土壤理化性质对腐朽红松根部土壤微生物数量的影响
2018-03-20曹延珺徐华东王立海周涵婷刘华
曹延珺,徐华东,王立海,周涵婷,刘华
0 引言
土壤是森林生态系统的物质基础,为林分的生长提供水分和营养物质[1-2],还为各种微生物的生长繁殖提供了必需的养分、水分、渗透压、温度等条件,是微生物的“天然培养基”。土壤微生物作为土壤中最活跃的部分,在森林中扮演着分解者的角色,分解了90%以上的森林枯落物和动植物残体,在促进森林土壤物质分解和转化中有着不可替代的作用[3]。红松是世界珍稀树种,为我国二级保护植物,具有很高的经济价值和生态价值。近年来,随着天然红松林面积的减少和人工林面积的增加,红松林内的腐朽等病害不断增加,而红松具有较长的生长周期,更新过程缓慢而复杂,因此,对红松进行合理的保护对原始红松林的维护具有重要的意义。目前关于红松的研究较多,Sun Fan[4]研究了松梢象对红松主枝和侧枝挥发性组分的影响,徐华东[5]对冻结红松木材内部的水分状况进行了相关研究,孙天用[6]对红松活立木腐朽的土壤影响因子进行了研究,而且已有学者对红松根部土壤微生物群落结构进行了研究[7]。但有关腐朽红松活立木根部土壤微生物数量变化与土壤理化性质的研究还比较缺乏。孙天用[8]研究发现,红松活立木干基腐朽与土壤理化性质之间存在着密切的关系。而土壤理化性质、地形和季节等条件与土壤微生物的数量和分布密切相关[9]。因此本文从研究腐朽红松活立木根部土壤微生物特征的角度出发,通过分析土壤三大类群微生物数量分布特征及其与土壤理化性质之间的关系,以期为红松活立木的保护提供新思路。
1 材料和方法
1.1 研究样地概况
凉水国家自然保护区地理坐标为东经128°53′20″,北纬 47°10′50″,位于黑龙江省伊春市小兴安岭南坡,共划分为31个林班,研究样地为第18林班。林班内的地形主要为山谷和山麓,地势较为平缓,海拔高度为300~500m,年均气温1.4℃,属寒温带大陆性季风气候。林班内的主要土壤类型为暗棕壤中的浅育暗棕壤,红松为优势树种,伴生有冷杉、水曲柳、榆树和椴树等树种,林分郁闭度在0.5~0.8之间。
1.2 试验材料
2016年5月,在凉水国家自然保护区第18林班内,采用随机法选取22棵存在腐朽的红松活立木,除去样木根部地表的植被和枯落物,选取地势平坦的位置作为取样点,将土钻垂直插入土壤中往下钻取,直到土钻20cm刻度处,取出土钻,将土钻中的土样装入无菌乙烯袋中密封,迅速带回实验室,置于4℃冰箱保存,用于微生物的培养计数。在土钻取样的位置,用铁锹挖一个V字型的土坑,深度20cm,在土坑内用铁锹倾斜向下切取一片土壤,然后从上到下取一部分土片装入铝盒密封编号记录,用于土壤含水率的测定,其余部分带回实验室自然风干后,过2mm筛用于土壤养分的测定。
1.3 试验方法
1.3.1 土壤理化性质的测定
对采回来的22个土样进行理化性质的测定。
土壤含水率的测定采用烘干法。将土样置于烘箱中(105℃)烘干至恒重。计算公式为:土壤含水率=(鲜土质量-烘干土质量)÷烘干土质量×100%[10]。
土壤全氮采用半微量氏法测定;土壤水解性氮采用碱解蒸馏法测定;土壤全钾采用氢氧化钠碱熔-火焰光度法测定;土壤速效钾采用乙酸铵浸提-火焰光度法测定;土壤全磷采用HClO4-H2SO4消煮法测定;土壤有效性磷采用盐酸-硫酸浸提法测定[11]。
1.3.2 土壤微生物培养计数
土壤微生物三大类群的数量测定采用稀释平板分离法进行。细菌采用牛肉膏蛋白胨培养基分离培养,真菌采用马丁氏培养基分离培养,放线菌采用高氏一号培养基分离培养。培养时间为细菌2d,真菌3d,放线菌7d,然后进行菌落计数。计数采用Interscience scan300全自动菌落计数器。计数后按照公式(1)[12],计算出每克鲜土中的菌落数。计算时,首先选择平均菌落数在30~300之间者进行。
每克样品中微生物活细胞数

1.4 数据统计分析
原始数据的处理、简单图表的绘制均采用Excel 2003软件;统计分析部分采用SPSS19.0统计软件进行处理。利用Pearson相关系数评价土壤微生物数量和土壤理化性质间的相关性,并通过逐步回归分析确定对微生物数量影响最显著的土壤因子。
2 结果与分析
2.1 土壤微生物三大类群数量分布规律
土壤微生物包括细菌、真菌、放线菌、藻类及原生动物等,类群十分丰富。细菌、真菌和放线菌这三大类群的数量在土壤微生物中占有绝对优势,因此是本文的主要研究对象。从三大类微生物数量分布(图1,主纵坐标轴表示细菌数量,次纵坐标轴表示放线菌和真菌数量)和所占的比例(表1)来看,细菌在土壤中的数量远大于放线菌和真菌,所占比例最高达99.98%,其次为放线菌,真菌最少。且细菌数量比放线菌和真菌高出了2~3个数量级,与森林土壤微生物相关研究结果一致[13-14]。这是由于细菌个体小、代谢强,其繁殖方式主要为裂殖,细胞以2n的速度进行增殖,繁殖十分迅速,世代时间短,因此为土壤微生物中的主要类群[15]。土壤放线菌数量仅次于细菌,占总菌数比例最高达5.59%。许多放线菌分解纤维素、石蜡、橡胶等物质的能力极强,因此它们在提高土壤肥力方面起着重大作用。真菌数量最少,但在土壤质量改善方面有着不可替代的贡献,具有降解植物主要成分(纤维素、果胶、木质素等)的能力。
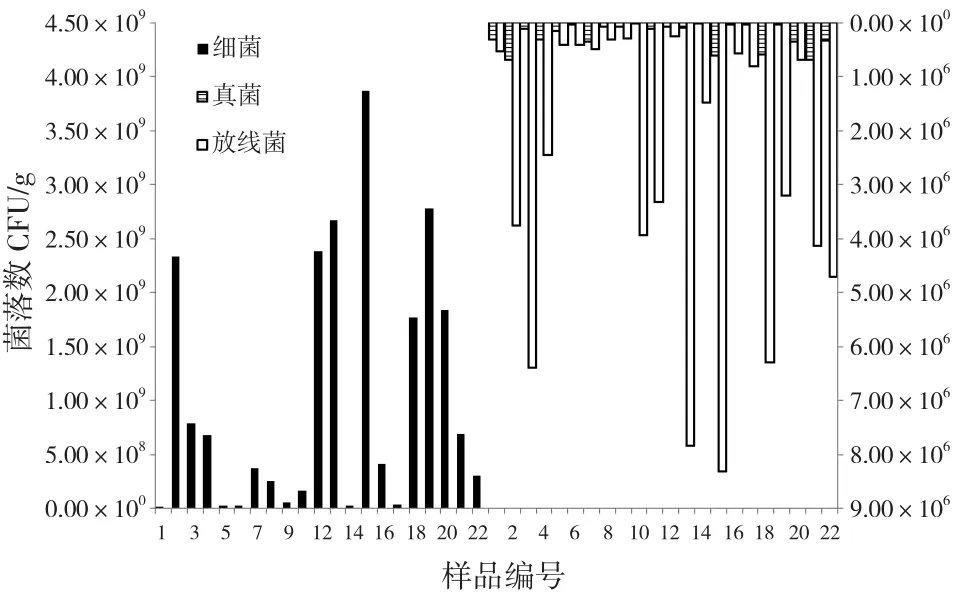
图1 土壤微生物数量分布Fig.1 Quantity distribution of microorganisms
2.2 土壤微生物和理化性质的关系
2.2.1 Person相关性分析
表2为22个土样的理化指标计算结果。由表2可知,腐朽红松活立木下,土壤全N含量较高,而土壤全N的92%~98%都贮藏在有机质中,不能直接被植物利用;全K和全P含量相对林伯群[16]对凉水自然保护区第16林班浅育暗棕壤理化分析结果较低,这可能与调查时间差异和微生物作用有关。N、P、K元素主要是以水解N 、速效P和速效K的方式被林木吸收利用,而它们的矿化分解作用与很多因素有关。从表2 中可以看出,腐朽红松活立木下,土壤速效P含量很低,速效K含量也较低。P元素在土壤中的行为较为复杂,微生物矿化分解作用的有效范围也比较窄。土壤速效K水热耦合效应研究表明,土壤速效K含量会随着土壤温度的升高和含水率的增加而增加[17-18]。水热条件可能是导致本实验速效K含量较低的重要原因。
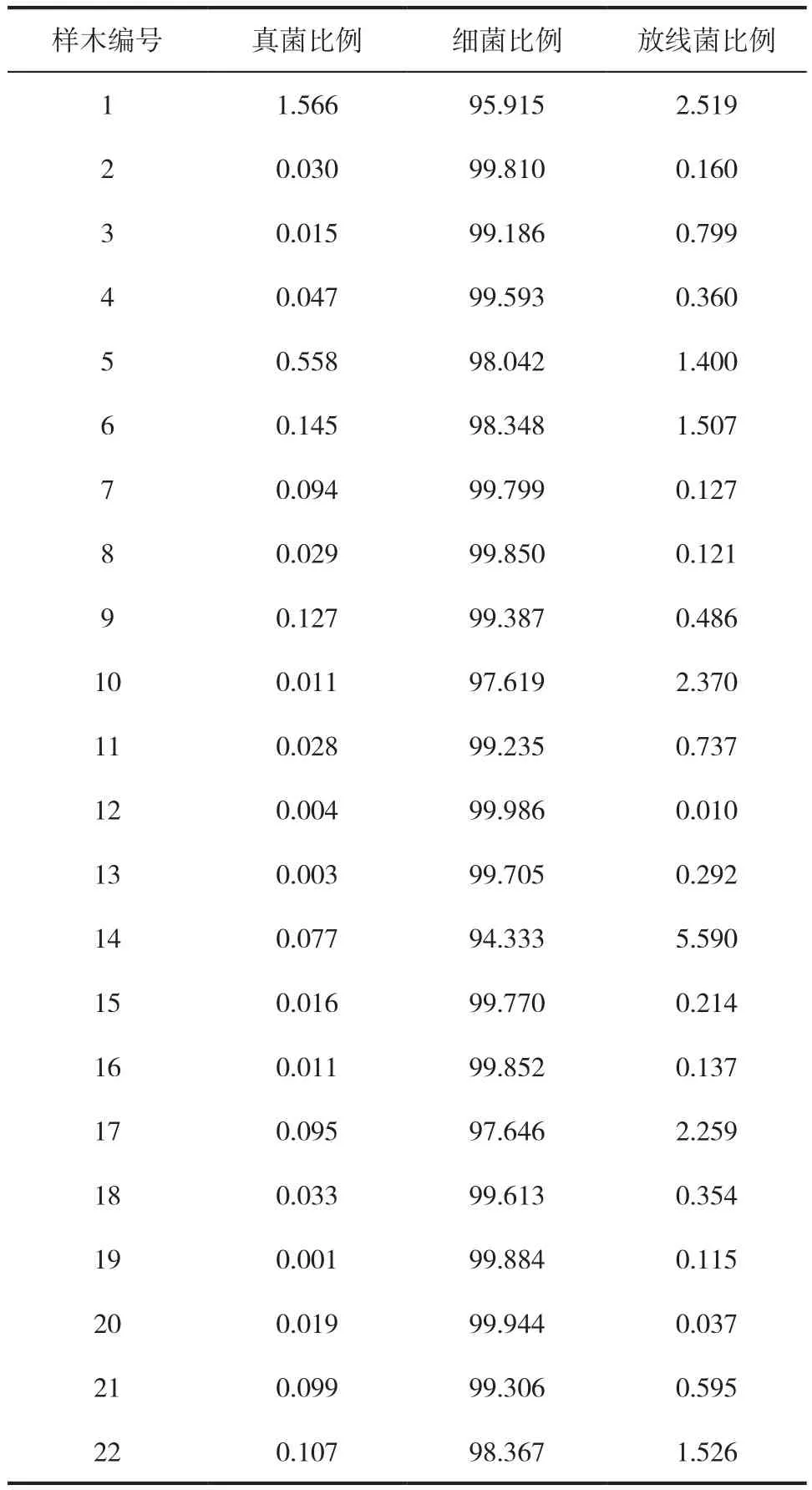
表1 土壤微生物数量比例 %Tab.1 Quantitative proportion of microorganisms
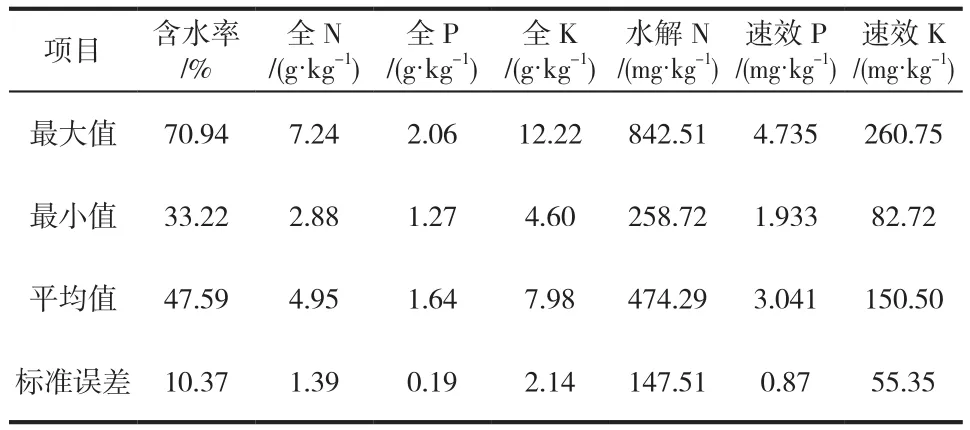
表2 土壤理化指标Tab.2 Soil properties
表3为土壤细菌、真菌和放线菌与土壤理化指标的相关性分析。由表3可以看出,土壤真菌数量与全N呈极显著正相关,与全K呈显著正相关,与含水率、全P、水解N、速效P和速效K没有显著的相关性。其中,真菌数量与土壤全N在置信度为99%时呈高度正相关(R=0.672)。这是由于土壤真菌大多为腐生型,必须从土壤中的有机物中获取碳源、氮源和能量,通过降解土壤中结构复杂的氮化物,提供自身生长繁殖所需要的物质,也为植物的生长提供能源。土壤含水率与真菌数量没有明显的相关性。贾倩民[19]对干旱区盐碱地土壤理化性质与微生物数量关系的研究发现,土壤真菌数量与含水率呈正相关,戴雅婷[20]在油蒿根际土壤微生物数量与土壤养分关系的研究中发现,土壤真菌数量与含水率呈负相关,赵萌[21]在杉木人工林地土壤微生物数量与土壤因子关系的研究中发现真菌数量与土壤含水率不具有相关性。因此,地表植被、土壤类型、采样时间等条件不同,研究结果也不同。
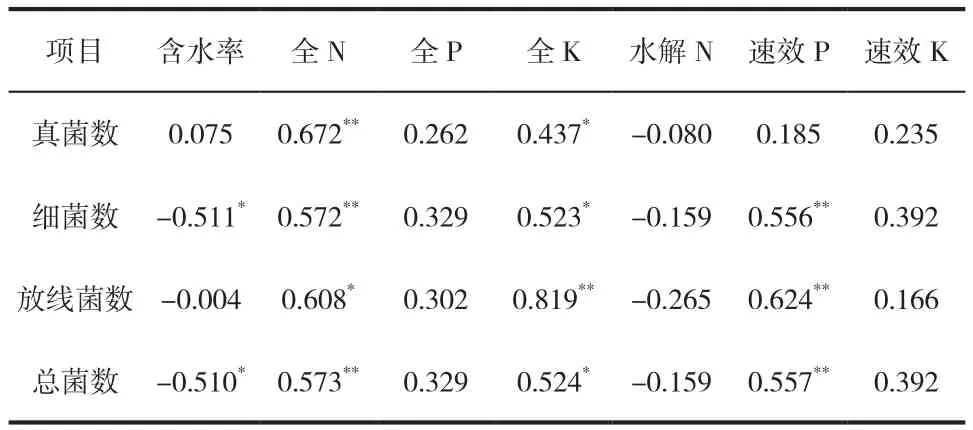
表3 土壤微生物数量与土壤理化指标相关性Tab.3 Correlation between microorganisms quantity and soil properties
土壤细菌数量与全N和速效P呈极显著正相关,与全K呈显著正相关,与含水率呈显著负相关,与全P、水解N和速效K没有显著相关性。
土壤细菌数量与含水率呈显著负相关,这与细菌喜湿的生长习性不符。张崇绑[22]在研究羊草草原土壤细菌数量动态与生态因子之间的关系时发现,土壤含水率、温度、速效钾和水解氮与细菌数量呈正相关。其中,土壤温度对细菌数量变化的影响最大(R=0.99),与其他三个因子相比,含水率对细菌数量的影响最小。由于采样时间为5月,样地内部分土壤尚未完全解冻,土壤温度偏低,且土壤通气性较差,使得相同含水率条件下土壤中细菌数量与常温时相差较大,这有可能是导致细菌数量与土壤含水率的关系发生变化的原因,有待进一步研究。
放线菌数量与全K和速效P呈极显著正相关,与全N呈显著正相关,与含水率、全P、水解N和速效K无显著相关性。其中,放线菌数量与土壤全K含量在置信度为99%时呈高度正相关(R=0.819)。放线菌适宜生长在含水量较低、有机物较丰富的土壤中,土壤中的钾对放线菌菌丝的生长和抗生素的产生具有不可或缺的作用[23]。
总菌数(土壤三大类微生物数量的总和)与土壤理化性质的相关性和细菌与土壤理化性相关性一样。这是由于土壤细菌数量所占总菌数比例均在94%以上,是土壤活动力的主力军。
2.2.2 回归分析
为了进一步确定影响微生物数量的主要土壤因子,分别以土壤真菌、细菌、放线菌和微生物总数作为因变量,与微生物数量存在线性关系的土壤全N、全K、速效P作为自变量,采用逐步回归法进行多元线性回归分析并建立回归方程。由于土壤含水率受温度影响,因此进行回归分析时直接剔除。得出的回归方程如下:
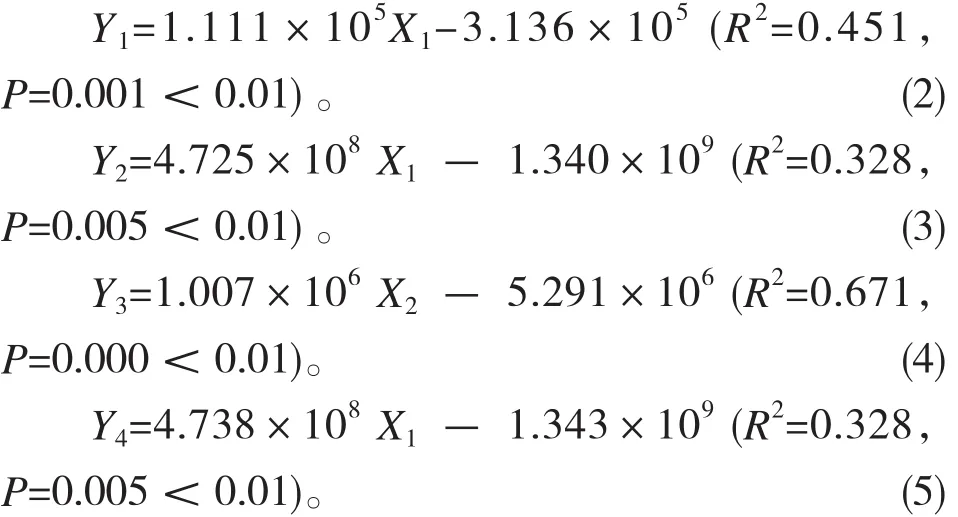
其中,Y1~Y4依次表示土壤真菌、细菌、放线菌数量和总菌数,X1~X3依次表示土壤全N、全K、速效P。
回归结果表明,红松根部土壤真菌、细菌数量和微生物总数主要受土壤全N含量的影响,放线菌数量主要受土壤全K含量的影响。土壤全N含量对真菌数量变化的贡献率为45.1%,对细菌和微生物总数变化的贡献率为32.8%,土壤全K含量对放线菌数量变化的贡献率为67.1%,这说明除了全N和全K,红松根部土壤微生物数量还受土壤温度、碳含量、地形条件等多种因素的影响。由于土样采自腐朽红松根部,因此也可能与红松的健康状况有关,还需要进一步研究。
3 结论
(1)腐朽红松活立木根部土壤三大类群微生物数量分布规律为细菌>放线菌>真菌。
(2)腐朽红松活立木根部的土壤真菌、细菌数量和微生物总数主要受土壤全N含量的影响,放线菌数量主要受土壤全K含量的影响。
土壤微生物作为土壤物质转化和能量流动的主要推动者,在一定程度上反映了土壤的肥力状况,从而对地表植被的生长产生影响。本文对腐朽红松根部土壤微生物数量分布等情况进行了初步研究,而影响微生物数量的因素还有很多,有待今后继续研究,同时要更加深入的研究植被健康水平和微生物的关系,为红松针阔混交林的可持续经营提供可靠的依据。
[1]王晓莉, 常禹, 陈宏伟, 等.黑龙江省大兴安岭主要森林生态系统生物量分配特征[J].生态学杂志, 2014, 33(6):1437-1444.
[2]向泽宇, 张莉, 张全发, 等.青海不同林分类型土壤养分与微生物功能多样性[J].林业科学, 2014, 50(4):22-31.
[3]Kirk J L, Beaudette L A, Hart M, et al. Methods of studying soil microbial diversity[J]. Journal of Microbiological Methods,2004, 58(2): 169-188.
[4]Sun F, Chi D, Yu J, et al. Relationship between volatiles from Korean pine and damage by Pissodes nitidus[J]. Journal of Northeast Forestry University, 2010, 38(1): 108-109.
[5]徐华东, 王立海.冻结红松和大青杨湿木材内部水分存在状态及含量测定[J].林业科学, 2012, 48(2):139 - 143.
[6]孙天用, 王立海, 侯捷建, 等.木材含水率和地形条件对红松活立木腐朽程度的影响[J].应用生态学报, 2015, 26(2):349 - 355.
[7]Liang S, Liu T, Li L, et al. Comprehensive evaluation model of korean pine microbial community structure in xiaoxing'an mountains[J]. Forest Engineering, 2014, 30(4) : 42-45.
[8]孙天用.小兴安岭针阔混交林立地条件对红松树干腐朽的影响[D].哈尔滨:东北林业大学, 2015.
[9]王伟东.微生物学[M].北京:中国农业大学出版社, 2015.
[10]耿增超.土壤学[M].北京:科学出版社, 2011.
[11]毛波, 董希斌. 大兴安岭低质山杨林改造效果的综合评价[J].东北林业大学学报, 2016, 44(8) : 7-12.
[12]尹军霞.微生物学实验指导[M].南京:南京大学出版社,2015.
[13]王轶.原始红松林土壤微生物可培养类群动态变化[D].哈尔滨:东北林业大学, 2008.
[14]王素英, 王海荣, 方亮.大青山不同立地条件林木根际土壤微生物数量变化研究[J].内蒙古林业科技, 2009,35(3):18-24.
[15]窦森.土壤有机质[M].北京:科学出版社, 2010.
[16]林伯群.森林土壤六十年[M].北京:科学出版社, 2010.
[17]张一平.土壤养分热力学[M].北京:科学出版社, 2010.
[18]张甜, 朱玉杰, 董希斌. 小兴安岭用材林土壤肥力综合评价及评价方法比较[J]. 东北林业大学学报, 2016, 44(12):10-14.
[19]贾倩民, 陈彦云, 刘秉儒, 等.干旱区盐碱地不同栽培草地土壤理化性质及微生物数量[J].草业科学, 2014, 31(7):1218-1225.
[20]戴雅婷, 闫志坚, 王慧, 等.油蒿根际土壤微生物数量及其与土壤养分的关系[J]. 中国草地学报, 2012, 34(2) : 71-75.[21]赵萌, 方晰, 田大伦.第2代杉木人工林地土壤微生物数量与土壤因子的关系[J]. 林业科学, 2007, 43(6) : 7-12.
[22]张崇绑.羊草草原土壤细菌数量动态与生态因子之间关系的研究[J].微生物学通报, 2001, 28(2) : 1-4.
[23]陈克龙, 刁治民.草地土壤微生物学[M].北京:经济科学出版社, 2016.
