红松人工林和相邻次生林林下红松天然更新种群格局分析
2018-03-20王琴香沈海龙和春庭NguyenThanhTuan
王琴香,沈海龙,*,和春庭,Nguyen Thanh Tuan
0 引言
红松(Pinus koraiensis)是温带地区地带性顶极群落阔叶红松林的建群种,其天然更新种群状况对群落的稳定发展具有重要意义。因此,红松人工更新一直是研究的重点[1],利用天然更新来维持和恢复重建阔叶红松混交林也一直是森林经营管理的关注重点[2-18]。从20世纪50年代中后期开始,东北林区营造了大量红松人工林,这些红松人工林通常与周围不同类型的天然次生林呈镶嵌分布。
自然条件下,植物实现成功更新必须具备三个要素:种源、传播和萌发幼苗[19],以及适合幼苗生长发育的微生境,如林隙[20]。目前,很多红松人工林已经达到结实盛期,可以提供丰富的种源。红松种子传播需要依靠动物的藏食习性,而对更新有利的动物主要是松鼠和星鸦。松鼠饱食后,会把松子埋入土壤中,通常每簇种子数2~4粒[6,21],最多达 18 粒[6]。星鸦颈部舌下囊可容纳大量松子,胃和舌下囊松子数量可达30~50粒[7]。正常条件下,红松人工林及其相邻次生林内均不缺乏传播红松种子的动物,种子传播有保障。但是,近年来人为球果采摘强度很高,导致红松更新的种源日渐减少[2-3]。这不仅直接减少了天然更新的种子来源,而且间接增加了种子传播动物对埋藏种子的消耗,从而减少了地下种子库的种源。在高强度球果采摘的条件下,红松人工林及其周围次生林中的红松天然更新是否会受影响?其受影响的程度如何?这些问题尚不清楚。
本研究以48 a红松人工林及其相邻蒙古栎(Quercus mongolica)次生林为对象,研究两种林分内天然更新红松幼苗、幼树种群的特点,分析相邻的红松人工林和次生林林下红松天然更新种群格局的特点和差异及其与球果采摘强度的关系,旨在为红松天然更新的调控提供科学依据,并为红松球果采摘强度的控制决策提供参考。
1 研究方法
1.1 研究地概况
调查样地位于吉林省敦化市寒葱岭林场(43°00´~43°12´N,127°55´~128°05´E), 该 林 场属于温带大陆性湿润季风气候,冬季干燥寒冷,夏季温暖湿润,年平均气温2.6℃,极端最高气温可达33.4℃,极端最低气温可达38.3℃,无霜期120 d左右,年平均降水量约621.4 mm。红松人工林样地设置在12林班46小班的红松人工林,样地坡向西,坡度13°,土壤为典型暗棕壤;林龄48 a,平均树高15.5 m,胸径20.4 cm,平均郁闭度0.7;草本层平均盖度40%,平均高为20 cm。蒙古栎次生林样地设置在同一林班的51小班,样地坡向北,坡度14°,郁闭度0.7,土壤为典型暗棕壤;平均年龄59 a,平均树高16.2 m,平均胸径14.6 cm;下木高度1.6 m,盖度30%;草本层平均高度30 cm,盖度40%。
1.2 调查方法
考虑到红松幼苗矮小不易发现,为了保证数据的精确度,选择了草木还没完全覆盖地面的六月份开展调查。在紧邻红松人工林的蒙古栎次生林林分内设置15个400 m2的样带(长×宽=20 m×20 m)调查次生林内红松天然更新,在红松人工林内的三个坡位上分别设置700 m2的三个条形样带(长×宽=70 m×10 m)。测量样地内所有红松幼苗幼树的年龄、轮枝数、株/簇数、高度、第一活枝高以及生长情况。幼苗幼树高度使用卷尺测量,测量精度为0.1 cm,地径使用电子游标卡尺测量。并记录样地内上层木的树种、树高、胸径和草本层的种类、盖度和平均高度。
2 结果与分析
2.1 红松人工林和蒙古栎次生林林内红松天然更新种群的年龄结构
由图1(a)可知,在蒙古栎次生林中,红松天然更新种群密度随年龄变化出现两个峰值,第一个峰值在1a时出现,更新密度达到515株/hm2,然后随年龄增加天然更新种群密度一直在下降,到14a时下降的趋势变为增加的趋势,到22 a时出现第二个峰值,更新密度达到32株/hm2。由图1(b)可知,与蒙古栎次生林相邻,并为其林分内红松更新提供种源的红松人工林内红松天然更新种群年龄结构不完整,最大只存在4 a更新苗,1 a更新苗的密度高达6 228株/hm2,以后随苗龄增加,更新密度下降剧烈,2 a更新苗的密度为132株/hm2,3 a更新苗的密度为456株/hm2,4 a更新苗的密度已低至7株/hm2。
2.2 红松人工林和蒙古栎次生林1-3a红松天然更新幼苗分布情况
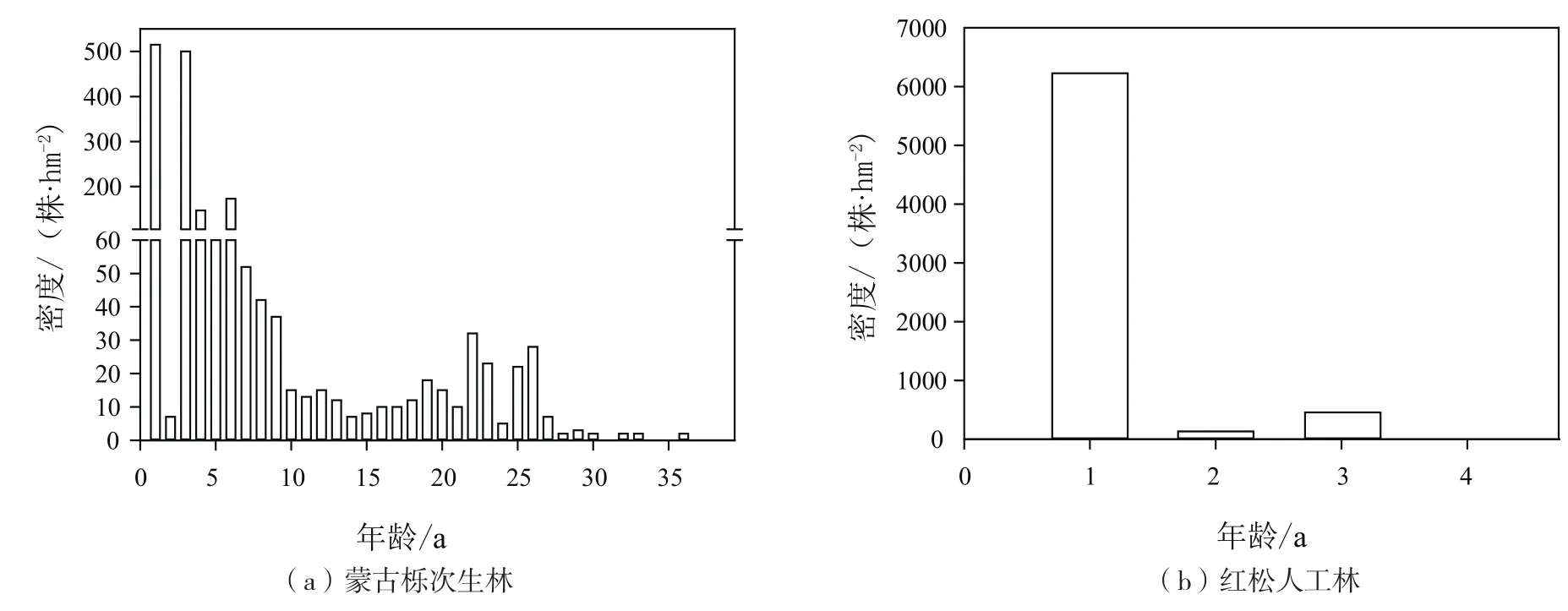
图1 蒙古栎次生林和红松人工林林内天然更新红松种群年龄结构Fig.1 Age structures of Korean pine natural regenerated seedlings in Mongolian oak secondary forest and Korean pine plantation
由图2可知,红松人工林和蒙古栎次生林两种林分中,红松天然更新1~3 a幼苗的分布不同于以往的“山”状分布—以簇生为主,而是以单株分布为主,随株/簇的增加,簇数呈指数下降,分布图表现为倒“J”形。在红松人工林林内更新苗簇数与株数的相关系数介于0.935 4~0.998 2,其中2 a苗的相关系数最大,达到0.998 2。蒙古栎次生林内更新苗簇数与株数的相关系数介于0.926 6~1,其中2 a苗的相关系数高达1。由图2(a)和图2(d)可知,1 a更新苗在红松人工林和蒙古栎次生林两种林分中的总簇数分别是1 746簇和157簇,人工林内1 a苗簇数约是次生林的11倍;由图2(b)和图2(e)可知,2 a更新苗在两种林分中的总簇数分别是63簇和7簇,人工林内2 a苗簇数是次生林的9倍;由图2(c)和图2(f)可知,3 a更新苗在两种林分中的总簇数分别是187簇和203簇,二者数量接近,比值约为1。考虑到自然条件下红松种子要经过两个冬天才能打破休眠而萌发,结合红松种子的丰欠年,1 a更新苗(产生种子时间:2014年,平年)、2 a更新苗(产生种子时间:2013年,欠年)、3 a更新苗(产生种子时间:2012年,丰年),结合图2可知,在丰年,球果采摘对不同林分类型的影响差别不大,次生林内也有相当多的种子,而在平年和欠年,球果采摘严重影响了种源附近林分的红松天然更新,差别能高达9~11倍。
2.3 红松人工林和蒙古栎次生林林内1~3a红松天然更新幼生长情况
由图3可知,在红松人工林中红松天然更新1,2,3 a幼苗高度分别为6.02、7.56、9.67 cm,地径分别为1.35、1.18、1.55 mm,高度表现为增长趋势,地径为先下降后增长。在蒙古栎次生林中红松天然更新1,2,3 a幼苗高度分别为7、8.2、12.24 cm,地径分别为:1.86、1.94、2.72 mm,高度和地径都表现出增加趋势。总的来说,蒙古栎次生林中红松天然更新幼苗生长情况优于红松人工林,蒙古栎次生林中1 a和2 a红松幼苗的高度大于红松人工林中相同年龄的更新苗,但差异不显著(p>0.05),而3 a更新苗的高度却极显著大于红松人工林中的更新苗(p<0.001)。在自然环境下,两种林分内1 a、2 a到3 a红松天然更新幼苗的地径之间差异均极显著(p<0.001),并且两种林分内红松天然更新苗地径差异随着苗龄增加而变得越来越大(1 a更新苗差值为0.51 mm,2 a更新苗差值为0.76 mm,3 a更新苗差值为1.17 mm)。
3 结论和讨论
种群年龄结构分析是揭示种群现状和更新的重要途径之一[22],一个健康发展的种群,其年龄结构往往呈现倒J形,幼苗数量最多,随年龄增长数量不断减少[23]。而在本次研究调查中,蒙古栎次生林和红松人工林内红松天然更新种群年龄结构皆不是健康发展种群的倒J形。蒙古栎次生林中两个更新峰值的出现代表了两种球果采摘强度下的红松天然更新情况,峰值分别出现在1 a和22 a,种群年龄结构表现为先下降后上升的趋势;在14 a的时候由下降转为上升,原因可能是以14a是两个不同采摘强度(强度一:丰年采摘,欠年不采;强度二:丰年、欠年全采)的分界点。结合寒葱岭当地的球果承包采摘历史,应该是近21年来采摘严重,采摘方式是丰欠年全树全采,种源缺少了导致苗龄<22 a的红松幼苗数量的不足;而22年前(1994年)松子价格低廉,球果采摘只集中在丰年,所以1994年以前红松种群是健康发展的,现在种群结构的不完整的主要原因是近21年来不断增强的球果采摘造成的。人工林有大量的萌发幼苗,但这些幼苗在第三年后死亡率几乎达到100%,具体原因尚待研究。
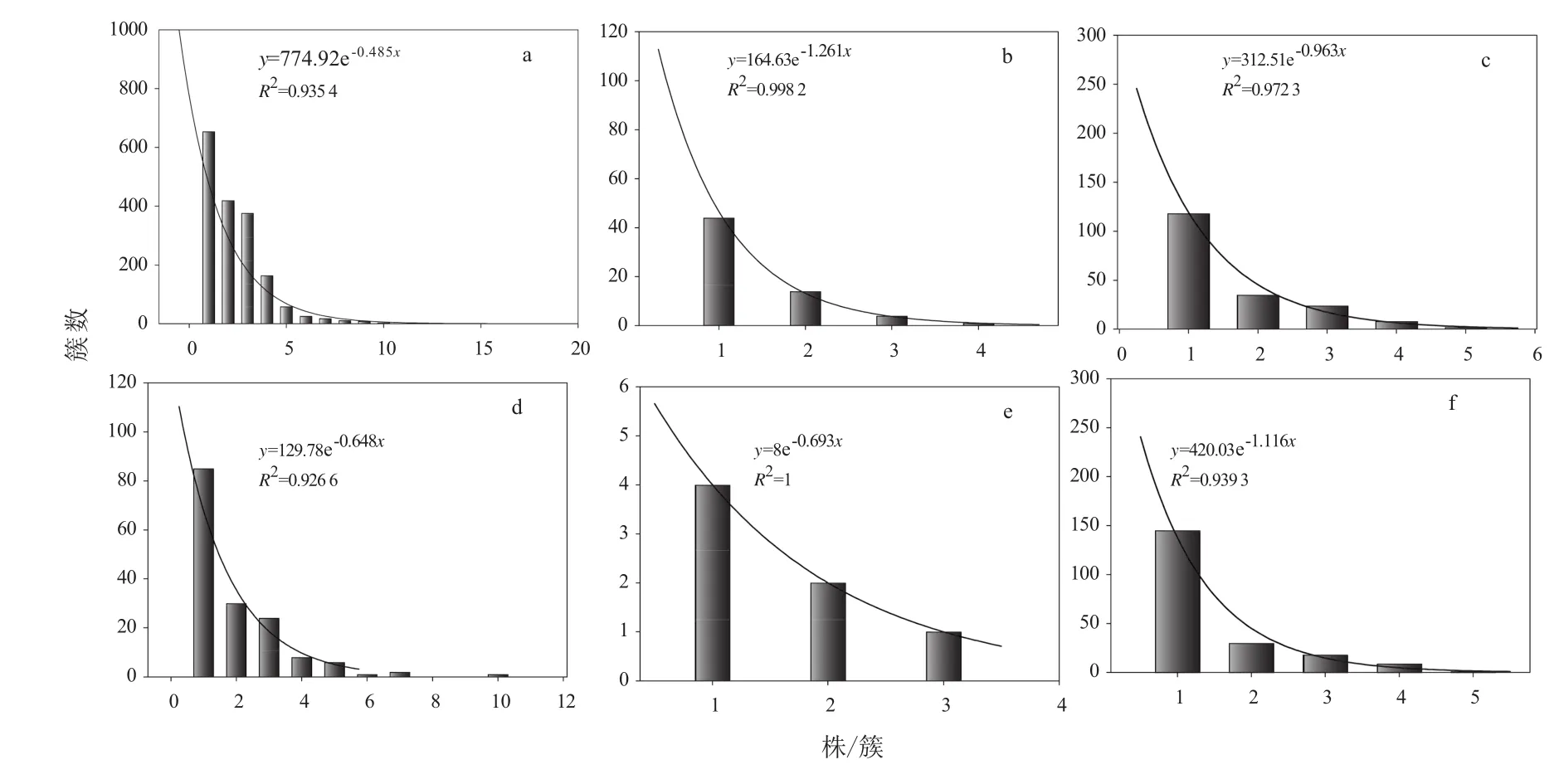
图2 红松人工林和蒙古栎次生林林内1~3 a幼苗的株/簇分布Fig.2 The distribution of Korean pine 1 to 3 year old seedlings regenerated in Korean pine plantation and Mongolian oak secondary forest
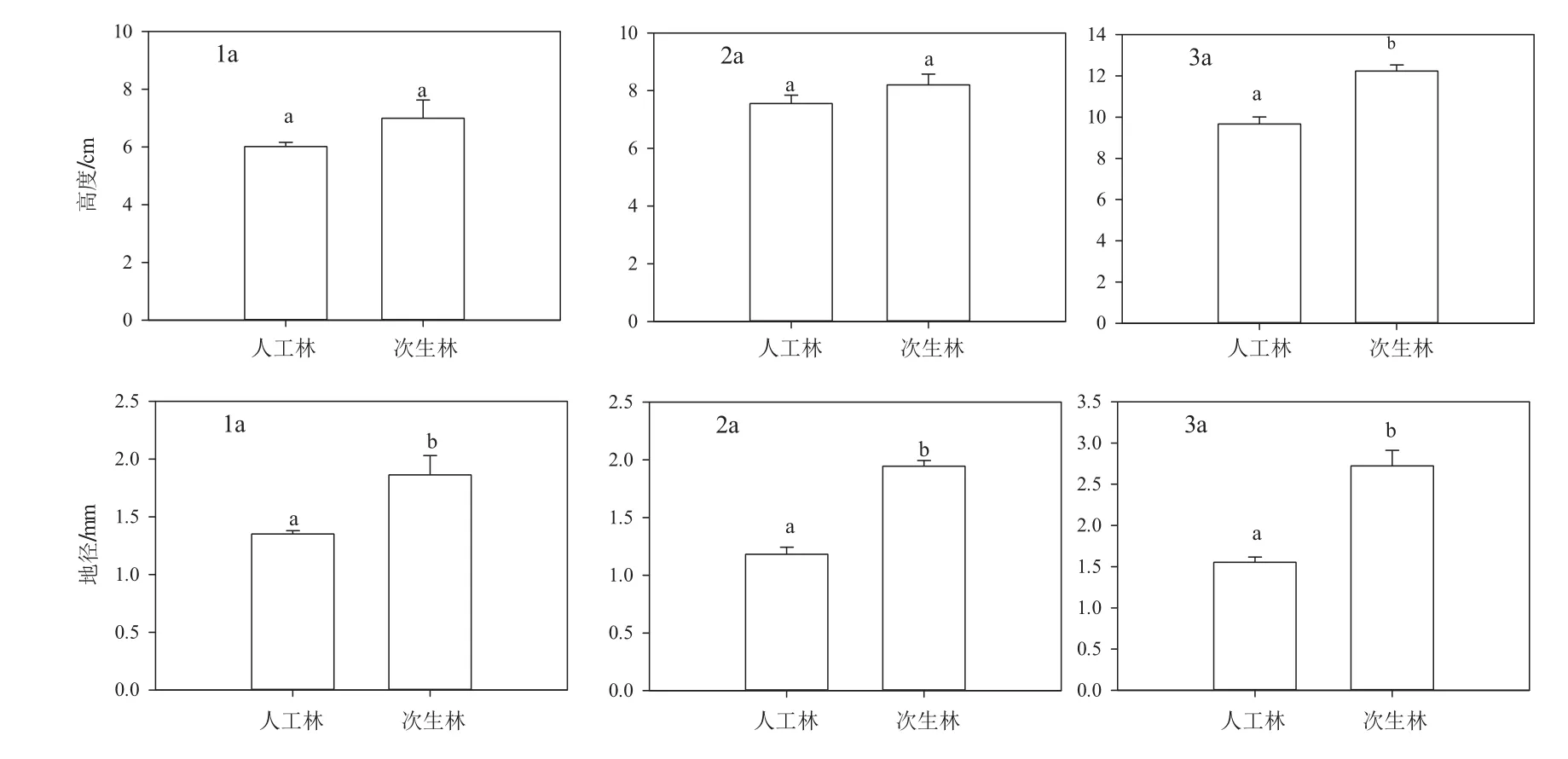
图3 蒙古栎次生林和红松人工林内天然更新红松幼苗苗高和地径生长状态Fig.3 The height and diameter growth of Korean pine seedlings natural regenerated in Mongolian oak secondary forest and Korean pine plantation
根据红松更新与传播动物之间的联系,松子以多粒被埋藏于土壤中,据上世纪80年代松子埋藏情况调查,单粒占全部埋藏种子数的9.7%,2~4粒 /簇占 60.3%,5~7粒 /簇占 16.6%,8~10粒 /簇占 5.7%,10~20粒 /簇占 7.6%[24]。由前人研究可知,埋藏种子/簇的分布情况呈山状,峰值出现在2~4粒/簇。而一年生幼苗直接来自于种子库,所以1 a萌发幼苗分布与种子库种子分布相似,只是萌发幼苗的数量少于种子的数量[25]。因而,1 a幼苗株/簇与簇数之间理论上应表现出“山”状分布(由1 a转化的2 a苗和3 a的分布情况也同理),幼苗多以2~4株/簇分布,单株和4株以上分布逐渐减少。然而在本研究中,不论是人工林、次生林,还是1、2、3 a苗,幼苗都以单株存在为主,随每簇株树的增加,簇数呈指数下降,分布图表现为倒“J”形。高的单株率的形成有以下两方面原因,一是高的动物遇见率,球果采摘后,捕食者在冬天食物不足时,就会多次重复取食,使得种子多呈单粒存在[3];二是自然环境的影响,幼苗生命力脆弱存活不易,死亡率很高。红松幼年耐荫且适当遮阴也有利于幼苗的生长[9-12],即使在郁闭度达到0.8~0.9的柞树红松林中天然更新也很好[13]。而次生林不同光照条件下红松3a幼苗栽植实验证明,4 a幼苗可在树冠开度7%以上生长[14]。这说明,自然环境影响应该不是导致这种分布状态的主要原因。高强度球果采摘导致的种子库种源不足、动物取食种子和幼苗频度大大增加,可能是主要的原因。
有研究用红松人工林下土壤和枯落叶凋落物浸提液培育红松种子,结果表明,人工林下浸提液抑制了红松幼苗的生长[26]。在本次研究中,蒙古栎次生林中红松天然更新幼苗生长情况优于红松人工林,比较二者可知,蒙古栎次生林的林内环境有利于红松天然更新幼苗生长,红松人工林内的环境不利于红松天然更新幼苗的生长,更新幼苗生长缓慢,长势越来越弱直至死亡,这是红松人工林内没有更大年龄更新苗的一个原因。
[1]冯志忠, 刘云洲, 宋海玉, 等. 阔叶红松林择伐强度及更新技术的研究[J]. 森林工程, 2001, 17(2): 1-2, 16.
[2]刘足根, 姬兰柱, 郝占庆, 等. 松果采摘对长白山自然保护区红松天然更新的影响[J]. 应用生态学报, 2004, 15(6): 958-962.
[3]金光泽, 杨桂燕, 马建章, 等. 松果采摘对小兴安岭主要林型红松土壤种子库和幼苗库的影响[J]. 自然资源学报, 2010,25(11): 1845-1854.
[4]鲁长虎. 动物与红松天然更新关系的研究综述[J]. 生态学杂志, 2003, 22(1): 49-53.
[5]陶大立, 赵大昌. 红松天然更新对动物的依赖性—一个排除动物影响的球果发芽实验[J]. 生物多样性, 1995,3(3): 131-133.
[6]鲁长虎, 刘伯文, 吴建平. 阔叶红松林中星鸦和松鼠对红松种子的取食和传播[J]. 东北林业大学学报, 2001, 29(5): 96-98.
[7]赵锡如. 星鸦与红松更新关系[J]. 林业实用技术, 1987 (9): 32.
[8]王树力, 葛剑平, 徐继成, 等. 小兴安岭杨桦林下红松种群天然更新的格局与过程[J]. 东北林业大学学报, 1993,21(5):7-13.
[9]王树力, 武敬辉, 史永纯. 红松种群天然更新及幼年生长与林分结构关系的研究[J]. 吉林林学院学报, 1998, 14 (1): 6-10.
[10]教士奇, 刘大兴, 聞殿墀. 紅松闊叶树混交林的采伐方式与更新方法[J]. 林业科学, 1966, 11(2): 106-113.
[11]朱济凡, 刘慎谔, 王战, 等. 小兴安岭红松针阔叶混交林[J].林业科学, 1958, 4(4): 355-369.
[12]金鑫, 胡万良, 丁磊, 等. 遮阴对红松幼苗生长及光合特性的影响[J]. 东北林业大学学报, 2009, 37(9): 12-13.
[13]徐振邦, 代力民, 陈吉泉, 等.长白山红松阔叶混交林森林天然更新条件的研究[J].生态学报, 2001, 21(9): 1413-1420.
[14]Sun Y R, Zhu J J, Sun O J, et al. Photosynthetic and growth responses of Pinus koraiensis, seedlings to canopy openness:Implications for the restoration of mixed-broadleaved Korean pine forests[J]. Environmental & Experimental Botany, 2016,129: 118-126.
[15]刘宏伟,王国义.红松果林营建与发展的几点建议[J].林业科技, 2016, 41(4) : 52-54.
[16]孙浩然,孟庆彬,张巍.小兴安岭林区红松不同家系子代生活力与出苗率测定[J].林业科技, 2016, 41(3) : 7-9.
[17]郭树平. 小兴安岭阔叶红松林林分结构与修复[J]. 森林工程, 2014, 30(6) : 8-12.
[18]郑颖.不同林龄红松优树结实的经济效益分析[J].林业科技, 2016, 41(2) : 44-45.
[19]肖治术, 张知彬, 王玉山. 以种子为繁殖体的植物更新模型研究[J]. 生态学杂志, 2003, 22(4): 70-75.
[20]于金莹, 亢新刚. 林隙影响天然林更新研究进展[J]. 森林工程, 2005, 21(5): 1-4.
[21]Hayashida M. Seed dispersal by red squirrels and subsequent establishment of Korean pine[J]. Forest Ecology &Management, 1989, 28(2):115-129.
[22]Brotherson J D, Rushforth S R, Evenson W E, et al. Population dynamics and age relationships of 8 tree species in Navajo National Monument, Arizona[J]. Journal of Range Management, 1983,36(2):173-196.
[23]梁士楚, 李久林, 程仕泽. 贵州青岩油杉种群年龄结构和动态的研究[J]. 应用生态学报, 2002, 13(1): 21-26.
[24]刘庆洪. 红松阔叶林中红松种子的分布及更新[J].植物生态学与地植物学学报.1988, 12(2):134-142.
[25]刘庆洪. 小兴安岭红松种群天然更新的特点[J].林业科学,1987, 23(3): 265-276.
[26]陈立新, 李少博, 乔璐, 等. 凋落物叶和土壤浸提液对红松种子萌发及幼苗生长的影响[J].南京林业大学学报(自然科学版).2016, 40(2): 81-87.
