Gamma神经振荡和信息整合加工
2018-03-24钱浩悦黄逸慧高湘萍
钱浩悦 黄逸慧 高湘萍
(上海师范大学教育学院心理系,上海200234)
1 引言
日常生活中,外部世界的信息常常以片段的形式得到加工,比如散落的色点,闪现的图像与声音。大脑是如何把这些来自单一或多个感觉通道的碎片化信息进行整合,知觉为一个有意义的客体对象?这一问题近年来受到来自各种研究取向脑科学家的高度关注(Yau,DeAngelis,&Angelaki, 2015)。其中有些研究者提出,人类所表现出的信息整合功能与神经振荡有关,特别是Gamma频段神经振荡(Senkowski, Schneider, Foxe,& Engel, 2008)。然而,有多位神经学家重新分析了 Gamma神经振荡(Gamma振荡)与人类认知功能的关系后,对“Gamma振荡对应于特征整合加工”的观点提出了反驳(Herrmann, Früend, & Lenz,2010; Bosman, Lansink, & Pennartz, 2014; Merker,2013)。尽管如此,大量的研究一致表明,Gamma振荡活动在人类实现“信息整合”功能中扮演着非常重要的角色。本文首先将回顾有关单一及多通道(跨通道)信息整合加工与Gamma振荡关系的研究;其次,从生理角度,阐释 Gamma振荡的生理基础及特点与整合加工的联系;然后,从多层级视角,论述Gamma振荡在“信息整合加工”中具体的作用机理;最后,就Gamma振荡是否可以作为判定整合加工发生的指标以及Gamma振荡在复杂情景的整合加工中将如何发挥作用,这两个热点问题进行探讨和展望。
2 Gamma振荡与人类的信息整合加工
2.1 单通道的信息整合与Gamma振荡
日常生活中,人们常基于个体的经验和知识整合纷繁的视觉信息或听觉信息,形成对一个有意义对象的完整认识(Klemen & Chambers, 2012)。已有大量研究对这种单通道整合加工与Gamma振荡的关系进行了探索。
2.1.1视觉
关于神经振荡与视觉信息整合加工的关系探索始于 Keil, Müller, Ray, Grube和Elbert (1999)的研究。在那个实验中,他们记录了被试看一张旋转的“Mooney”面孔两可图(正立为笑脸,倒立为哭脸)时的大脑 EEG信号。EEG结果显示,旋转的“Mooney”面孔图比静止的正立面孔图激发了更大的Gamma振荡,并且在旋转到最易于识别位置时(正立)的Gamma振荡最大。这一发现表明,在面孔视觉识别的加工中,Gamma振荡起到了加强整体性识别的作用。后续,通过整体 EEG/ERP范式结合向量机分类(vector machineclassification)的方法,Castelhano,Rebola, Leitão, Rodriguez和Castelo-Branco (2013)确认了Gamma振荡确实反映了自上而下地对“Mooney”对象(面孔和吉他,如图 1a)的整体性识别加工, 而非简单的知觉加工。近期, Moratti, Méndez-Bértolo, Del-Pozo 和Strange (2014)使用动态因果模型的方法, 清晰地展现了 Gamma振荡是如何在整个面孔识别加工中发挥作用的。他们的结果显示, 相较于加工碎片面孔, 加工“Mooney”面孔时, Gamma振荡在初级视觉区、梭状皮层及顶叶皮层都有显著增强。并且, 在初级视觉皮层的Gamma振荡受到了来自顶叶皮层Gamma振荡神经活动的调控。这一证据清楚地展现了 Gamma振荡在自上而下的视觉信息整合加工中的作用。

图1 (a)和(b)单通道信息整合加工示例, (c)和(d)多通道信息整合加工示例
除了“Mooney”图象识别, “Kanizsa”图象(图形缺失部分能被识别为一个完整对象图形, 如图1b)识别也是探究 Gamma振荡与视觉信息整合加工关系的有效观测途径。Tallon-Baudry, Bertrand,Delpuech 和 Pernier (1996)很早就对“Kanizsa”图象识别与神经振荡变化的关系进行了探究。他们发现, “Kanizsa”三角图象的出现, 引发了Gamma振荡的增强, 而不能构成”Kanizsa”错觉的图象未使Gamma振荡增强。后续的研究支持了以上的结论;并且发现, 如果被试的注意力从“Kanizsa”图形上离开, “Kanizsa”图形与非“Kanizsa”图形并不会激发显著的Gamma振荡差异(Herrmann & Mecklinger,2000)。这一发现近一步表明, “Kanizsa”图形造成的Gamma振荡增强, 反映了一种自上而下的信息整合加工。最近的一项脑损伤病人研究也提供了Gamma振荡与“Kanizsa” 图象识别有关的有力证据:作为典型的视觉整体加工缺失人群——精神分裂症病患, 并没有像正常人一样, 加工“Kanizsa”图形比加工非“Kanizsa”图形激发更大的 Gamma振荡(Wynn et al., 2015)。
除了错觉, 在辨识日常熟悉物品的脑电探究中, 研究者也发现了Gamma振荡与视觉信息整合加工有关的证据。比如, 在早期的事件相关电位(ERP)成分上, 加工真实存在的熟悉对象与无意义的不熟悉对象并无差异(两者物理属性相同)。但是, 加工熟悉对象引发了更强烈的 Gamma振荡(Busch, Herrmann, Müeller, Lenz, & Gruber, 2006)。值得注意的是, 这一结果的报告经过了两个实验室的独立验证。后续, 研究者通过Granger因果模型, 推衍出人们在加工熟悉物品时引发的Gamma振荡神经活动是双向的(既有自上而下也有自下而上), 而加工非熟悉物品只能激发单向的Gamma振荡神经活动(Supp, Schlöegl, Trujillo-Barreto, Müeller, & Gruber, 2007)。该研究较好地表明, 在对真实世界的熟悉物品进行加工时, 人们会依赖经验自上而下地对碎片化信息进行整合;而Gamma振荡与这种整合伴随而生。
2.1.2 听觉
在碎片听觉信息的整合加工中, Gamma 振荡也扮演着重要的角色。比如, 研究者发现, 相较于预期读到一组不符合音律规则的单词, 当被试预期读一组符合音律规则的单词时, 会激发更强的Gamma振荡。也就是说, 音律的知识会对语音、语调信息的整合产生影响, 并且Gamma振荡可能在其中发挥着作用(Eulitz & Hannemann, 2010)。除了知识, 研究人员还发现, Gamma振荡与期望造成的语音特征信息整合加工也有关(Schadow,Lenz, Dettler, Früend, & Herrmann, 2009)。在一项研究中, 主试记录了被试在听一系列由 6个正弦声调组成的声调序列时的脑电。这些声调序列中有一些具有连续的特点, 比如连续上升或连续下降; 另一些在第 4或 5个声调的地方打破了声调的连续性。具有连续性特点的声调序列能给被试带来很强的预期感受。结果显示, 具有连续特征的声调序列伴随着明显的Gamma振荡增加。这预示着 Gamma振荡在预期造成的听觉信息特征整合加工中发挥着作用。
以上研究发现提示, Gamma振荡的增强能与单通道信息的整合相伴而生。单通道的整合往往具有借助已有内在表征(概念)的特点(Tang, Wu, &Shen, 2016)。并且, 这种整合往往与高级认知皮层的Gamma振荡增强有关。也就是说, 高级认知皮层 Gamma振荡可能是“自上而下”信息整合加工的一种生理基础。当然, 这一结论仍有待后续更多的实证研究支持。
2.2 跨通道的信息整合与Gamma振荡
在生活中, 我们常常会同时面临着来自不同感觉通道的信息。我们需要把这些信息进行跨通道整合, 以形成完整的认知对象。已有大量的研究涉及到 Gamma振荡与跨通道信息整合的关系探索。
2.2.1 视觉与听觉的整合
跨通道信息整合加工中, 研究最多的是视听信息的整合。在听觉诱发视错觉(sound-induced visual illusion) 的大脑电生理研究中, 人们很早就发现了 Gamma振荡与视听信息整合加工有关。听觉诱发视错觉是指, 当一个光闪出现在两个间隔在 60到 100 ms的短促声音刺激之间时,被试常会产生有两个光闪的错觉(如图 1c)。Bhattacharya, Shams和Shimojo (2002)的研究显示,当听觉成功诱发了视错觉时, 会伴随视听通道信息的超叠交互现象(supra-additive, 多通道信息所产生的认知效应量超过所有单一通道信息所产生的效应量之和; 相关细节见Calvert, Campbell, &Brammer, 2000)以及增强的Gamma振荡。Mishra,Martinez, Sejnowski和 Hillyard (2007) 对“听觉诱发视错觉”的进一步探究显示:第2个声音激发听觉皮层后, 感觉联合皮层也被激发, 并伴随着视觉皮层Gamma振荡的突增, 进而导致了听觉诱发视错觉的发生。他们的结果表明, 跨感觉皮层的Gamma振荡同步化增强, 可能是听觉诱发视错觉的关键性大脑生理基础。
另一类有关 Gamma振荡在视听信息整合中作用的研究, 主要涉及语音识别中的视听同步化的现象。Kaiser, Hertrich, Ackermann和Lutzenberger(2006)发现, 当被试看到/pa/这个发音的口型却同时听到/ta/这个发音时, 他们可能会产生自己听到的是/pa/这个音的错觉。并且, 同步记录到的MEG信号显示, 这种由视觉对听觉产生影响而形成的错觉与中线枕叶Gamma振荡高度相关。更直接反映 Gamma振荡与语音加工中视听信息整合有关的证据来自 Doesburg, Emberson, Rahi, Cameron和Ward (2008)。他们探究了视听信息时间窗口同步化对语音整合加工的影响及其大脑反应模式。他们让被试看一段大约2秒长的语音视频并记录被试的大脑EEG信号。语音视频共有两类:一类是视觉信息比听觉信息快 30 ms (时间同步); 另一类是视觉信息比听觉信息快300 ms (时间不同步)。他们发现, 涉及视听信息时间同步化的脑网络非常庞大, 这些脑网络通过Gamma振荡同步化来实现暂时性的功能连接, 以实现信息整合。
2.2.2 触觉与视觉的整合
除了视听信息整合, 也有大量研究涉及Gamma振荡与触觉和视觉信息整合的关系。Bauer,Oostenveld和 Fries (2009) 记录了被试在加工视觉与触觉同步化刺激时的大脑 MEG信号。在实验中, 被试在看见视觉图形刺激的同时, 放在盲文触碰器的右手食指会有 50%概率收到触觉信号。被试任务为对视觉图形是否出现作出快速反应并用左手食指按键。结果显示, 与视觉信息同步化的触觉信号能促进被试在任务中的判别表现,并且这种反应的促进与大脑 Gamma振荡同步化活动高度相关。
与视听信息整合加工类似, 一些错觉现象也是观察 Gamma振荡与触觉和视觉信息整合之间关系的途径。比如, 视触联合双闪错觉(double flash illusion)现象。在双闪错觉研究中, 被试在90 ms内会接收到视觉及触觉的组合通道刺激(包括2次视觉与触觉联合刺激; 2次视觉刺激伴随1次触觉刺激, 2次触觉刺激伴随 1次视觉刺激)或单一通道刺激(2次或1次视觉/触觉刺激), 并需报告共感受到了几次触觉刺激。当出现2次触觉刺激及 1次视觉刺激时, 被试却报告只出现了 1次触觉刺激(另 1次触觉刺激被感知为视觉刺激),既认为双闪错觉发生。研究发现, 视触联合双闪错觉的发生与大脑枕区(视觉)及大脑体感区(触觉)的 Gamma振荡交互的同步振荡增强有关(Lange,Oostenveld, & Fries, 2011)。
另一种观察 Gamma振荡和视触双通道信息整合关系的研究场景为橡胶手错觉(rubber hand illusion)。Kanayama团队开展了一系列有关橡胶手错觉的大脑生理电活动研究。在研究中, 主试会把一只橡胶手放置在一箱子上, 同时要求被试把他们自己的手放在一个箱子里(如图 1d)。当被试自己的手被触碰时, 他们会同时看见橡胶手被碰触了相同的位置或不同的位置(中指或拇指)。EEG的结果显示, 电极点 Gamma频段的振荡同步化程度与橡胶手错觉体验强度具有正相关性(Kanayama, Sato, & Ohira, 2007)。他们后续的研究重复了以上的发现, 并且发现这种视触整合错觉发生是不需自主意识努力的, 与Gamma振荡具有强烈的伴随性关系(Kanayama & Ohira, 2009)。
可以看出, Gamma振荡与跨通道信息整合高度相关。进一步归纳可以发现, 这种跨通道信息整合的常常是自下而上的, 受限于一定的时间范围(Tang et al., 2016)。并且, 这种跨通道整合常与感觉皮层的Gamma频段神经振荡同步化有关。当然, 以上结论所依赖的证据绝大多数来自视听信息以及视触信息的整合加工研究。未来还可探究其它跨通道信息整合加工与Gamma振荡的关系。
3 Gamma振荡生理基础及特点与整合加工的联系
以上的研究表明, Gamma振荡确与人们的整合加工有关。那为何Gamma振荡会被大自然选择为“整合加工”的神经基础呢?这可能与它的生理基础及特点有关。
3.1 Gamma节律振荡的生理基础
Gamma神经振荡是一种频率在30~100 Hz的高频波, 存在于感觉、运动及高级认知功能皮层中(Fries, Nikolić, & Singer, 2007)。神经学家认为,Gamma节律发生的生理模型主要有两种:1)锥体神经元(E细胞)与中间神经元(I细胞)交互模型(PING), 2)中间神经元间(I细胞群)交互抑制模型(ING; Tiesinga & Sejnowski, 2009)。
PING模型描述了 E和 I细胞互动, 产生Gamma振荡环路的过程。当E细胞受到外部刺激后, 会产生动作电位, 进而激活快速放电 I细胞(放电频率为 20~80 Hz)。被激活的 I细胞反过来抑制 E细胞的放电。这种交互活动, 便形成了一个 E、I细胞放电振荡回路, 放电频率在 Gamma波段。
ING模型描述了I细胞间互动, 产生Gamma振荡环路的过程。在一个处于工作状态的 I细胞连接网络中, 当部分快速放电的 I细胞接受同一外部输入时, 这些细胞便以同一速率放电(频率大于40 Hz)并在突触释放GABA, 抑制这个网络中其余I细胞活动。被抑制的I细胞的放电进程因此被延缓, 且下一次放电的时间更接近抑制它放电的I细胞放电时间。当 GABA能衰退时, 被抑制的I细胞群会活跃并抑制之前放电的I细胞。这种I细胞网络的群体性活动, 便促成了Gamma振荡放电回路的形成。
3.2 Gamma振荡特点与“整合加工”的渊源
Gamma振荡作为神经元互动的产物, 具有两大特点:1)在大脑各皮层“普遍存在”, 2)环路的形成具有“时间动态性”。这两个特点使Gamma振荡具备了作为“整合加工”生理基础的预设条件。
首先, Gamma振荡在大脑各皮层“普遍存在”的特性使它拥有了成为“整合加工”生理基础的潜质。生物学家的研究显示, Gamma节律的神经振荡存在于蟑螂的附脊髓生理节律性神经网络结构(Schneider & Stengl, 2007), 以及软体动物的脑神经节(Kay, 2015)等原始脑功能区中。神经进化学者提出, 由于生物进化压力的作用, Gamma振荡作为一种基础神经元互动模式在新皮层中被保留了下来(Bosman et al., 2014; Michalareas et al.,2016)。因此, Gamma振荡作为一种神经元间的互动模式, 广泛地存在于新旧皮层中。这一“普遍存在”特性, 使 Gamma振荡具备了作为一种通用皮层“工作语言”的条件, 进而能在促进皮层间的交互, 实现“碎片信息”整合的任务。
其次, 根据Gamma振荡产生的生理基础不难发现, Gamma环路的形成是一个动态的过程。当两群细胞具有相同的内部 Gamma节律共振性时,这两群细胞将处于相同的可互动时间窗口下, 它们的突触连接有效性会得到增强; 当两组细胞群的内部Gamma节律共振性不同时, 它们将处于不同的可互动时间窗口下, 它们的互动性会被抑制(Womelsdorf et al., 2007)。也就是说, Gamma振荡能使皮层之间实现具有“自组织性”及“动态调节性”的信息交换。这种基于神经振荡所产生的 “暂时性功能连接关系”, 能保障“信息整合”加工所需的灵活性与动态性。
总之, Gamma振荡的生理特性使它与“整合加工”产生了潜在的紧密联系(Bosman & Aboitiz,2015)。
4 Gamma振荡在人类“整合加工”中的作用机理
生理层面所体现出的 Gamma振荡与整合加工的联系, 并没有清晰地表明 Gamma在单通道“自上而下”的信息整合和跨通道“自下而上”的信息整合是如何发挥作用的。下文将从多层级整合加工视角, 论述Gamma振荡在人类“整合加工”中具体的作用机理。
4.1 整合加工的多层级性
多层级结构被神经科学家们认为是大脑在执行信息整合加工时的核心模式特征之一(Ursino,Cuppini, & Magosso, 2014)。根据多感知加工互联模型(Driver & Noesselt, 2008), 碎片信息会从多个感觉通道流入; 然后, 在不同水平皮层上进行着复杂的动态整合。具体来说, 感受器接收到外部信号并把它传递给皮层下组织区域, 皮层下区域向前把信号转递给初级感觉皮层, 并受到这些皮层的反馈回路的影响。初级感觉皮层之间互相进行着信息交换, 同时将信号进一步向前传递给感觉联合皮层区域(如,丘脑, 后顶叶皮层), 并得到感觉联合皮层的反馈(自下而上的整合, 感知觉皮层)。额顶调控皮层区域(如, 腹外侧前额叶)对所有进入的信息进行自上而下的整合, 并通过反馈机制影响到感觉联合皮层及初级感觉皮层的信息整合(自上而下的整合, 高级认知皮层; 多层级的整合加工结构, 请参见图 2)。下文将基于多层级整合视角, 简介有关Gamma振荡在不同水平整合加工中作用的理论。
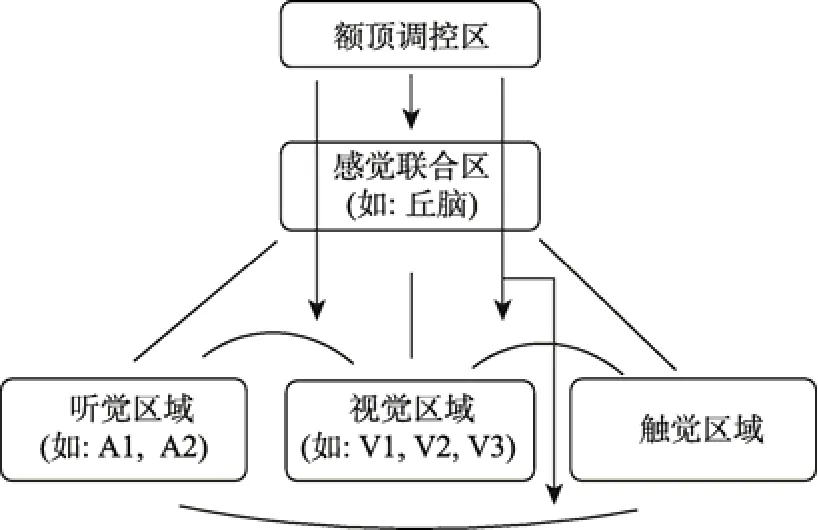
图2 整合加工的多层级结构
4.2 Gamma 振荡与多层级整合加工的相关理论解释
4.2.1 感知觉皮层的“信息整合”
发生在感知觉皮层的“自下而上”信息整合加工常与信息出现的时间同步性有关(Tang et al.,2016)。Womelsdorf等(2007)提出的神经耦合理论很好地预示了 Gamma振荡在这种知觉层面信息整合中的促进作用。他们认为, 当信号输入不同感觉皮层后, 这些皮层会连续将接收到的信号向前传递到下游神经元皮层。在这一过程中, 来自不同感觉通道的信号常会发生时间性耦合。这种耦合会引发关联性感觉皮层脑区和接受信号的感觉联合区产生相位相关的高频同步化放电, 也就是多脑区的Gamma同步化振荡, 从而促进跨通道的信息整合。
4.2.2 高级认知皮层的“信息整合”
匹配利用(MUM, match and utilization )模型(Herrmann, Munk, & Engel, 2004)较好地预示了Gamma振荡在高级认知皮层“自上而下”的信息整合中的促进作用。当个体因某些原因反复以一种固定模式对一客体对象进行认知加工时, 这种模式所对应的神经连接将经常被激活, 有关连接突触也将得到增强。当被试再次对有关客体对象进行认知操作时, 已形成的增强连接突触会激发更强的Gamma振荡, 从而促进他们对这一客体对象有关碎片信息的整合加工。也就是说, 个体会评估加工环境后, 形成加工对象的预期(match), 进而通过 Gamma 振荡有效地整合与预期对象有关的信息(utilization)。
可以看到, 不管是“自下而上” (感知觉皮层)还是“自上而下” (高级认知皮层)的信息整合加工,学者们都已提出了相关的理论模型加以解释。把这两者拼接在一起似乎就能构成一个比较完整的理论框架。但是, 这两种“整合加工”不是完全独立的系统, 存在一定的交互。那么在同时涉及多个水平整合加工时, 大脑是如何工作的?Gamma振荡在其中如何发挥作用?关于这一点, 现有理论还没有办法回答。我们将在展望中对这一重要问题进行一定的探讨。
5 总结
根据以上的论述, Gamma振荡无疑与整合加工密切相关。这一发现给了学者们两个方面的思考。第一, 既然Gamma振荡极有可能是整合加工的一种生理基础, 与整合加工有共生关系。那么是否意味着 Gamma振荡可以成为判断是否发生整合加工的重要指标呢?第二, 如前所述, 当下学者们对 Gamma振荡在简单背景整合加工中的作用已经有比较深入的了解, 那它在复杂情景整合加工中的作用是怎样的?下文将对这两个问题进行阐述与展望。
5.1 Gamma振荡在整合加工中的指标意义
根据上文所述, Gamma振荡与“信息整合加工”总是伴随性出现的。那么, 是否意味着“Gamma振荡”的出现是“信息整合加工”出现的充要条件。近期有学者对于这个问题进行了全面的探讨, 认为孤立地用 Gamma振荡来判断“信息整合加工是否发生”是不恰当的(Merker, 2013;Bosman et al., 2014)。综合现有研究发现, 我们觉得在使用 Gamma振荡作为判定“整合加工”是否发生的指标时应注意以下二点。
第一, 控制好刺激材料的物理属性。已有研究显示, 视觉刺激的大小和对比度(Perry, Hamandi,Brindley, Muthukumaraswamy, & Singh, 2013) 以及声音的响度(Ott, Stier, Herrmann, & Jäncke,2013)等刺激物理强度变化都会引起 Gamma振荡的变化。也就是说, 刺激材料物理特征会使刺激变得凸显, 引发更大的Gamma振荡。但这种振荡变化与整合加工无关。所以, 如果实验中刺激的物理属性差异不能被很好的控制, 研究者将很难推论 Gamma振荡的变化是由刺激材料本身引起的还是与整合加工有关, 从而造成对结果的错误判读(Rossiter, Worthen, Witton, Hall, & Furlong,2013)。
第二, 应尽量结合脑区信息来解读相应的发现。如前文所述, 不同类别的信息整合加工常常涉及到不同脑区的 Gamma振荡。比如, “自下而上”的整合加工常常与感觉皮层的Gamma同步化有关; “自上而下”的整合加工常常与额顶等高级皮层的 Gamma振荡增强有关。因此, 确定Gamma振荡变化发生的具体脑区对研究者解释相关结果会有很大帮助。Castelhano, Duarte, Wibral,Rodriguez和Castelo-Branco (2014)的研究就提供了一个案例。他们同时使用EEG及fMRI对被试在加工Mooney图象以及打碎了的Mooney图象时的神经信号进行记录。结果显示, 在视觉皮层出现了较高频段的Gamma振荡(60 Hz)。由此, 他们推断这项任务涉及“自下而上”的整合加工。
5.2 Gamma振荡在复杂情景整合加工中的作用
如前所述, 学者们已经对Gamma振荡在简单情景整合加工中的作用有了比较深入的认识, 但它在复杂情景整合加工中起到什么作用尚不清楚。这将是未来研究的一个很重要的方向。
5.2.1 Gamma振荡在多个水平的信息整合中的作用
日常生活中, 信息常以复杂的形式出现, 需要多水平的信息整合加工。所以, 一个亟待回答的问题是, Gamma振荡在这种多水平的信息整合中是如何发挥作用的?这一问题近期已开始得到学者们的关注。比如, Kanayama, Tamè, Ohira和Pavani (2012)探究了预期下视觉与触觉信息整合加工的神经振荡特点。他们发现同时出现的一致性视觉与触觉信息能促进个体的反应, 但这一促进会受到可预测性(高概率80% vs.低概率20%)的调节。他们还发现, 在知觉阶段的整合加工中, 视觉及触觉皮层 Gamma振荡有较大的增强, 表明Gamma振荡在感觉信息整合中起了作用; 对高概率事件的预测性加工与颞叶及前额区的 Gamma振荡增强有关, 表明Gamma振荡可能同样在高级认知水平的信息整合加工中发挥着作用。另一项有关预期条件下视听信息整合加工的研究也给出了类似的结果。在视听信息整合加工中, 视觉与听觉皮层的 Gamma振荡同步增强; 而人们在有预期的情况下进行跨通道信息整合加工时, 额顶区及视听感觉皮层的 Gamma振荡会增强 (Friese et al., 2016)。以上两项研究表明,在复杂的多水平信息整合中, Gamma振荡可能会分饰两个角色,以完成促进感知觉水平及高级认知水平的信息整合。但是, 相关研究还太少。不清楚在更复杂的多水平整合加工中, Gamma振荡会起怎样的作用及是否会承担新的角色。
5.2.2 Gamma振荡与社会信息整合加工
另一种复杂的信息整合加工情景为社会信息的整合加工。一个放在研究者面前的问题是, 社会信息整合所依赖的生理基础是否还是 Gamma振荡呢?近期, 一些来自社会情绪信息整合的研究显示, Gamma振荡可能仍是社会信息整合的生理基础之一。比如, Hagan等人(2009)发现, 当人们同时加工具有恐惧情绪的语调与面孔表情信息时, 会在颞上沟(重要视听联合区)出现神经振荡的超叠交互效应(效应解释见 2.2.1部分), 并且这种效应主要由Theta和Gamma振荡所构成。当人们同时加工具有中性情绪的语调与面孔表情信息时, 就不会在颞上沟出现神经振荡超叠交互效应。他们后续的研究显示, 以上的超叠交互效应在加工厌恶表情时也会发生, 并且Gamma振荡在这种超叠交互效应中发挥着主导性的角色(Hagan,Woods, Johnson, Green, & Young, 2013)。但是, 也有研究显示, 相比于单通道情绪信息加工, 当相关的视觉与听觉的情绪信息被同时加工时, 在Theta波段而非Gamma波段出现了神经振荡的超叠交互效应, 并且这种神经超叠交互效应与行为上的超叠交互效应高度相关(Chen et al., 2016)。所以, 关于Gamma振荡在社会情绪整合中的作用还不明确, 需更多的研究去回答这一问题。
Bauer, M., Oostenveld, R., & Fries, P. (2009). Tactile stimulation accelerates behavioral responses to visual stimuli through enhancement of occipital gamma-band activity.Vision Research, 49(9), 931–942.
Bhattacharya, J., Shams, L., & Shimojo, S. (2002).Sound-induced illusory flash perception: Role of gamma band responses.Neuroreport, 13(14), 1727–1730.
Bosman, C. A., & Aboitiz, F. (2015). Functional constraints in the evolution of brain circuits.Frontiers in Neuroscience,9, 303.
Bosman, C. A., Lansink, C. S., & Pennartz, C. M. A. (2014).Functions of gamma-band synchronization in cognition:from single circuits to functional diversity across cortical and subcortical systems.European Journal of Neuroscience,39(11), 1982–1999.
Busch, N. A., Herrmann, C. S., Müeller, M. M., Lenz, D., &Gruber, T. (2006). A cross-laboratory study of eventrelated gamma activity in a standard object recognition paradigm.NeuroImage, 33(4), 1169–1177.
Castelhano, J., Duarte, I. C., Wibral, M., Rodriguez, E., &Castelo-Branco, M. (2014). The dual facet of gamma oscillations: Separate visual and decision making circuits as revealed by simultaneous EEG/fMRI.Human Brain Mapping, 35(10), 5219–5235.
Castelhano, J., Rebola, J., Leitão, B., Rodriguez, E., &Castelo-Branco, M. (2013). To perceive or not perceive:The role of gamma-band activity in signaling object percepts.PLoS One, 8(6), e66363.
Calvert, G. A., Campbell, R., & Brammer, M. J. (2000).Evidence from functional magnetic resonance imaging of crossmodal binding in the human heteromodal cortex.Current Biology, 10(11), 649–57.
Chen, X. H., Pan, Z. H., Wang, P., Yang, X. H., Liu, P., You,X. Q., & Yuan, J. J. (2016). The integration of facial and vocal cues during emotional change perception: EEG markers.Social Cognitive and Affective Neuroscience,11(7), 1152–1161.
Doesburg, S. M., Emberson, L. L., Rahi, A., Cameron, D., &Ward, L. M. (2008). Asynchrony from synchrony:Long-range gamma-band neural synchrony accompanies perception of audiovisual speech asynchrony.Experimental Brain Research, 185(1), 11–20.
Driver, J., & Noesselt, T. (2008). Multisensory interplay reveals crossmodal influences on ‘sensory-specific’ brain regions, neural responses, and judgments.Neuron, 57(1),11–23.
Eulitz, C., & Hannemann, R. (2010). On the matching of top-down knowledge with sensory input in the perception of ambiguous speech.BMC Neuroscience,11, 67.
Fries, P., Nikolić, D., & Singer, W. (2007). The gamma cycle.Trends in Neurosciences, 30(7), 309–316.
Friese, U., Daume, J., Göeschl, F., König, P., Wang, P., &Engel, A. K. (2016). Oscillatory brain activity during multisensory attention reflects activation, disinhibition,and cognitive control.Scientific Reports, 6, 32775.
Hagan, C. C., Woods, W., Johnson, S., Calder, A. J., Green,G. G. R., & Young, A. W. (2009). MEG demonstrates a supra-additive response to facial and vocal emotion in the right superior temporal sulcus.Proceedings ofthe National Academy of Sciences of the United States of America, 106(47), 20010–20015.
Hagan, C. C., Woods, W., Johnson, S., Green, G. G. R., &Young, A. W. (2013). Involvement of right STS in audio-visual integration for affective speech demonstrated using MEG.PLoS One, 8(8), e70648.
Herrmann, C. S., Früend, I., & Lenz, D. (2010). Human gamma-band activity: A review on cognitive and behavioral correlates and network models.Neuroscience& Biobehavioral Reviews, 34(7), 981–992.
Herrmann, C. S., & Mecklinger, A. (2000).Magnetoencephalographic responses to illusory figures:early evoked gamma is affected by processing of stimulus features.International Journal of Psychophysiology, 38(3),265–281.
Herrmann, C. S., Munk, M. H. J., & Engel, A. K. (2004).Cognitive functions of gamma-band activity: Memory match and utilization.Trends in Cognitive Sciences, 8(8),347–355.
Kaiser, J., Hertrich, I., Ackermann, H., & Lutzenberger, W.(2006). Gamma-band activity over early sensory areas predicts detection of changes in audiovisual speech stimuli.NeuroImage, 30(4), 1376–1382.
Kanayama, N., & Ohira, H. (2009). Multisensory processing and neural oscillatory responses: Separation of visuotactile congruency effect and corresponding electroencephalogram activities.Neuroreport, 20(3), 289–293.
Kanayama, N., Sato, A., & Ohira, H. (2007). Crossmodal effect with rubber hand illusion and gamma-band activity.Psychophysiology, 44(3), 392–402.
Kanayama, N., Tamè, L., Ohira, H., & Pavani, F. (2012). Top down influence on visuo-tactile interaction modulates neural oscillatory responses.NeuroImage, 59(4), 3406–3417.
Kay, L. M. (2015). Olfactory system oscillations across phyla.Current Opinion in Neurobiology, 31, 141–147.
Keil, A., Müller, M. M., Ray, W. J., Gruber, T., & Elbert, T.(1999). Human gamma band activity and perception of a gestalt.The Journal of Neuroscience, 19(16), 7152–7161.
Klemen, J., & Chambers, C. D. (2012). Current perspectives and methods in studying neural mechanisms of multisensory interactions.Neuroscience & Biobehavioral Reviews, 36(1), 111–133.
Lange, J., Oostenveld, R., & Fries, P. (2011). Perception of the touch-induced visual double-flash illusion correlates with changes of rhythmic neuronal activity in human visual and somatosensory areas.NeuroImage, 54(2), 1395–1405.
Merker, B. (2013). Cortical gamma oscillations: The functional key is activation, not cognition.Neuroscience and Biobehavioral Reviews, 37(3), 401–417.
Michalareas, G., Vezoli, J., van Pelt, S., Schoffelen, J.-M.,Kennedy, H., & Fries, P. (2016). Alpha-beta and gamma rhythms subserve feedback and feedforward influences among human visual cortical areas.Neuron, 89(2), 384–397.
Mishra, J., Martinez, A., Sejnowski, T. J., & Hillyard, S. A.(2007). Early cross-modal interactions in auditory and visual cortex underlie a sound-induced visual illusion.Journal of Neuroscience, 27(15), 4120–4131.
Moratti, S., Méndez-Bértolo, C., Del-Pozo, F., & Strange, B.A. (2014). Dynamic gamma frequency feedback coupling between higher and lower order visual cortices underlies perceptual completion in humans.NeuroImage, 86,470–479.
Ott, C. G. M., Stier, C., Herrmann, C. S., & Jäncke, L. (2013).Musical expertise affects attention as reflected by auditory-evoked gamma-band activity in human EEG.Neuroreport, 24(9), 445–450.
Perry, G., Hamandi, K., Brindley, L. M., Muthukumaraswamy, S.D., & Singh, K. D. (2013). The properties of inducedgammaoscillations in human visual cortex show individual variability in their dependence on stimulus size.NeuroImage, 68, 83–92.
Rossiter, H. E., Worthen, S. F., Witton, C., Hall, S. D., &Furlong, P. L. (2013). Gamma oscillatory amplitude encodes stimulus intensity in primary somatosensory cortex.Frontiers in Human Neuroscience, 7, 362.
Schadow, J., Lenz, D., Dettler, N., Früend, I., & Herrmann, C.S. (2009). Early gamma-band responses reflect anticipatory top-down modulation in the auditory cortex.NeuroImage,47(2), 651–658.
Schneider, N.-L., & Stengl, M. (2007). Extracellular longterm recordings of the isolated accessory medulla, the circadian pacemaker center of the cockroach Leucophaea maderae, reveal ultradian and hint circadian rhythms.Journal of Comparative Physiology A, 193(1), 35–42.
Senkowski, D., Schneider, T. R., Foxe, J. J., & Engel, A. K.(2008). Crossmodal binding through neural coherence:Implications for multisensory processing.Trends in Neurosciences, 31(8), 401–409.
Supp, G. G., Schlögl, A., Trujillo-Barreto, N., Müeller, M. M.,& Gruber, T. (2007). Directed cortical information flow during human object recognition: Analyzing induced EEG gamma-band responses in brain’s source space.PLoS One,2(8), e684.
Tallon-Baudry, C., Bertrand, O., Delpuech, C., & Pernier, J.(1996). Stimulus specificity of phase-locked and non-phase-locked 40 Hz visual responses in human.The Journal of Neuroscience, 16(13), 4240–4249.
Tang, X. Y., Wu, J. L., & Shen, Y. (2016). The interactions of multisensory integration with endogenous and exogenous attention.Neuroscience & Biobehavioral Reviews, 61,208–224.
Tiesinga, P., & Sejnowski, T. J. (2009). Cortical enlightenment:Are attentional gamma oscillations driven by ING or PING?Neuron,63(6), 727–732.
Ursino, M., Cuppini, C., & Magosso, E. (2014).Neurocomputational approaches to modelling multisensory integration in the brain: A review.Neural Networks, 60,141–165.
Womelsdorf, T., Schoffelen, J.-M., Oostenveld, R., Singer,W., Desimone, R., Engel, A. K., & Fries, P. (2007).Modulation of neuronal interactions through neuronal synchronization.Science, 316(5831), 1609–1612.
Wynn, J. K., Roach, B. J., Lee, J., Horan, W. P., Ford, J. M.,Jimenez, A. M., & Green, M. F. (2015). EEG findings of reduced neural synchronization during visual integration in schizophrenia.PLoS One, 10(3), e0119849.
Yau, J. M., DeAngelis, G. C., & Angelaki, D. E. (2015).Dissecting neural circuits for multisensory integration and crossmodal processing.Philosophical Transactions of the Royal Society B: Biological Sciences, 370(1677), 20140203
