舟山近岸海域春秋季鱼类种类组成及其数量时空分布
2018-03-26邓小艳俞存根水玉跃翁良才丁建伟张平许永久郑基谢旭苗露
邓小艳, 俞存根, *, 水玉跃, 翁良才, 丁建伟, 张平, 许永久, 郑基, 谢旭, 苗露
舟山近岸海域春秋季鱼类种类组成及其数量时空分布
邓小艳1, 俞存根1, *, 水玉跃2, 翁良才2, 丁建伟2, 张平1, 许永久1, 郑基1, 谢旭1, 苗露1
1. 浙江海洋大学水产学院, 舟山 316022 2. 舟山市海洋与渔业局, 舟山 316000
根据2015年11月(秋季)、2016年5月(春季)在舟山近岸海域开展的渔业资源调查中所获得的鱼类资源资料, 对该海域鱼类的种类组成、数量分布、优势种变化及其影响因素等进行了研究。结果如下: 根据本次调查所获得的样品, 经分析共鉴定出鱼类种类81种, 隶属于12目39科63属; 春季和秋季鱼类重量密度分别为10.36 kg·km-2和98.01 kg·km-2, 尾数密度分别为26.29×103ind·km-2和33.30 ind·km-2; 两个季节鱼类资源密度的平面分布特征呈相同规律, 都是东部外侧海域高于122.3°E以西海域; 不同季节优势种种类不同, 春季的优势种为六丝钝尾虾虎鱼, 秋季优势种为龙头鱼, 均为小型低值鱼类; 鱼类春季和秋季总种类数变化较为稳定, 但种类数的季节变化较大, 春季以沿岸浅海附近种类较多, 秋季以调查海域东北部较多; 水深对鱼类资源密度及种类数分布影响明显; ABC曲线显示, 舟山近岸海域的鱼类群落处于严重干扰的状态。与以往调查结果相比, 舟山渔场鱼类种类组成、群落结构等都发生了较大的变化。
鱼类; 种类组成; 数量分布; 资源密度; ABC曲线; 舟山近岸海域
1 前言
舟山近岸海域沿岸线曲折, 岛屿密布, 台湾暖流、沿岸流在此交汇, 生境类型丰富[1], 是多种经济渔业生物产卵场和索饵场[2], 是名副其实的“鱼的故乡”[3]。但随着捕捞强度的增加以及环境污染加剧, 使该渔场渔业资源遭到了严重破坏, 海洋生态恶化日趋严重, 出现传统渔场、渔汛消失, 捕捞对象更替, 经济价值高、个体大的鱼类群落逐渐被小型低值鱼类群落所取代的现象, 渔业资源生物链以及平衡被打破, 鱼类群落结构已经发生了一系变化[4-5]。
目前, 有关对东海鱼类群落结构的研究已有很多[6-9], 对舟山渔场鱼类资源的报道也有不少, 但主要集中于对某一海域的鱼类资源进行研究[10-12], 而针对以整个舟山近岸海域鱼类资源为主体的种类组成和资源分布的研究尚不多见, 并且这一区域是舟山渔场某些鱼类重要的产卵场和索饵场。因此, 研究该海域鱼类种类组成、优势种、资源密度平面分布等有重要的科学意义。本文以2015年11月(秋季)、2016年5月(春季)在舟山近岸海域开展的渔业资源调查中所获得的鱼类资源资料为基础, 分析了该海域鱼类的种类组成、资源分布、优势种变化等, 并与以往该海域调查资料进行对比分析, 目的是为了了解舟山近岸海域鱼类资源的现状和分布特点, 以及优势种种群演替规律等, 为保护舟山近岸海域的鱼类资源, 修复生态环境, 建立可持续发展的海洋生态系统及科学合理的渔业管理体系提供科学依据。
2 材料与方法
2.1 采样方法
本文所用数据取自2015年11月(秋季)、2016年5月(春季)“浙普渔32207号”单拖网船在舟山近岸海域29°20'—31°00'N, 121°40'—123°00'E范围内实施的渔业资源调查时所获得的鱼类资源资料, 在该范围内共设80个调查站位(图1)。采样网具为底拖网, 网具规格为800目×80 mm, 每个站位拖曳时间约为1 h, 拖速约为3.5 kn。记录渔获物中鱼类的网产量, 统计每个物种的重量和尾数, 并对每个物种进行生物学测定。调查海域的水深等测定采用CTD仪与拖网调查同步进行。
2.2 数据分析
2.2.1 鱼类资源密度
根据扫海面积法[13]估算鱼类资源密度, 公式如下:
ρ=C/aq
式中:ρ为第站的鱼类资源密度(重量: kg·km-2; 尾数: 103ind·km-2);C为第站的每小时拖网渔获物中鱼类数量(重量: kg·h-1; 尾数: ind·h-1);a为第站的网具每小时扫海面积(km2·h-1), 为网口水平扩张宽度(km, 本网具为0.0107 km)×拖曳距离(km), 拖曳距离为拖网速度(km·h-1)和实际拖网时间(h)的乘积;为网具捕获率(可捕系数=1-逃逸率), 依据本拖网网具, 重量尾数逃逸率取0.5。
2.2.2 优势种
优势种的计算采用相对重要性指数, 计算公式如下:
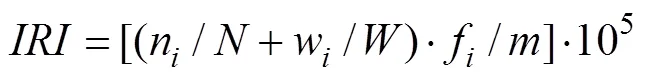
式中:n、w分别为第i种生物的个体数和生物量;分别为调查所获得总个体数和总生物量;f为第i种生物在次取样中出现的频率;为取样次数。将相对重要性指数()值大于1000者定为优势种, 数值在100—1000之间者定为常见种[10]。
2.2.3 丰度生物量比较曲线
丰度生物量比较曲线(ABC曲线)是在同一坐标系中比较生物量优势度曲线和数量优势度曲线, 通过两条曲线的分布情况来分析群落处于不同干扰状况下的特征[14], 其统计量用表示[15], 计算公式如下:
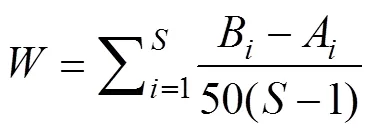
其中: S为出现的物种数, Bi、Ai分别为曲线中种类序号对应的生物量和丰度的累计百分比, 当生物量优势度曲线在丰度优势度曲线之上时, W为正, 反之为负。
为了分析调查海域鱼类资源密度以及种类数的空间分布差异, 将调查海域按水深分为15 m以浅、15—45 m、45 m以深三个水深带分别进行计算。
调查站位图及鱼类数量分布图采用Surfer 11软件绘制。数据分析处理采用Excel 2003、PRIMER 5.0软件完成。
3 结果
3.1 种类与组成
3.1.1 种类
舟山近岸海域春季和秋季调查共鉴定出鱼类81种, 隶属于12目39科63属, 其中春季58种, 隶属于11目34科49属, 秋季53种, 隶属于11目25科41属。从目级分类阶元来看, 以鲈形目种类最多, 共33种, 占鱼类总种数的40.74%; 其次是鯡形目, 共13种, 占鱼类总种数的16.05%。经济价值较高的种类有龙头鱼(Harpodon nehereus)、中华小沙丁鱼(Sardinella nymphaea)、海鳗()、鮸鱼(Miichthys miiuy)、银鲳()、棘头梅童鱼()、凤鲚()、绿鳍鱼(Chelidonichthys kumu)、黑鮟鱇(Lophiomus setigerus)、小黄鱼()、带鱼(Trichiurus haumela)等, 但仅龙头鱼的群体数量较大, 其余种类均较低(表1)。
3.1.2 组成
春季和秋季调查所获鱼类中生物量比例大于0.01%且出现率大于1.25%的鱼类有37种(表1)。其中, 六丝钝尾虾虎鱼()是春季生物量所占比例最高的种类(42.16%), 其次是银鲳(12.31%)、棘头梅童鱼(6.87%)、褐菖鲉(Sebastiscus marmoratus)(6.77%), 其余种类生物量的比例均在5%以下。秋季生物量所占比例最高的是龙头鱼, 为43.56%, 其次是黄鲫()(12.30%)、银鲳(9.28%)、鮸鱼(8.09%)、中华小沙丁鱼(8.04%)、海鳗(4.25%), 其余种类所占比例较低, 均低于3%(表1)。可见本次调查春、秋季鱼类群体数量主要集中在少数几个种类上, 大部分种类的群体数较少。
从表1还可以看出, 春季六丝钝尾虾虎鱼的出现率最高(98.75%), 其他依次为蓝园鲹(71.25%)、绿鳍鱼(61.25%)、银鲳(52.50%)等, 秋季各种类的出现率明显高于春季, 秋季龙头鱼的出现率最高(100%), 其他依次为刺冠海龙(Corythoichthys crenulatus)(82.50%)、红鳗虾虎鱼() (58.75%)、棘头梅童鱼(48.75%)等。春季和秋季生物量比例大于0.01%且出现率大于1.25%的种类分别有30种和28种, 各占春、秋季种类总数的51.72%和52.83%(表1)。

表1 舟山近岸海域春、秋季鱼类组成及优势种
注:为生物量所占比例;为尾数所占比例;为出现率。
3.2 数量分布
3.2.1 鱼类密度分布
本次调查海域春季和秋季鱼类的重量密度分别为10.36 kg·km-2和98.01 kg·km-2, 尾数密度分别为26.29×103ind·km-2和31.62×103ind·km-2。
春季, 整个调查海域鱼类的重量密度较低, 各站位的重量密度分布范围为0.23—70.43 kg·km-2, 高低相差306倍, 以东部外侧海域及东北部海域鱼类重量密度较高, 而位于122.3°E以西海域站位的重量密度均较低, 超过50%站位的重量密度在10 kg·km-2以下, 其中位于杭州湾海域站位的重量密度最低(图2)。尾数密度分布情况与重量密度分布情况类似, 以调查海域南部及杭州湾附近海域的尾数密度较低, 均在0—10×103ind·km-2范围内(图3)。重量密度和尾数密度都以46号站位最高, 分别为70.43 kg·km-2和237.31×103ind·km-2, 主要种类也都为六丝钝尾虾虎鱼。
秋季, 整个调查海域鱼类的重量密度明显高于春季, 各站位的重量密度分布范围为3.11—2562.7 kg·km-2, 高低相差824倍, 重量密度较高的站位集中在调查海域东北部的嵊泗列岛附近, 其中2号站位的重量密度最高, 有2562.7 kg·km-2, 主要由数量较多的黄鲫、银鲳、龙头鱼等鱼种构成。另外位于调查海域中部区域的衢山岛附近海域站位的重量密度也较高, 这些站位均有大量龙头鱼出现(图2)。尾数密度以调查海域东北部的2号站位最高, 达346.7×103ind·km-2, 绝大部分由龙头鱼构成。调查海域南部区域的尾数密度明显低于北部区域, 内侧水域明显低于外侧水域(图3)。
3.2.2 优势种及其数量时空分布
根据对本次舟山近岸海域拖网渔获物中鱼类的测定, 春季优势种为六丝钝尾虾虎鱼(表1), 其平均重量密度和尾数密度分别为4.37 kg·km-2和16.66× 103ind·km-2, 分别占春季总密度的42.18%和63.37%, 均以东部外侧海域出现数量较多, 而以杭州湾附近海域最低(图4、图5); 秋季优势种为龙头鱼(表1), 该物种是秋季调查海域优势度最高的种类, 其平均重量密度和尾数密度分别为42.69 kg·km-2和26.82× 103ind·km-2, 分别占总密度的43.56%和84.82%, 均以东部及东北部海域出现数量较多, 而调查海域南部较低(图4、图5)。
3.2.3 鱼类种数的空间分布和季节变化
舟山近岸海域春、秋季平均每个站位出现鱼类的种数分别为8种和7种, 其空间分布情况如图6所示, 图中等值线中的数值代表种类数。
由图6可知, 春季, 各站位种类数分布在3—17种之间, 由西向东呈逐渐增多的趋势, 以13号站位物种数最高, 物种数在10种以上的站位只有29个, 岛屿附近的物种数高于其他区域, 物种数最低的29和49号站位都处于较开阔的海域; 秋季, 各站位种类数分布在2—24种之间, 自西向东北有逐渐增多的趋势, 以4号站位的物种数最高, 物种数在10种以上的站位只有21个, 少于春季, 物种数较多的地区出现在嵊泗列岛附近。

图2 舟山近岸海域春、秋季鱼类重量密度分布

图3 舟山近岸海域春、秋季鱼类尾数密度分布
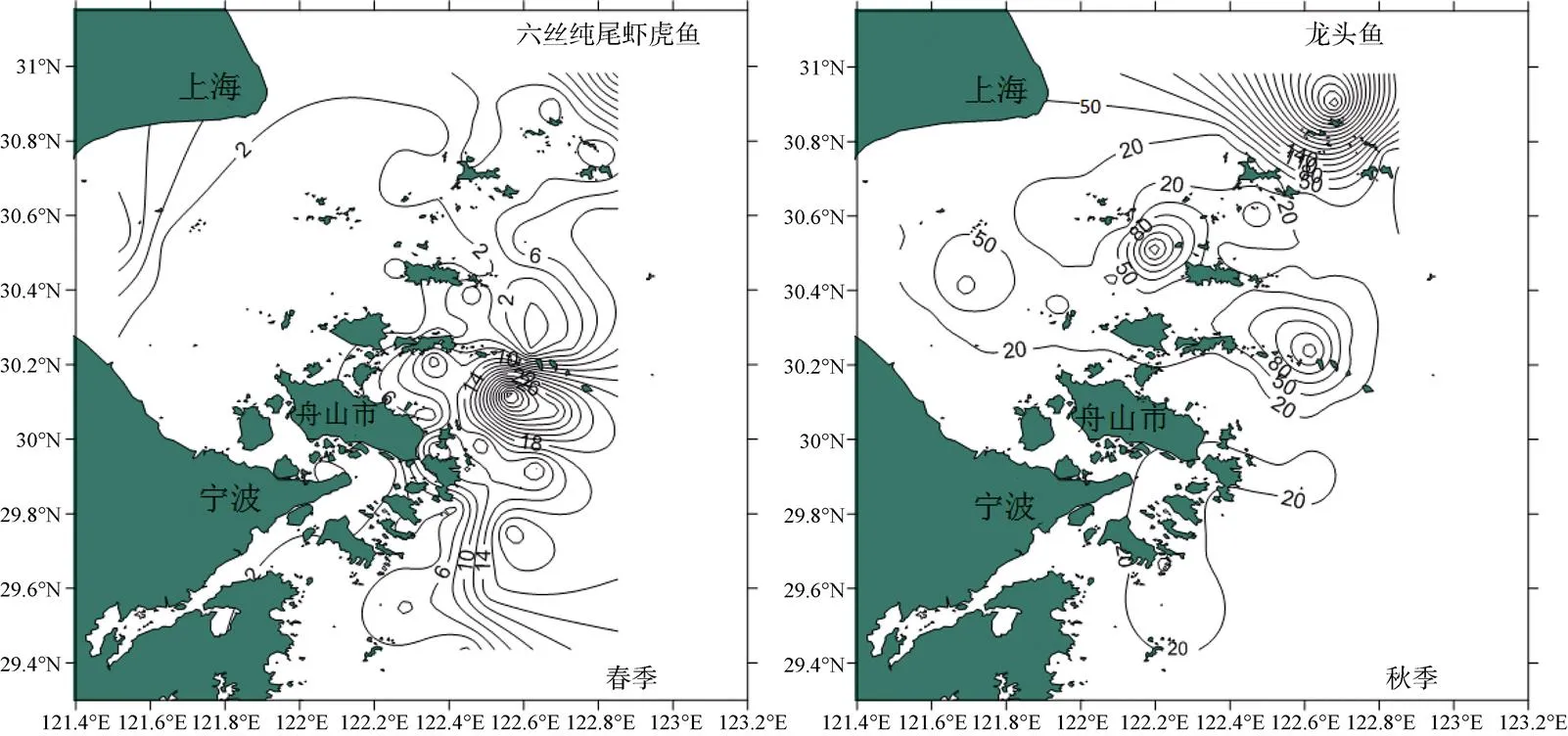
图4 舟山近岸海域鱼类优势种重量密度分布
3.2.4 不同水深条件下鱼类密度以及种类数的季节变化
不同水深带秋季的重量密度都高于春季, 而尾数密度除小于15 m水深带低于春季外, 其余两个水深带均高于春季。春季, 在15—45 m水深带鱼类的重量密度和尾数密度均高于其余两个水深带,而大于45 m水深带鱼类的密度最低。秋季, 在大于45 m水深带鱼类的重量密度和尾数密度都最高, 并且随水深增加有逐渐增加的趋势, 在15—45 m水深带最低(表2)。
不同水深之间鱼类种类数变化较大, 总种数以15—45 m的范围内最多。从不同季节来看, 春、秋季以小于15 m水域的鱼的种类与秋季相差最大, 水深越深, 秋季与春季种类数差距越大(表3)。

图5 舟山近岸海域鱼类优势种尾数密度分布

图6 舟山近岸海域春、秋季鱼类种数分布
表2 舟山近岸海域不同水深鱼类资源密度分布

Tab.2 The distribution of resource density of fish in different depth in Zhoushan coastal waters
表3 舟山近岸海域不同水深鱼类种类数的季节变化

Tab.3 The seasonal variation of fish species number in different depth in Zhoushan coastal waters
3.3 ABC曲线特征
调查海域春、秋季丰度和生物量曲线(ABC曲线)如图7所示。由图7可知, 春季和秋季舟山近岸海域的丰度曲线始终比生物量曲线要高。随着时间的推移, 两条曲线慢慢接近, 之后几乎重合在一起, W统计值均为负值。秋季生物量曲线的起点高于春季生物量曲线, 表明秋季的优势度高于春季。
4 讨论
4.1 鱼类种类组成
根据本次调查结果, 共鉴定出鱼类81种, 其中春季58种, 秋季53种, 秋季种类略少于春季其原因可能是因为秋季台湾暖流减弱[16], 导致该海域水温有所降低, 种类数会随着水温的降低有所减少, 而且一些鱼类会向外海深水区进行洄游。春季和秋季尾数密度都较高但重量密度却不是很高, 很大原因是因为春季幼鱼占绝大部分, 尾数多重量低, 而秋季多以小型鱼为主, 大型鱼类少有捕到。
将本次调查结果分别与60年代初成庆泰等[17]和2006、2007年俞存根等[18]在该海域调查结果相比可发现, 60年代初和2006、2007年两次调查秋季的种类数(分别为89种、79种)都高于春季(分别为39种、75种), 并且具有较多经济价值高且群体数量大的种类。而本次结果与之相反(为春季58种大于秋季53种), 一方面是因为春季有多种鱼类洄游到该海域进行生殖产卵[18], 另一方面可能是由于本次调查春季的降雨量较大, 降雨量的增加会使河口的生态类群往适应较低盐度的类群改变[19], 从而使河口区的种类增加, 其余原因如具体调查时间、调查站位设置的影响还有待进一步研究。但鱼类种类数总体较过去两次调查相比明显减少, 种类组成也发生了较大的变化, 这不仅与使用的调查网具有关, 本次调查使用的是底拖网而过去两次调查均使用的是桁杆拖虾网, 不同网具捕获的对象不同, 更多的可能是过度捕捞引起渔业资源优势种群衰退和群落结构发生演替有关[18]。并且本次调查海域主要是在舟山近岸海域, 不如过去的调查范围广, 近岸海域不仅是陆地与海洋进行物质和能量交换的重要区域, 也是海洋工程建设的场所, 容易受到人类活动的影响, 环境也更恶劣, 因此生态系统更显复杂和脆弱, 导致鱼类种类减少。
4.2 主要优势种分析
春季优势种为六丝钝尾虾虎鱼, 秋季优势种为龙头鱼, 不同季节优势种存在差异。六丝钝尾虾虎鱼的经济价值不高, 但却是本次调查海域春季的优势种, 可见以前的大型高值鱼类种群已逐渐被小型低值的种群所代替, 这种趋势与其他学者研究结果较一致[20-21]。龙头鱼的优势度在秋季最高, 很大可能是因为龙头鱼是杂食性、广温广盐性鱼类, 生存能力较强, 并常至河口水域觅食[22-23]常栖息于浅海底泥的环境中, 本次调查范围所涉站点大多在岛屿附近, 浅海环境居多, 因此渔获数量较高。根据李建生等[24-25]和程家骅等[26]对舟山渔场优势种的研究可发现, 近50年来, 舟山渔场的优势种发生了很大的变化, 鱼类优势种逐渐由以底层和近底层为主演变为以小型中上层鱼类为主, 这种变化趋势同其他学者研究结果相一致[26-27], 这种情况出现的原因最为突出的是过度捕捞, 其次是环境污染等导致生境发生变化, 从而致使鱼类等生物种类逐渐减少。
图7 舟山近岸海域春、秋季鱼类的ABC曲线以及W统计值
Fig.7 The ABC curves and W value of fish in spring and autumn in Zhoushan coastal waters
4.3 数量时空分布特征
从鱼类数量的季节和时空分布变化(图2、图3)可以看出, 秋季鱼类的重量密度明显高于春季, 这可能是因为春季是鱼类主要的产卵季节[11], 所捕渔获中有大部分是鱼类幼体; 而秋季是鱼类的索饵肥育季节[28], 大多数鱼类长成成鱼, 鱼体长、体重等都有所增加, 规格也较春季大。春季和秋季均以东部外侧及东北部海域的渔获数量较高, 其原因是该区域所处的位置受到来自台湾暖流的高温高盐水和江浙沿岸水的共同影响[16], 使该区域水温和营养盐较其他区域高, 并且又在岛屿附近, 有上升流等流态的海流, 营养物质较丰富, 所以鱼类大多在此栖息。
从鱼类物种数的平面分布(图6)来看, 春季调查海域东部且在岛屿附近海域物种数高于其他海域, 其原因可能是因为春季大多数鱼类进行生殖洄游, 由深海或外海向浅海或近岸进行洄游[29], 并且近岸有沿岸径流等含营养物质较高的海流, 给某些鱼类成体和幼体的生长提供了饵料基础, 因而吸引大多数鱼类在此聚集产卵。秋季调查海域东北部靠近嵊泗列岛附近海域的种类数高于其他海域, 这可能是因为这里岛屿较多, 并且旅游业也较发达, 比较注重对该海区水域环境的保护, 水质较近岸好, 适合鱼类栖息生存。
4.4 不同水深对鱼类资源密度及种类数的影响
有研究表明, 影响游泳动物生长、发育、分布的因素主要有温度、盐度和水深等非生物环境因子, 而水深又是影响其群落结构变化的最主要因素[30-32], 不同深度的区域具有不同的环境[33]。根据本次调查结果可知, 鱼类资源密度和种类分布随水深变化明显, 春季鱼类的重量密度和尾数密度均在15—45 m的水深范围最高, 可见大部分鱼类主要栖息在这个水层, 而此次调查范围在舟山群岛近岸附近, 岛屿众多, 大部分站位都设在岛屿附近, 其水深较开阔区域水深浅, 海域水深多数在15—45 m的范围内。秋季鱼类的重量密度和尾数密度在大于45 m的范围最高, 其原因可能是因为在该水深范围内的站位只有5个, 因此平均密度较高。春季鱼类种类数以小于15 m的水深范围较多, 大于45 m水深的种类数最少, 秋季则是水深越深种类越多, 其原因是春季鱼类多数在浅海区进行产卵, 而秋季水深较深的地方温度高于浅水区, 鱼类要进行越冬洄游等活动所引起的。
4.5 ABC曲线分析
对ABC曲线进行分析可得, 春季舟山近岸海域鱼类个体较小, 不存在生物量占据明显优势的一些大个体。秋季的鱼类ABC曲线与春季基本类似, 说明秋季舟山海域鱼类也不存在生物量占据明显优势的一些大个体。一般认为正常稳定的群落, 生物量曲线始终位于丰度曲线上[34],但本次春、秋季鱼类的丰度优势度曲线却始终位于生物量优势度曲线上方, 表明舟山近岸海域的鱼类群落处于严重干扰的状态, 其原因可能是该海域受人为因素的影响较大, 使该海域稳定性降低[35]。对比春、秋季鱼类群落的ABC曲线可以发现, 虽然两个季节的曲线走向基本一致, 但春季的丰度曲线比生物量曲线要高得多, 说明春季鱼类个体更小, 相较丰度, 生物量更加不占优势; 春季W值比秋季更小也充分说明这一事实。
[1] 赵淑江, 夏灵敏, 李汝伟, 等. 舟山渔场的过去、现在与未来[J]. 海洋开发与管理, 2015, (02): 44–48.
[2] 王垚, 卢占晖, 张亚洲, 等. 春季舟山渔场鱼类种类组成研究[J]. 浙江海洋学院学报: 自然科学版, 2012, (02): 129–132.
[3] 王强华. 保护、整治舟山渔场的生态环境[J]. 农业经济问题, 1983, 12: 43–44.
[4] 倪海儿, 陆杰华, 舟山渔场主要渔业资源利用现状[J]. 应用生态学报, 2003, 04: 569–572.
[5] 倪海儿, 陆杰华. 舟山渔场渔业资源动态解析[J]. 水产学报, 2002, 05: 428–432.
[6] SHEN Jinao, CHENG Yanhong. On the deep sea demersal fish communities of the East China Sea[J]. Chinese Journal of Oceanology and Limnology, 1989, 7(02): 157–168.
[7] KISHIDA S, KITAJIMA T. On the species area relation and diversity of demersal fishes in the East China Sea[J]. Bull Seikai Reg Fish Res Lab, 1980, 55: 53–63.
[8] OTAKI H. Demersal fish resources in the East China and Yellow seas[J]. Marine Behaviour and Physiology, 1993, 22(4): 195–269.
[9] 朱元鼎. 黄海和东海海洋状况和经济生物区系的综合报告[C]. 太平洋西部渔业委员会第三次全体会议论文集. 北京:科学出版社, 1960.
[10] 曹东阳, 俞存根, 周青松, 等. 舟山渔场普陀山附近海域鱼类种类组成和数量分布[J]. 福建水产, 2015, (05): 353– 362.
[11] 陈华, 徐兆礼. 舟山渔场衢山岛海域春夏季鱼类数量变化[J]. 海洋渔业, 2009, (02): 179–185.
[12] 钟毅飞, 俞存根, 周青松, 等. 舟山渔场宁波-舟山港海域春秋季鱼类种类组成和数量分布[J]. 浙江海洋学院学报: 自然科学版, 2016, (01): 1–7.
[13] 詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995.
[14] 孙儒泳, 李庆芬, 牛翠娟, 等. 基础生态学[M]. 北京: 高等教育出版社, 2002.
[15] CLARKE K R, WARWICK R M. Change in marine communities: an approach to statistical analysis and interpretation: 2nd ed[M]. Plymouth: PRIMPER-E, 2001.
[16] 徐兆礼. 瓯江口海域夏秋季鱼类多样性[J]. 生态学报, 2008, 28(12): 5948–5956.
[17] 成庆泰, 郑葆珊. 中国鱼类系统检索[M]. 北京: 科学出版社, 1987.
[18] 俞存根, 陈全震, 陈小庆, 等. 舟山渔场及邻近海域鱼类种类组成和数量分布[J]. 海洋与湖沼, 2010, (03): 410– 417.
[19] VICTOR S K, ROBERT R T, JOAN A K, et al. Coastal and Marine Ecosystems & Global Climate Change—Potential Effects onU.S. Resources[M/OL]. [2002—08]. http: //www. pew climate org /doc Uploads/marine_ecosystems. Pdf.
[20] 徐宾铎, 金显仕, 梁振林. 秋季黄海底层鱼类群落结构的变化[J]. 中国水产科学, 2003, (02): 148–154.
[21] 卢占晖, 苗振清, 林楠. 浙江中部近海及其邻近海域春季鱼类群落结构及其多样性[J]. 浙江海洋学院学报: 自然科学版, 2009, (01): 51–56.
[22] 林龙山. 东海区龙头鱼数量分布及其环境特征[J]. 上海海洋大学学报, 2009, (01): 66–71.
[23] 杨星星, 洪小括, 叶海滨. 浙南沿岸龙头鱼数量分布调查[J]. 上海海洋大学学报, 2012, (03): 410–414.
[24] 李建生, 李圣法, 程家骅. 长江口渔场鱼类组成和多样性[J]. 海洋渔业, 2006, (01): 37–41.
[25] 李建生, 李圣法, 丁峰元, 等. 长江口近海鱼类多样性的年际变化[J]. 中国水产科学, 2007, 04: 637–643.
[26] 程家骅, 丁峰元, 李圣法, 等. 夏季东海北部近海鱼类群落结构变化[J]. 自然资源学报, 2006, 05: 775–781.
[27] 徐汉祥.跨世纪东海渔业资源利用和管理若干问题的探讨[J]. 浙江海洋学院学报: 自然科学版, 2000, 03: 197– 203.
[28] 徐兆礼. 夏秋季瓯江口海域鱼类数量的时空分布[J]. 动物学报, 2008, (06): 981–987.
[29] 夏陆军, 周青松, 俞存根, 等. 乐清湾口海域春秋季鱼类群落多样性研究[J]. 渔业现代化, 2016, (02): 68–75.
[30] ODUM E P, BARRETT C W. Fundamentals of Ecology[M]. Bejing: People’s Education Press, 1981.
[31] 邱永松. 南海北部大陆架鱼类群落的区域性变化[J]. 水产学报, 1988, (04): 303–313.
[32] COLVOCORESSES J A, MUSICK J A. Species associations and community composition of middle Atlantic Bight continental shelf demersal fishes[J]. Fish Bull, 1984, 82(2): 295–313.
[33] 王雪辉, 杜飞雁, 邱永松, 等. 大亚湾鱼类群落格局分析[J]. 生态科学, 2015,(06): 64–70.
[34] 韩婵, 高春霞, 田思泉, 等. 淀山湖鱼类群落结构多样性的年际变化[J]. 上海海洋大学学报, 2014, (03): 403–410.
[35] WARWICK R M. A new method for detecting pollution effects on marine macrobenthic communities[J]. Marine Biology, 1986, 92(4): 557–562.
Spatial and temporal variation in fish species composition and abundance in Zhoushan coastal waters during spring and autumn
DENG Xiaoyan1, YU Cungen1,*, SHUI Yuyue2, WENG Liangcai2, DING Jianwei2, ZHANG Ping1, XU Yongjiu1, ZHENG Ji1, XIE Xu1, MIAO Lu1
1. School of Fisheries Zhejiang Ocean University, Zhoushan 316022, China 2. Zhoushan Bureau of Oceanology and Fishery, Zhoushan 316000, China
Based on the data of fishery resource surveys in November, 2015(autumn) and May, 2016(spring) in Zhoushan coastal waters, we studied the species composition, quantitative distribution, dominant species variation and the influential factors. The results were as follows. According to the survey of the samples, a total of 81 fish species were identified, which belonged to 63 genera in 39 families of 12 orders. The biomass of fish in spring and autumn was 10.36 kg·km-2and 98.01 kg·km-2, respectively. The abundance of fish in spring and autumn was26.29×103ind·km-2and 33.30×103ind·km-2, respectively. The spatial distribution of fish resources density was generally similar in spring and autumn, and the density was higher in the east of the outside waters than the west of the east longitude 122.3 waters. Different types of dominant species were in different seasons:in spring andin autumn, and both of them were small and low-value fishes. The number of fish species variation was more stable in spring and autumn, but the seasonal variation of species number was larger. Most of species distributed near shallow sea of coast in spring,and most of species distributed in northeast of survey area in autumn. The depth of water had an obvious impact on the density of fish resources and distribution of species number. ABC curve showed that the community of fish was in a state of serious disturbance in the Zhoushan coastal waters. Compared with the previous findings, the species composition and community structure of fish in the Zhoushan fishing ground changed significantly.
fish; species composition; quantitative distribution; resource density; ABC curve; Zhoushan coastal waters
Q958.8; S932.4
A
1008-8873(2018)01-010-10
10.14108/j.cnki.1008-8873.2018.01.002
2017-01-13;
2017-03-20
国家自然科学基金项目(31270527); 舟山市海洋与渔业局委托项目2015.10—2017.03.
邓小艳(1991—), 女, 重庆北碚人, 硕士研究生, 主要从事渔业资源研究。E-mail: dxy672712@163.com
俞存根, 男, 教授, 主要从事渔业资源研究, E-mail: cgyu@zjou.edu.cn
邓小艳, 俞存根, 水玉跃,等. 舟山近岸海域春秋季鱼类种类组成及其数量时空分布[J]. 生态科学, 2018, 37(1): 10-19.
DENG Xiaoyan, YU Cungen, SHUI Yuyue, et al. Spatial and temporal variation in fish species composition and abundance in Zhoushan coastal waters during spring and autumn[J]. Ecological Science, 2018, 37(1): 10-19.
