,控制暂养池水温在14 ℃,每日升温1 ℃。水温升至17 ℃,从暂养池选择健康有活力的虾夷扇贝进行第一组温度梯度试验,水温升至19 ℃和22 ℃时,进行第二和第三组温度梯度试验。为保证试验虾夷扇贝处于相同饵料水平,暂养期间不投饵。
1.2.2 试验组别设置
制冷机循环水设备控温的7 ℃冷水和常温海水混合,配置14 ℃、18 ℃和22 ℃海水,由于室温和水体较小原因,日均水温有一定波动,3个温度区间试验组为14~17 ℃、18~20 ℃和22~24 ℃;每个温度区间条件下,设置3个单胞藻饵料密度投喂组:0 个/L(饥饿组)、107个/L组和108个/L组。每组设3个平行,每个平行15枚扇贝。
试验在80 L的圆筒中进行,试验基地培养饵料密度为1.6×106个/mL,根据饵料投喂密度每个试验组每次分别投喂新月菱形藻(Nitzschiaclosterium)0 mL,500 mL和5 L。投喂3次/d,换水1次/d。
1.3 数据处理
试验过程中统计各试验组每天虾夷扇贝死亡个数,解剖扇贝,统计闭壳肌中有脓胞出现扇贝个数。
2 结果与分析
2.1 筏养虾夷扇贝海区浮游植物丰度
2.1.1 浮游植物种群构成
2015年4—9月,筏养虾夷扇贝海区浮游植物优势种以硅藻类为主(表1)。4月和5月的优势种为柔弱几内亚藻(Guinardiadelicatula)和具槽直链藻(Paraliasulcata)。4月两种藻类优势度为0.88和0.06;5月两种藻类优势度为0.85和0.04;6月优势种为中肋骨条藻(Skeletonemacostatum)和丹麦细柱藻(Leptocylindrusdanicus),优势度为0.28和0.17;7月优势种为中肋骨条藻、丹麦细柱藻、具槽直链藻、海洋原甲藻(Prorocentrummicans)、菱形藻(Nitzschia)和圆柱角毛藻(Chaetocerosteres),优势度为0.33、0.15、0.13、0.11、0.09和0.03;8月优势种为丹麦细柱藻和短角弯角藻(Eucampiazodiacus),优势度为0.71和0.2;9月优势种为柔弱伪菱形藻(Pseudonitzschiadelicatissima)、旋链角毛藻(C.curvisetus)和具槽直链藻,优势度为0.13、0.09和0.06。
2.1.2 浮游植物丰度
2015年4—9月,调查海区浮游植物丰度月度变化趋势为4月最高,密度为16.65×105个/L。 5—7月连续下降,5月为8.45×105个/L,6月为4.14×105个/L,7月为2.27×105个/L,8月数量有所回升,达到8.18×105个/L,9月浮游植物丰度又大幅下降,仅为0.6×105个/L(图2)。
2.2 扇贝死亡数量和水温监测
2.2.1 虾夷扇贝死亡数量统计
2015年4—9月,调查海域的二龄虾夷扇贝死亡率和脓胞出现个数的统计结果见表2。4月,虾夷扇贝以每层20枚扇贝(壳高5~6 cm)的密度装入20层板养殖笼,9月底统计时累计死亡率达到91%;利用四格表卡方检验各月平均死亡率, 6—7月月平均死亡率最高,达到57.9%(P<0.05),出现脓胞个数也最多;8—9月月死亡率为53.7%。因此死亡最高峰发生在6—7月,但8—9月高温期扇贝死亡率也很高。

图2 2015年4—9月大长山岛虾夷扇贝浮筏养殖海区浮游植物丰度月度变化
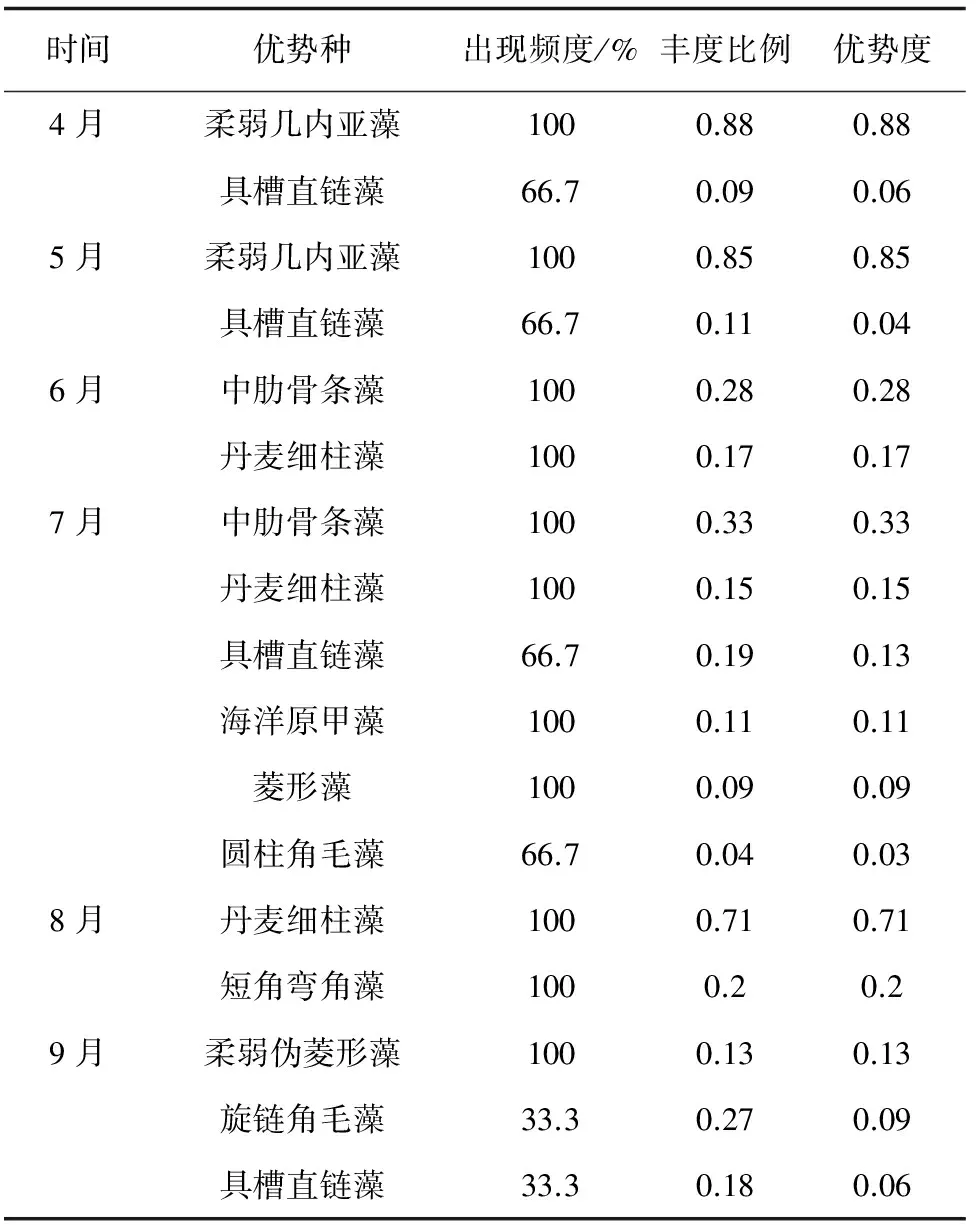
表1 2015年4—9月浮筏虾夷扇贝海区浮游植物优势种

表2 2015年4—9月筏养虾夷扇贝死亡情况统计
注:表格中同列肩标相同表示差异不显著(P>0.05).
2.2.2 海区水温实时变化
2015年4—9月,调查海区水温变化见图3、图4。
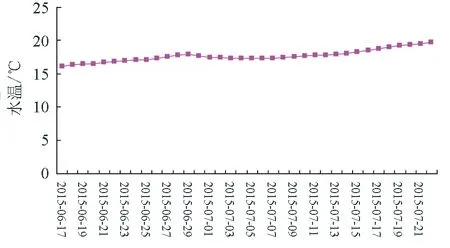
图3 2015年6月17日—7月22日水温变化
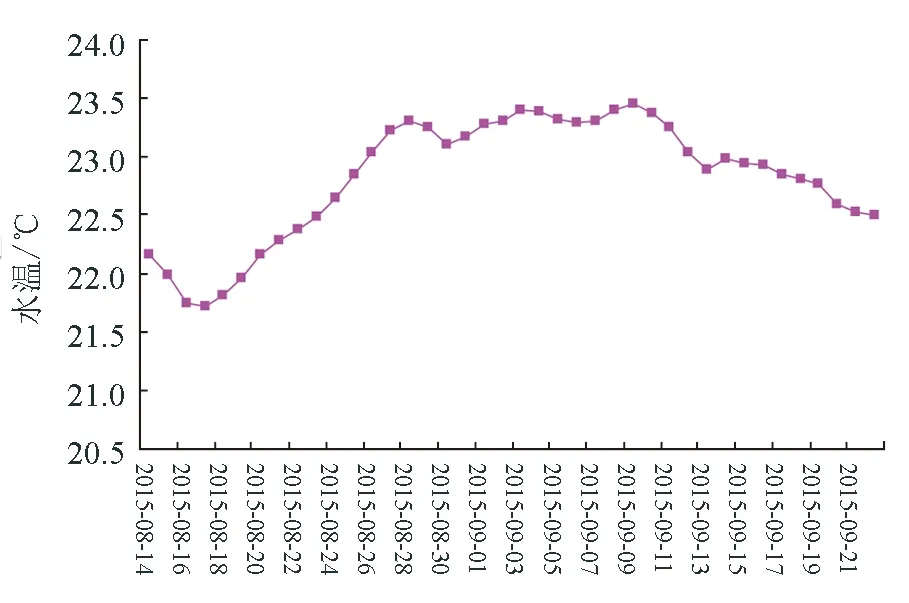
图4 2015年8月14日—9月22日水温变化
6—7月,日水温变化在16.29~20.19 ℃,月平均水温18.06 ℃;8—9月,日水温变化在22.09~23.78 ℃,日平均水温超过23 ℃的天数达到23 d。
2.3 室内不同温度不同饵料密度养殖试验结果
18~20 ℃温度区间饥饿组的死亡率及脓胞出现比例最高(P<0.05);在任一饵料密度组,3个温度区间组的虾夷扇贝死亡率均有显著差异,18~20 ℃温度区间组死亡率最高,22~24 ℃温度区间组死亡率次之,14~17 ℃温度区间组死亡率最低。脓胞出现比例在18~20 ℃温度区间最高,但其他两个温度区间组脓胞出现比例差异不显著;在18~20 ℃温度区间,饥饿组虾夷扇贝死亡率和脓胞出现比例均为最高,但在2个温度区间,3个单细胞饵料(饥饿、107个/L组和108个/L组),死亡率和脓胞出现比例无显著性差异(表3)。
统计模拟养殖试验进程中3个温度区间组每日虾夷扇贝总死亡个数(图5),进行线性回归分析,发现14~17 ℃温度区间和18~20 ℃温度区间虾夷扇贝的死亡规律比较相似,相关系数分别为0.881和0.8795,而22~24 ℃温度区间虾夷扇贝死亡规律和另外2个温度区间组不相同,相关系数为0.9884。表明死亡原因可能有所差异,22~24 ℃温度区间出现的脓胞比例和14~17 ℃温度区间虽然没有显著性差异,但此时脓胞和14~17 ℃温度区间的相比更加明显,很可能是病灶发展的结果,并非新产生的脓胞。

表3 室内模拟养殖不同试验组虾夷扇贝死亡情况和脓胞出现个数统计
注:表格中同行肩标相同小写字母表示差异不显著(P>0.05).
3 讨 论
3.1 水温对虾夷扇贝大规模死亡的影响
研究显示,在环境因子良好的情况下,海水温度过高可导致栉孔扇贝死亡,且扇贝死亡率随着高温刺激时间的延长呈上升趋势[20-21]。日本虾夷扇贝在20世纪70年代曾发生过大规模死亡,夏季的高水温,使已经出现生理障碍的扇贝对环境变化的抵抗力减弱,同样被认为是大规模死亡的主要原因之一[22]。本研究探讨水温和虾夷扇贝大规模死亡的联系,根据室内模拟养殖试验结果和海区流行病学调查结果,高水温不是导致虾夷扇贝开始死亡的直接原因,17 ℃左右的水温可能是虾夷扇贝开始死亡的起始温度,当水温高于17 ℃,虾夷扇贝月平均死亡率开始大幅提高[15]。本研究中2015年6月17日—7月22日的虾夷扇贝月平均死亡率最高,此时海水温度为16.29~20.19 ℃,室内模拟养殖试验中18~20 ℃区间组死亡率同样最高,虾夷扇贝死亡率最高时期的水温未超出其适宜生长水温范围,表明高水温不是6—7月虾夷扇贝大规模死亡的原因;进入8—9月,海区水温达到最高值,但脓胞出现比例却显著低于6—7月,但由于日平均水温超过23 ℃ 的时间连续达到23 d,虾夷扇贝月平均死亡率达到53.7%,仅略低于6—7月的月平均死亡率;室内22~24 ℃温度区间组的脓胞出现比例也低于18~20 ℃温度区间组,但由于高温刺激,虾夷扇贝的死亡率也仅略低于18~20 ℃温度区间组。说明高水温尤其是连续高温刺激加剧了虾夷扇贝大规模死亡。总体而言,长海县水温变化和虾夷扇贝大规模死亡密切相关,17 ℃左右的水温是虾夷扇贝开始大规模死亡的重要节点,而高水温不是导致虾夷扇贝开始大规模死亡的原因,但在8—9月期间加剧了扇贝死亡。
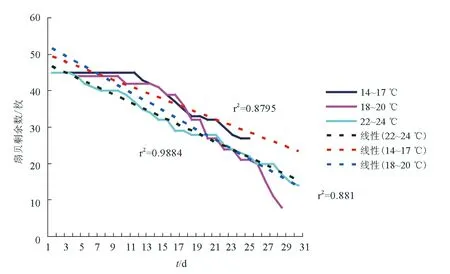
图5 3个温度区间组虾夷扇贝存活扇贝个数趋势比较
3.2 饥饿对虾夷扇贝大规模死亡的影响
Navarro等[23-24]认为,大规模养殖浮筏会形成一定屏障效益,导致养殖区饵料供应不足,造成扇贝的营养障碍机体能量收支失衡,引发抗性下降,易受病原侵入。长海县虾夷扇贝浮筏养殖面积最高达到2.7×104hm2,截至2016年仍有1.0×104hm2,且筏间距越来越小,筏身长度越来越长,每层扇贝养殖数量也很多,同时海区牡蛎养殖和海湾扇贝养殖规模越来越大,因此对海区环境承载力的压力仍然很大。这可能会导致海区饵料严重不足,扇贝体质变弱,引发病害。
本试验从调查海区浮游植物丰度月度变化入手,发现海区浮游植物主要优势种主要为硅藻,丰度变化则比较明显,4月最高,随后急剧下降,5—7月份逐步降低,8月略有上升,这与付成东等[25]对长海县浮游植物丰度的调查结果一致。5—7月,虾夷扇贝排卵后需要大量摄食补充营养,也是虾夷扇贝生长最快,滤水率最高的时期,此时浮游植物丰度的大幅下降可能会导致扇贝饵料不足。通过室内模拟养殖试验,在不同水温不同饵料条件下,18~20 ℃温度区间的饥饿组脓胞出现比例和死亡率最高,而在另外两个温度区间组,不同饵料密度组在死亡率和脓胞出现比例方面没有显著差异。这说明单纯饥饿并不能导致扇贝大规模死亡,14~17 ℃温度区间表现出差异的可能原因为此时病原刚刚开始侵袭宿主,不同饵料密度虽然使虾夷扇贝对病原的抵抗力有所不同,但在这个温度区间,病原毒力相对较弱,且刚开始感染,因此饥饿组在死亡率和脓胞出现比例方面未与另外2个饵料密度组表现出显著性差异。22~24 ℃温度区间表现出差异的可能原因是在此温度区间,扇贝的滤水率和摄食率都很低,不同饵料密度组造成的差异相对较小,而这段温度区间高水温和病害都是造成虾夷扇贝死亡的主要原因。
3.3 病害对虾夷扇贝大规模死亡的影响
近几十年,我国海洋贝类养殖规模发展迅速,养殖贝类大规模死亡问题日益严重,菲律宾蛤仔(Ruditapesphilippinarum)、贻贝(Mytilusedulis)、栉孔扇贝都因为病害问题发生过大规模死亡[26]。长海县自2007发生风暴潮以后,筏养虾夷扇贝也连续多年发生大规模死亡。根据流行病学调查,虾夷扇贝出现脓胞是死亡扇贝典型症状[15],因此本研究将脓胞作为病害发生的标记。2015年4—9月的海区调查结果显示虾夷扇贝月死亡率最高的时期是6—7月,脓胞出现个数也最高;而室内模拟养殖试验结果同样表明,虾夷扇贝死亡率最高的试验组脓胞出现比例也最高,这说明脓胞出现个数最多时,也是扇贝死亡最严重的时期,病害和扇贝死亡密切相关;由于虾夷扇贝大规模死亡具有典型的周期性(在适宜水温开始出现死亡,每年在6—9月期间1龄至2龄养成期虾夷扇贝都会发生大规模死亡)、具有典型发病症状(闭壳肌出现脓胞),并且在扇贝大规模死亡各个时期均能发现脓胞,因此病害很可能是导致虾夷扇贝大规模死亡的主要直接原因。
3.4 水温、饥饿和病害对虾夷扇贝大规模死亡的共同影响
大规模高密度养殖导致饵料不足,进一步导致虾夷扇贝对病原的抵抗力和对环境的耐受力都开始下降,在适当水温条件下,病原感染虾夷扇贝,宿主开始大规模死亡,同时由于高温胁迫,加剧了虾夷扇贝死亡。根据本研究结果,在14~17 ℃温度区间和18~20 ℃温度区间组虾夷扇贝的死亡规律比较相似,相关系数比较相近,此时病害起主要作用;在22~24 ℃温度区间,相关系数显著变大,扇贝死亡速度明显加快,虽然22~24 ℃温度区间出现的脓胞比例和14~17 ℃温度区间虽然没有显著性差异,但此时脓胞和14~17 ℃温度区间的相比更加明显,很可能是病灶发展的结果,并不是新产生的脓胞。因此,22~24 ℃温度区间组虾夷扇贝的死亡是病害和高温共同作用结果。虾夷扇贝大规模死亡是在饵料不足的条件下,病害和水温两个因素是扇贝死亡不同阶段的关键原因。6—7月,适宜水温条件下(约17 ℃),病害是虾夷扇贝大规模死亡的主要直接原因。8—9月,高水温和病害共同导致虾夷扇贝大规模死亡。
[1] 王运涛,相建海.栉孔扇贝大规模死亡的原因探讨[J].海洋与湖沼,1999,30(6):770-774.
[2] 张福绥,杨红生.山东沿岸夏季栉孔扇贝大规模死亡原因分析[J].海洋科学,1999(1):44-47.
[3] 宋微波,王崇明,王秀华,等.栉孔扇贝大规模死亡的病原研究新进展[J].海洋科学,2001,25(12):23-26.
[4] 王崇明,王秀华,艾海新,等.栉孔扇贝大规模死亡致病病原的研究[J].水产学报,2004,28(5):547-553.
[5] Ravagnolo O,Misztal I.Effect of heat stress on nonreturn rate in Holstein cows: genetic analyses[J].Journal of Dairy Science,2002,85(11): 3092-3100.
[6] West J.Effects of heat-stress on production in dairy cattle[J].Journal of Dairy Science,2003,86(6):2131-2144.
[7] Mashaly M,Hendricks G,Kalama M, et al.Effect of heat stress on production parameters and immune responses of commercial laying hens[J].Poultry Science,2004,83(6):889-894.
[8] Starkie R,Hargreaves M,Rolland J, et al.Heat stress,cytokines,and the immune response to exercise[J].Brain,Behavor,and Immunity,2005,19(5):404-412.
[9] 陆彤霞,王国良,尤仲杰,等.我国海洋养殖贝类病害研究概况及防治对策[J].浙江海洋学院学报,2002,21(2):154-159.
[10] Pampanin D M,Camus L,Gomiero A,et al.Susceptibility to oxidative stress of mussels (Mytilusgalloprovincialis) in the Venice Lagoon (Italy)[J].Marine Pollution Bulletin,2005,50(12):1548-1557.
[11] Matozzo V,Gallo C,Marinet M,et al.Effects of temperature on cellular and biochemical parameters in the crabCarcinusaestuarii(Crustacea,Decapoda)[J].Marine Environmental Research,2011,71(5):351-356.
[12] Gagnaire B,Frouin H,Moreau K,et al.Effects of temperature and salinity on haemocyte activities of the Pacific oyster,CrassostreaGigas(Thunberg)[J].Fish & Shellfish Immunology,2006,20(4):536-547.
[13] Monari M,Monari M,Foschi J,et al.Effects of salinity on the clamChameleagallinahaemocytes.Part Ⅱ: superoxide dismutase response[J].Marine Biology,2007,151(3):1059-1068.
[14] 滕炜鸣,李文姬,张明,等.虾夷扇贝脓胞病病原的分离、鉴定与致病性[J].水产学报,2012,36(6):937-943.
[15] 于佐安,李文姬,张明,等.虾夷扇贝大规模死亡的流行病学研究及可能致病因子的探讨[J].中国海洋大学学报:自然科学版,2013,43(6):52-57.
[16] 国家海洋局.海洋调查规范[M].北京: 海洋出版社,1975.
[17] 孙儒泳.动物生态学原理[M].北京: 北京师范大学出版社,1992:360-361.
[18] Geng G Y.Epidemiology [M].Beijing: People’s Hygiene Press,1989: 5-22.
[19] Angusdsa A.Survey toolbox for livestock diseases [M].Australia:ACLAR Monograph No 54,1999:26-27.
[20] 过峰,孙耀.日照石臼扇贝养殖水域夏季环境调查分析[J].海洋水产研究,1999,20(1):45-51.
[21] 柳中传,吴建芹,金福江.栉孔扇贝死亡原因及预防措施[J].海洋科学,1992,(5):9-10.
[22] 李文姬,谭克非.日本解决虾夷扇贝大规模死亡的启示[J].水产科学,2009,28(10):609-612.
[23] Navarro E,Iglesias I P,Camacho A P,et al.The physiological energetics of mussels(Mytilusgalloprovincials lmk) from different cultivation rafts in Ria de Arosa(Galicia,N.W.Spain)[J].Aquaculture,1991(94):197-212.
[24] Kennedy V S.The ecological role of eastern oyster,Crassostreavirginica,with remarks on disease[J].Journal of Shellfish Research,1996,15(1):177-183.
[25] 付成东,李大成,于佐安,等.长海县西北部海域浮游植物种类及密度的变化[J].水产科学,2012,31(7):408-412.
[26] 张国范,李霞,薛真福.我国养殖贝类大规模死亡的原因分析及防治对策[J].中国水产,1999(9):34-38.
