Mn2+、Pb2+对野生垂穗披碱草种子萌发与幼苗生长的影响
2018-03-26徐雅梅王传旗武俊喜张文静王小川赤列催珍徐德飞包赛很那苗彦军
徐雅梅,王传旗,武俊喜,张文静,王小川,赤列催珍,徐德飞,包赛很那,苗彦军*
(1.西藏农牧学院,西藏 林芝 860000;2.中国科学院地理科学与资源研究所,北京 100101)
重金属污染是当今面积最广、危害最大的环境问题之一[1-2]。 虽然有些金属如Mn、Cu、Zn、Mo等为植物不可缺少的矿物质营养元素,但是当植物体内重金属过量或超过某一临界值时,就会对植物产生胁迫作用,导致其体内代谢过程紊乱,生长发育受到抑制,甚至导致植物枯萎、死亡[3]。以往,由于藏中部地区金属矿的开采,造成了矿区土壤与水体重金属严重污染,使得矿区及其周边地区的生存环境适宜性降低,生物多样性下降,生态系统脆弱性增加[4]。矿区退化植被恢复成为生态恢复研究的热点和焦点[5]。植物修复技术是在不破坏土壤生态环境、保持土壤结构和微生物活性的情况下,通过植物根系及其共存微生物体系直接将大量的重金属元素吸收,进而收获植物地上部分并将它们无害化处理,从而达到治理修复被污染的土壤的目的[6-9]。
垂穗披碱草(Elymusnutans)为禾本科披碱草属牧草,又名钩头草、弯穗草[10]。原为野生种,在我国西藏及西北、华北等地区均有分布,在青藏高原海拔2500~4000 m的高寒湿润地区为建群种,是有价值的饲用植物[11]。除具有饲用价值外,生态适应性广,具有防风固沙、保持水土、抗旱、抗寒、耐盐碱等优点,对生态保护和生态恢复有重要意义[12]。Mn2+、Pb2+胁迫对西藏地区野生垂穗披碱草种子萌发及幼苗生长的研究尚未见详细报道。因此,本研究选择了西藏海拔较高的巴青县野生垂穗披碱草为试验材料,探讨了重金属Mn2+、Pb2+离子对其种子萌发和幼苗生长的生理影响,以期为西藏矿区退化植被恢复和土壤重金属污染的治理提供基础。
1 材料与方法
1.1 试验材料
供试材料于2003年9月采自西藏那曲区巴青县(31°53′57″ N,93°10′44″ E,海拔4675 m)。之后在西藏农牧学院农场(29°39′46″ N,94°21′31″ E,海拔2990 m)经过13年人工选育栽培。试验于2017年4-7月,在西藏高原作物栽培学与耕作学实验室进行。试验前在室温条件下做预试验,测得供试材料种子千粒重为(4.8±0.05) g,平均发芽率为91.5%。
1.2 试验方法
选取籽粒饱满、大小均一的种子,用0.5% KMnO4溶液消毒15 min,然后用蒸馏水冲洗干净,自然晾干备用[13]。供应金属离子试剂MnCl2·4H2O和Pb(NO3)2,均为分析纯。根据中华人民共和国国家土壤环境质量标准(GB15618-2008)中的三级土壤环境标准量设定重金属离子浓度[14],试验设定浓度依次为0,200,400,600,800,1000 mg·L-1。以蒸馏水为对照。
种子发芽阶段:试验采用纸上发芽法,每个塑料发芽盒内铺双层滤纸,放入100粒种子,将配置好的金属溶液置于发芽盒内,至滤纸饱和为止。每个处理均为5次重复。系列发芽试验均在RXZ智能人工气候箱内进行。温度设置为20 ℃/30 ℃,每天12 h/12 h(低温/高温),12 h/12 h(光照/黑暗)依次交替,光照强度400 μmol·m-2·s-1。试验培养周期为15 d,每隔2 d用万分之一天平称重一次,并使用蒸馏水补充因挥发而失去的水分,期间保持发芽盒湿润且无明水。以芽长超过种子本身长度的1/2作为种子发芽的标志,每天统计发芽数并及时清理发霉腐烂的种子。第7天统计发芽势,第15天时统计种子发芽率、发芽指数和活力指数并测量幼苗芽长、根长。每个指标3次重复,取平均值。
苗期阶段:待种子培养结束之后,对各金属浓度下的垂穗披碱草幼苗叶片进行电导率、叶绿素含量、丙二醛含量的测定。每个指标3次重复,取平均值。
1.3 测定指标及方法
1.3.1种子萌发参数的计算公式
种子发芽率(germination percentage) GP=(n/N)×100%
式中:n为最终正常发芽种子数,N为供试种子数。
发芽势(germination vigor) GV=(7 d内种子正常发芽数/供试种子数)×100%[15]
发芽指数(germination index) GI=∑(Gt/Dt)
式中:Gt指t日发芽数,Dt指相应的天数[16]。
活力指数(vigor index) VI=GI×Sx
式中:Sx为根平均长度[17]。
胚根和胚芽长的测定:从每个处理浓度的塑料发芽盒内随机取20个正常生长的幼苗用直尺进行胚根和胚芽长的测量,不够20株的全部都测,取平均值。
1.3.2幼苗部分生理指标的测定 叶绿素含量测定采用丙酮浸提法[18],细胞质膜透性测定采用电导仪法[18],丙二醛含量测定采用硫代巴比妥酸法[18]。
1.4 数据处理与分析
使用DPS 6.0统计分析软件进行方差分析,样本间的差异显著性用Tukey检验,图表用WPS软件制作,相关指标数据以平均值±标准差表示。
2 结果与分析
2.1 Mn2+、Pb2+对垂穗披碱草种子萌发的影响
由表1和表2可知,Mn2+、Pb2+对巴青野生垂穗披碱草种子发芽率具有极显著影响(P<0.01),且随着Mn2+、Pb2+溶液浓度的升高,抑制作用越明显。200 mg·L-1Mn2+、Pb2+溶液浓度处理下,垂穗披碱草种子发芽率略低于CK,高于400 mg·L-1处理时则出现较大幅度的下降;当Mn2+、Pb2+溶液浓度为1000 mg·L-1时,发芽率分别较CK下降了70%和53%。
垂穗披碱草种子发芽势随Mn2+溶液浓度的升高呈下降趋势。垂穗披碱草在Mn2+溶液浓度为0和200 mg·L-1时,种子发芽势分别为69%和66%,二者无显著性差异;600、800 mg·L-1Mn2+溶液浓度处理下,与对照相比,发芽势显著下降(P<0.05);当Mn2+溶液浓度达到1000 mg·L-1时,种子发芽势仅16%,与CK相比具有极显著差异(P<0.01)。200、400 mg·L-1Pb2+溶液浓度下垂穗披碱草发芽势和CK无显著差异;600、800 mg·L-1Pb2+溶液浓度下垂穗披碱草种子发芽势显著低于CK(P<0.05);当Pb2+溶液浓度为1000 mg·L-1时,种子发芽势达到了最低水平(表1和表2)。
Mn2+、Pb2+离子胁迫对垂穗披碱草种子发芽指数的影响与发芽率和发芽势基本相似。巴青野生垂穗披碱草种子发芽指数随Mn2+、Pb2+溶液浓度的升高而降低。在Mn2+、Pb2+溶液浓度为200 mg·L-1时,其发芽指数分别为55和52,较对照略有下降,但无显著性差异;在高浓度1000 mg·L-1Mn2+、Pb2+处理下,其发芽指数分别为34和30,较CK具有极显著差异(P<0.01)(表1和表2)。
西藏巴青野生垂穗披碱草种子活力指数随Mn2+、Pb2+溶液浓度的升高呈不断下降的态势。垂穗披碱草在各Mn2+、Pb2+溶液浓度处理下活力指数均显著(P<0.05)或极显著(P<0.01)低于CK,且同等条件下Mn2+比Pb2+的抑制作用显著,即Mn2+离子胁迫对垂穗披碱草种子活力指数的毒害更大(表1和表2)。
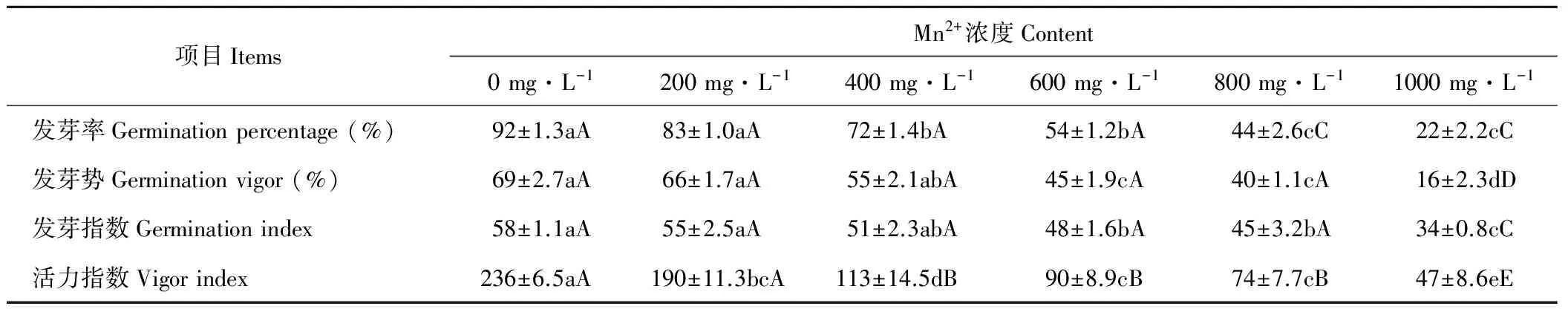
表1 Mn2+对垂穗披碱草种子萌发的影响Table 1 Effects of Mn2+ on the seed germination of E. nutans
注:不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01),下同。
Note: The different small and capital letters represent significant differences at 0.05 and 0.01 level, respectively. The same below.

表2 Pb2+对垂穗披碱草种子萌发的影响Table 2 Effects of Pb2+ on the seed germination of E. nutans
由图1和图2可知,Mn2+、Pb2+处理对巴青野生垂穗披碱草胚根和胚芽的生长影响较大。整体上,垂穗披碱草胚根和胚芽长随重金属离子处理浓度的升高表现为下降的趋势。正常处理条件下,垂穗披碱草胚芽、胚根长为13.80 cm和5.69 cm;当Mn2+、Pb2+溶液浓度为1000 mg·L-1时,其胚芽、胚根长度均达到最低水平,与CK相比,胚芽具有极显著差异(P<0.01),而胚根具有显著差异(P<0.05);200,400,600,800 mg·L-1Mn2+溶液浓度下,与对照相比,胚芽长显著下降(P<0.05)。在200 mg·L-1Mn2+、Pb2+溶液浓度处理下,垂穗披碱草胚根比CK略长。
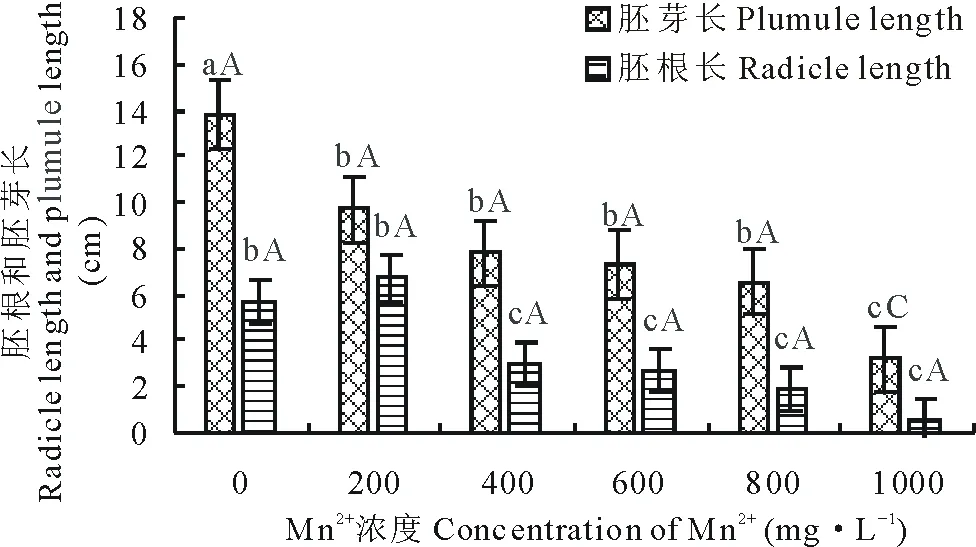
图1 Mn2+对垂穗披碱草胚根长和胚芽长的影响Fig.1 Effects of Mn2+ on the radicle length and plumule length of E. nutans

图2 Pb2+对垂穗披碱草胚根长和胚芽长的影响Fig.2 Effects of Pb2+on the radicle length and plumule length of E. nutans
不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)。The different small and capital letters represent significant differences at 0.05 and 0.01 level, respectively.
2.2 Mn2+、Pb2+对垂穗披碱草幼苗部分生理指标的影响
2.2.1Mn2+、Pb2+对垂穗披碱草幼苗叶绿素含量的影响 由图3和图4可知,巴青野生垂穗披碱草幼苗叶片中叶绿素a和叶绿素b含量随着Mn2+、Pb2+离子胁迫程度的增加呈下降趋势。其中,Mn2+离子浓度与叶绿素a和b的相关关系分别为y=-2.16x+13.87 (R2=0.95),y=-1.68x+11.14 (R2=0.97);Pb2+离子浓度与叶绿素a和b的相关关系分别为y=-2.13x+14.23 (R2=0.95),y=-1.64x+11.50 (R2=0.99)。可见,Mn2+、Pb2+离子浓度与垂穗披碱草叶绿素a和b的含量呈负相关。
2.2.2Mn2+、Pb2+对垂穗披碱草幼苗MDA含量的影响 MDA的含量是植物在逆境中受到伤害程度的标志之一。Mn2+、Pb2+对垂穗披碱草幼苗叶片MDA含量具有极显著影响(P<0.01)(图5)。幼苗叶片MDA含量随着Mn2+、Pb2+溶液浓度的升高而增加,几乎呈直线水平上升。其中,Mn2+离子浓度与垂穗披碱草幼苗叶片MDA含量的相关关系为y=6.28x-4.07 (R2=0.99);Pb2+离子浓度与垂穗披碱草幼苗叶片MDA含量的相关关系为y=5.31x-4.47 (R2=0.98)。
2.2.3Mn2+、Pb2+对垂穗披碱草幼苗叶片质膜透性的影响 随着Mn2+、Pb2+溶液浓度的升高,垂穗披碱草幼苗叶片电导率明显升高(图6)。1000 mg·L-1Mn2+、Pb2+溶液浓度处理下,幼苗叶片电导率达到最大值,与对照相比,分别增加了38.90%和34.66%。Mn2+离子胁迫浓度与垂穗披碱草幼苗叶片电导率的相关关系为y=8.07x-2.44 (R2=0.99);Pb2+离子胁迫浓度与垂穗披碱草幼苗叶片电导率的相关关系为y=7.36x-2.99(R2=0.98)。可见,Mn2+、Pb2+胁迫浓度与垂穗披碱草幼苗叶片电导率呈正相关。

图3 Mn2+、Pb2+对垂穗披碱草叶绿素a含量的影响Fig.3 Effects of Mn2+, Pb2+ on the chlorophyll a content of E. nutans

图4 Mn2+、Pb2+对垂穗披碱草叶绿素b含量的影响Fig.4 Effects of Mn2+, Pb2+ on the chlorophyll b content of E. nutans

图5 Mn2+、Pb2+对垂穗披碱草MDA含量的影响Fig.5 Effects of Mn2+, Pb2+ on the malondialdehyde content of E. nutans

图6 Mn2+、Pb2+对垂穗披碱草叶片电导率的影响Fig.6 Effects of Mn2+, Pb2+ on the electrical conductance of E. nutans
3 讨论
本研究结果表明,Mn2+、Pb2+对巴青野生垂穗披碱草种子萌发及幼苗生长具有极显著影响(P<0.01)。Mn2+、Pb2+在抑制垂穗披碱草种子萌发和幼苗生长过程中表现出了一定的相似性。随着 Mn2+、Pb2+溶液浓度的升高,种子发芽率、发芽势、发芽指数和活力指数出现了不同程度的下降趋势,这与多人研究结果相类似[19-20]。可能是Mn2+、Pb2+离子抑制了种子蛋白质的分解,使酶活性降低,从而影响了种子萌发。
根是植物吸收和积累重金属的主要器官,也是最直接的受害部位。试验显示,随着Mn2+、Pb2+溶液浓度的升高,整体上西藏巴青县野生垂穗披碱草种子胚芽和胚根长表现为下降的趋势,这和其他植物对金属胁迫的反应一致[21]。原因可能是高浓度培养液使大量Mn2+、Pb2+富集在植物根部,使得根尖细胞无法正常分裂或分裂速度减慢,抑制了根的伸长,最终影响到整个植株的生长[22]。然而,试验发现,200 mg·L-1Mn2+、Pb2+溶液浓度处理下垂穗披碱草胚根比CK略长,但差异不显著。说明在轻度金属胁迫条件下垂穗披碱草更多的生物量用于根部生长,即将有限的养分资源优先分配到根部以求得生存。
叶绿素含量的多少与牧草的光合作用及其强度密切相关[23]。本试验中,垂穗披碱草幼苗叶片叶绿素a和b均随 Mn2+、Pb2+溶液浓度的升高而降低,可能是金属胁迫使叶绿体内的酶活性比例失调,叶绿素分解加快而导致叶绿素含量下降。这与岑画梦等[24]在Cd2+对狗牙根(Cynodondactylon)、假俭草(Eremochloaophiuroides)种子萌发及幼苗生长的影响研究结果相似。
MDA是膜脂质过氧化作用的主要产物,其含量成为鉴别植物细胞膜破坏程度的指标之一。本研究显示,随着 Mn2+、Pb2+溶液浓度的升高,垂穗披碱草幼苗叶片MDA呈上升趋势,可能是Mn2+、Pb2+离子引起氧化胁迫导致植物细胞膜脂过氧化,且随着金属离子胁迫浓度的升高幼苗叶片MDA含量越高[25]。与叶片MDA含量变化趋势相似,幼苗叶片电导率随着Mn2+、Pb2+溶液浓度的升高呈上升变化趋势。可能是高浓度的金属胁迫破坏了幼苗叶片细胞膜的完整性,使细胞内的一些可溶性物质外渗,导致膜渗透性增大[26]。
4 结论
Mn2+、Pb2+对西藏巴青县野生垂穗披碱草种子发芽率、发芽势、发芽指数和活力指数具有抑制作用,并对垂穗披碱草胚根和胚芽的生长产生抑制效应。西藏巴青县野生垂穗披碱草对Mn2+、Pb2+具有一定的耐受性,即使在800 mg·L-1Pb2+和600 mg·L-1Mn2+溶液浓度处理下,其发芽率仍高达50%以上。因此,本研究建议西藏巴青县野生垂穗披碱草可作为治理西藏Mn2+、Pb2+土壤污染与植被修复的重要备选草种。
本试验仅研究了Mn2+、Pb2+对西藏巴青县野生垂穗披碱草种子萌发及幼苗生长的影响,在一定程度上揭示了垂穗披碱草对Mn2+、Pb2+金属胁迫的生理响应机理。但土壤重金属污染大部分是属于复合污染,因此关于巴青县野生垂穗披碱草对于其他金属的生理响应机制还需要进一步的探索与研究。
References:
[1] Jia W J. Effects of cadmium on seed germination, seeding growth and antioxidant system ofThellungielahalophila. Jinan: Shandong Normal University, 2012.
贾文娟. 镉胁迫对盐芥种子萌发、幼苗生长及抗氧化酶的影响. 济南: 山东师范大学, 2012.
[2] Yang M, Li X Y. Dynamic changes and cause analysis of heavy metals in street dust in Guiyang City China. Acta Scientiae Circumstantiae, 2014, 34(8): 2070-2076.
杨梅, 李晓燕. 贵阳市冬季地表灰尘重金属含量动态变化及原因探析. 环境科学学报, 2014, 34(8): 2070-2076.
[3] Duan D C, Yu M G, Shi J Y. Research advances in uptake, translocation, accumulation and detoxification of Pb in plants. Chinese Journal of Applied Ecology, 2014, 25(1): 287-296.
段德超, 于明革, 施积炎. 植物对铅的吸收、转运、累积和解毒机制研究进展. 应用生态学报, 2014, 25(1): 287-296.
[4] Zhang F P, Cao C G, Li P,etal. Effects of heavy metal pollution on microbial characteristics of mine soils in central Tibet. Journal of Agro-Environment Science, 2010, 29(4): 698-704.
张涪平, 曹凑贵, 李苹, 等. 藏中矿区重金属污染对土壤微生物学特性的影响. 农业环境科学, 2010, 29(4): 698-704.
[5] Jing J W, Zhao Y H, Zhang F P,etal. Evaluation of heavy metals pollution of surface soil in central Tibet mining area. Guizhou Agricultural Sciences, 2011, 39(7): 126-128.
敬久旺, 赵玉红, 张涪平, 等. 藏中矿区表层土壤重金属污染评价. 贵州农业科学, 2011, 39(7): 126-128.
[6] Fan T, Ye W L, Chen H Y,etal. Review on contamination and remediation technology of heavy metal in agricultural soil. Ecology and Environmental Sciences, 2013, 22(10): 1727-1736.
樊霆, 叶文玲, 陈海燕, 等. 农田土壤重金属污染状况及修复技术研究. 生态环境学报, 2013, 22(10): 1727-1736.
[7] Hu R, Su X, Yan M S,etal. Effects of Pb (Ⅱ) on seed germination of three different plants of Chenopodiaceae. Bulletin of Botanical Research, 2009, 29(3): 362-367.
虎瑞, 苏雪, 晏民生, 等. 重金属Pb(Ⅱ)对3种藜科植物种子萌发的影响. 植物研究, 2009, 29(3): 362-367.
[8] Hei L. Research progress on mechanism and prospect of phytoremediation of soils contaminated by heavy metals. Journal of Anhui Agricultural Sciences, 2014, 42(18): 5739-5740, 5777.
黑亮. 植物修复技术治理土壤重金属污染的机制研究进展及其应用前景. 安徽农业科学, 2014, 42(18): 5739-5740, 5777.
[9] Wang W, Zhang J T, Wang W,etal. Analysis of spatial differentiation of benefits of phytoremediation of heavy metal contaminated soil using coupled model. Acta Pedologica Sinica, 2014, 51(3): 547-554.
王维, 张金婷, 王伟, 等. 基于耦合模型的重金属污染土壤植物修复效益空间差异分析. 土壤学报, 2014, 51(3): 547-554.
[10] Lu G P, Nie B. Field evaluation ofElymusnutansunder alpine grassland conditions. Pratacultural Science, 2002, 19(9): 13-15.
陆光平, 聂斌. 垂穗披碱草利用价值评价. 草业科学, 2002, 19(9): 13-15.
[11] Chen M J, Jia S X. Feeding plants. Beijing: China Agricultural Press, 2002: 119-120.
陈默君, 贾慎修. 中国饲用植物. 北京: 中国农业出版社, 2002: 119-120.
[12] Wang C Q, Miao Y J, Wang J L,etal. Studies on drought resistance of wildElymusnutansin Tibet under drought stress at seedling stage. Chinese Journal of Grassland, 2017, 39(4): 116-120.
王传旗, 苗彦军, 王建林, 等. 西藏野生垂穗披碱草苗期抗旱性研究. 中国草地学报, 2017, 39(4): 116-120.
[13] Wang C Q, Liang S, Zhaxi Ciren,etal. Effects of temperature on Tibet three wildElymusforage seed germination. Journal of Tibet University (Natural Science Edition), 2016, (2): 64-67.
王传旗, 梁莎, 扎西次仁, 等. 温度对西藏3种野生披碱草属牧草种子萌发的影响. 西藏大学学报(自然科学版), 2016, (2): 64-67.
[14] Gao F F. The effects of Cu, Zn, Cd and Pb on the growth of three leguminous plants and the metal-absorption abilities of the plants. Shenyang: Northeastern University, 2008: 13-14.
高菲菲. Cu、Zn、Cd、Pb对三种豆科植物生长的影响及其吸附性能的研究. 沈阳: 东北大学, 2008: 13-14.
[15] Sun Y R, Shi Y, Chen G J,etal. Evaluation of the germination characteristics and drought resistance of green manure crops under PEG stress. Acta Prataculturae Sinica, 2015, 24(3): 89-98.
孙艳茹, 石屹, 陈国军, 等. PFG模拟干旱胁迫下8种绿肥作物萌发特性与抗旱性评价. 草业学报, 2015, 24(3): 89-98.
[16] Ji Y, Zhang X Q, Peng Y,etal. Seed germination response to osmotic stress and tolerance evaluation of Orchardgrass. Acta Agrestia Sinica, 2013, 21(4): 737-743.
季杨, 张新全, 彭燕, 等. 鸭茅种子萌发对渗透胁迫响应与耐旱性评价. 草地学报, 2013, 21(4): 737-743.
[17] Guo J M, Liu J, Dong K H. Effect of PEG-6000 stress on seed germination ofBothriochicaischaemum. Chinese Journal of Grassland, 2015, 37(2): 58-62.
郭晋梅, 刘娟, 董宽虎. PEG胁迫对白羊草种子萌发的影响. 中国草地学报, 2015, 37(2): 58-62.
[18] Gao J F. The experimental guide for plant physiology. Beijing: Higher Education Press, 2006.
高俊风. 植物生理学实验教程. 北京: 高等教育出版社, 2006.
[19] Yu X J, Zhang J W, Pan T T,etal. Effects of heavy metals: Copper, Cadmium and Lead on the seed germination and seedling growth of Leguminous forage. Acta Agrestia Sinica, 2015, 23(4): 794-803.
鱼小军, 张建文, 潘涛涛, 等. 铜、镐、铅对7种豆科牧草种子萌发和幼苗生长的影响. 草地学报, 2015, 23(4): 794-803.
[20] Chen W, Zhang M M, Song Y Y,etal. Inhibitory effect of heavy metal stress on the seed germination of four turfgrass types. Acta Agrestia Sinica, 2013, 21(3): 557-563.
陈伟, 张苗苗, 宋阳阳, 等. 重金属胁迫对4种草坪草种子萌发的影响. 草地学报, 2013, 21(3): 557-563.
[21] Zhao Y H, Laba Quji, Luo B,etal. Effects of heavy metals opper, cadmium, lead and zinc on seed germination and seedling growth of Leguminous species. Seed, 2017, 36(1): 22-27.
赵玉红, 拉巴曲吉, 罗布, 等. 铜、镉、铅、锌对4种豆科植物种子萌发的影响. 种子, 2017, 36(1): 22-27.
[22] Wang J, Guo Y, Wu R,etal. Effects of different planting years and organic manure fertilization on Zn accumulation in greenhouse soil. Journal of Agro-Environment Science, 2009, 28(1): 89-94.
王俊, 郭颖, 吴蕊, 等. 不同种植年限和施肥量对日光温室土壤锌累积的影响. 农业环境科学学报, 2009, 28(1): 89-94.
[23] Qi J, Xu Z, Wang H Q,etal.ElymussibiricusL. andElymusdahuricusat seedling stage and comprehensive evaluation of drought resistance. Acta Agrestia Sinica, 2009, 17(1): 36-42.
祁娟, 徐柱, 王海清, 等. 披碱草与老芒麦苗期抗旱性综合评价. 草地学报, 2009, 17(1): 36-42.
[24] Cen H M, Peng L L, Yang X,etal. Effects of Cd2+on the seed germination and seedling growth ofCynodondactylonandEremochloaophiuroides. Acta Prataculturae Sinica, 2015, 24(5): 100-107.
岑画梦, 彭玲莉, 杨雪, 等. Cd2+对狗牙根、假俭草种子萌发及幼苗生长的影响. 草业学报, 2015, 24(5): 100-107.
[25] Sclote D S, Khanna-Chopra R. Drought acclimation confers oxidative stress tolerance by inducing co-ordinated antioxidant defense at cellular and subcellular level in leaves of wheat seedlings. Physiologia Plantarum, 2006, 127: 494-506.
[26] Zhen Q, Yan M, Yang H F,etal. Coercion and damage of Cu pollution onArtemisialavandulaefoliagrowth. Chinese Journal of Applied Ecology, 2006, 17(8): 1505-1510.
甄泉, 严密, 杨红飞, 等. 铜污染对野艾蒿生长发育的胁迫及伤害. 应用生态学报, 2006, 17(8): 1505-1510.
