复合植物精油对脂多糖刺激仔猪肝脏的保护作用
2018-03-20汪中兴吴梦郡纪昌正徐海旺侯永清
李 鹏 ,汪中兴 ,吴梦郡 ,余 魁 ,纪昌正 ,徐海旺 ,易 丹 ,2,王 蕾 ,2,赵 迪 ,2,侯永清 ,2*
(1.武汉轻工大学动物营养与饲料科学湖北省重点实验室,湖北武汉 430023;2.武汉轻工大学动物营养与饲料安全湖北省协同创新中心,湖北武汉 430023)
脂多糖(Lipopolysaccharide,LPS)是革兰氏阴性菌细胞壁的主要成分之一,进入动物体内会引发免疫应激,导致大量自由基产生,造成肠道、肝脏等组织损伤。LPS刺激仔猪被认为是模拟应激的经典模型[1]。近年来,由于抗生素滥用导致的耐药菌及抗生素残留问题对公共健康构成严重威胁,开发抗生素替代品成为当前动物营养学界的研究热点。作为天然植物提取物,植物精油逐渐受到人们的青睐[2]。
肉桂油(Cimmmomum Oil)是樟树科植物肉桂的干皮及树皮经水蒸气蒸馏得到的挥发油,其主要成分为肉桂醛,对细菌、酵母、霉菌的生长有较强的抑制作用[3]。牛至油(Origanum Oil)为牛至的提取物,其主要成分为香芹酚、百里酚等,可抑制和杀灭多种微生物[4]。百里香油(Thymus Oil)为百里香的提取物,其主要成分为百里香酚、香芹酚,被欧盟批准为植保产品,具有抗菌和抗氧化等功效等[5-6]。肝脏是机体新陈代谢的主要场所之一,同时也是体内的主要解毒和免疫防御器官。饲养环境中的致病性和非致病性抗原会刺激仔猪肝脏巨噬细胞产生并释放大量炎性介质,进而造成肝脏结构和功能损伤[7]。因此,探究3种精油的复合物对仔猪肝脏在外界抗原刺激下的保护作用,对保证肝脏的健康、维持机体正常的新陈代谢至关重要。
本课题组前期研究表明,日粮中添加复合植物精油(OCT)可以降低仔猪腹泻率,促进仔猪生长[8]。本试验通过建立仔猪LPS刺激模型,探究OCT对LPS刺激仔猪肝脏的保护作用,为开发绿色的添加剂产品提供理论基础。
1 材料与方法
1.1 试验设计 选取 18头体重为(7.34±0.62)kg的21日龄健康杜×长×大仔猪,随机分为对照组、LPS组、OCT+LPS组,每个处理组6个重复,每个重复1头猪。预试期4 d,正试期21 d。试验期间,对照组与LPS组饲喂基础日粮,OCT+LPS组在基础日粮中添加50 mg/kg OCT,OCT由牛至油、肉桂油和百里香油按质量比1:1:1混合配制而成。于试验第21天,LPS组与OCT+LPS组仔猪腹腔注射LPS (100 μg/kg 体重),对照组注射等量的生理盐水,3 h后前腔静脉采血,6 h后屠宰取肝脏,待测。
1.2 试验日粮与饲养管理 基础日粮配制参照NRC(2012)猪的营养需要,基础日粮的组成和营养成分见表1。猪舍为全封闭式,屋顶以排气扇通风,舍温维持在25~27℃,料槽为可调式,鸭嘴式饮水器自动供水。进栏前清洗猪舍并消毒。每笼1头试验猪,每天饲喂5次,猪舍定期清扫和消毒。
1.3 测定指标与方法
1.3.1 血浆生化指标 于试验第21天注射LPS 后3 h,对每头猪前腔静脉采血,用2 mL的EDTA-Na抗凝真空采血管收集1.5 mL血液。将血样于冰上静置15 min,离心(3 000 r/min,10 min)分离血浆,采用日立7020全自动血浆生化分析仪,采用德赛诊断系统(上海)有限公司的试剂盒测定谷草转氨酶(AST)、谷丙转氨酶(ALT)、碱性磷酸酶(ALP)、谷酰转肽酶(GGT)。
1.3.2 肝脏组织抗氧化指标 于试验第21天注射LPS后6 h,对每头猪进行解剖取样,取肝脏,置于冰上剪碎,采用锡箔纸包好放入纱布袋投入液氮中速冻,然后将肝脏样品转至盛有液氮的研钵中研磨成粉末,分装。迅速转移至-80℃冰箱冻存。将肝脏组织从-80℃冰箱中取出,称取0.5 g左右,迅速置于预冷的玻璃匀浆器中,加入9倍体积的生理盐水,在冰浴环境下匀浆,然后离心(4℃,3 000 r/min,15 min),取上清液。采用南京建成试剂盒检测髓过氧化物酶(MPO)、一氧化氮合酶(T-NOS)、诱导型一氧化氮合酶(i-NOS)。
1.3.2 肝脏相关基因mRNA水平的检测 用RPL4基因做 内 参, 检 测HSP70、EGF、IL-10、IGF-1、mTOR基因的相对表达量,相关基因的引物序列见表2。总RNA的提取和cDNA的合成均采用宝生物工程(大连)有限公司RNAiso Plus(Total RNA提取试剂)、PrimeScript®RT reagent kit with gDNA eraser(cDNA合成试剂盒)和SYBR®Premix Ex TaqTM(Tli RNaseH Plus)(Real time PCR试剂盒)操作完成。参照Fu等[9]方法(2-△△CT法)统计分析各基因qPCR的数据结果。
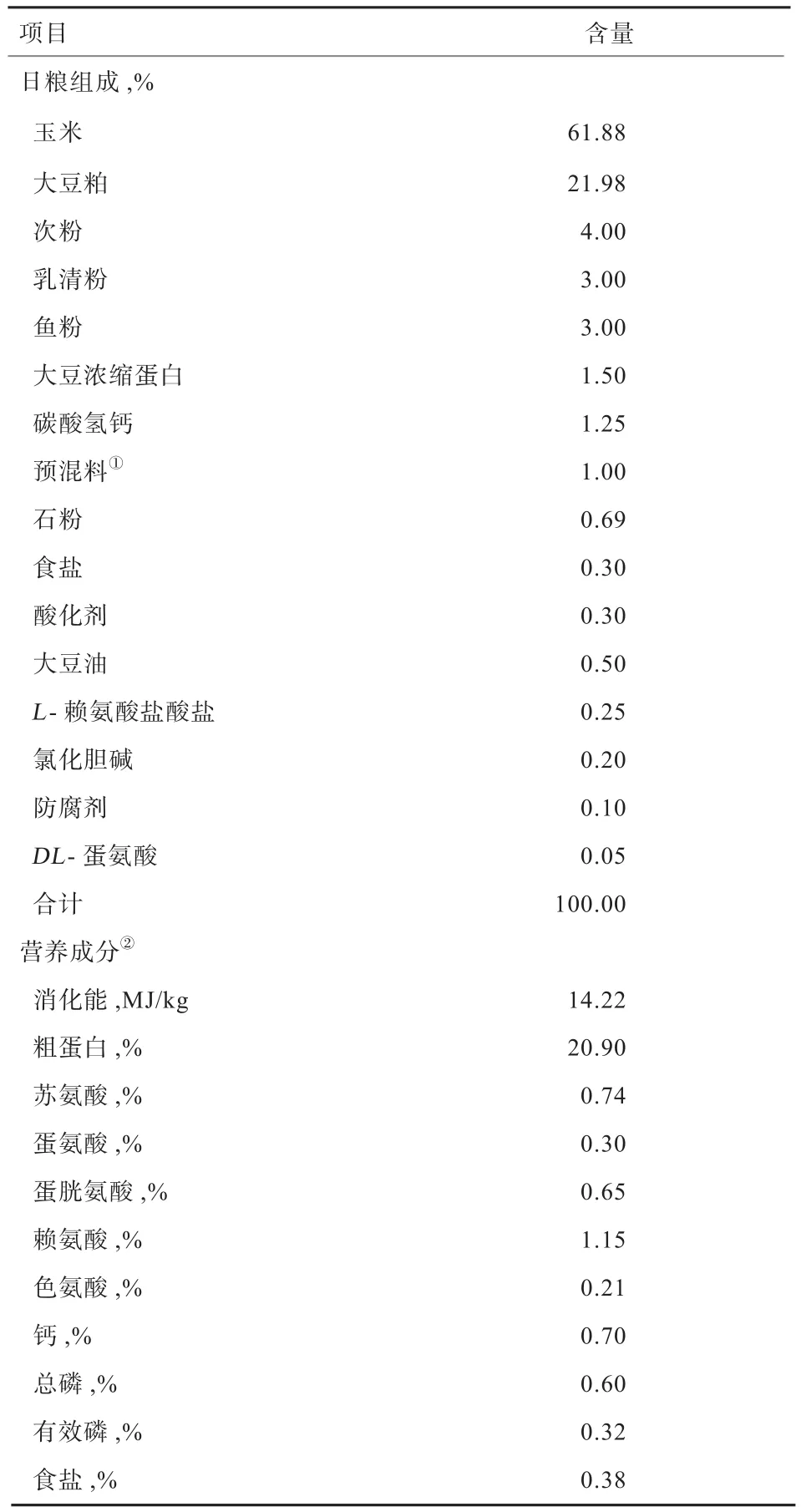
表1 基础日粮组成及营养成分(风干基础)
1.4 引物的设计合成 利用Primer premier 6.0软件设计,由宝生物工程(大连)有限公司合成。各基因引物序列见表2。
1.5 统计分析 试验采用SPSS 17.0统计软件进行单因素方差分析和Duncan's多重比较,结果用平均值±标准差来表示。以P<0.05表示差异显著,以0.05<P<0.10表示具有显著性趋势。

表2 基因的引物序列表
2 结 果
2.1 OCT对LPS刺激仔猪血液生化指标的影响 由表3可知,与对照组相比,LPS刺激显著提高了仔猪血浆ALT、AST、ALP、GGT的活力(P<0.05),而日粮中添加50 mg/kg的OCT可显著缓解LPS导致的仔猪血浆ALT、AST、GGT活力的升高(P<0.05)。
2.2 OCT对LPS刺激仔猪肝脏MPO、T-NOS和i-NOS含量的影响 由表4可知,与对照组相比,LPS刺激显著提高了仔猪肝脏i-NOS的活力(P<0.05),且有升高肝脏MPO活力的趋势(P<0.1);日粮中添加50 mg/kg的OCT可缓解LPS导致的仔猪肝脏i-NOS活力的升高(P<0.05),且有缓解LPS导致的仔猪肝脏MPO活力升高的趋势(P<0.1)。
2.3 肝脏相关基因mRNA水平 由表5可知,与对照组相比,LPS刺激显著上调了仔猪肝脏HSP70基因的相对表达量(P<0.05),下调了仔猪肝脏EGF、IL-10、IGF-1、mTOR基因的相对表达量(P<0.05)。日粮中添加50 mg/kg的OCT可缓解LPS导致的仔猪肝脏HSP70基因相对表达量的升高以及EGF、IL-10、IGF-1、mTOR等基因相对表达量的降低(P<0.05)。
3 讨 论
功能完整的肠黏膜可对细菌和内毒素构成屏障作用,但是在LPS刺激后肠黏膜受损,大量细菌和毒素经门静脉和肠系膜淋巴系统,进入人体血液循环引起全身炎症反应综合征,导致内脏器官的损伤[10]。ALT和AST是血液常规检测指标之一,它们是蛋白质代谢的重要酶类,也是常用的评价肝脏损伤的指标。当机体正常时,血液中ALT与AST含量很低,肝脏受损时,血液中的ALT与AST活性会显著升高[11-12]。肠道疾病、自身免疫疾病及恶性肿瘤患者的ALP含量会显著升高[13],GGT是由肝细胞产生,并且在原发性肝癌患者体内呈显著性升高,是诊断原发性肝癌的重要肿瘤标志物[14]。本试验发现,OCT可以缓解LPS导致的仔猪血浆ALT、AST、ALP、GGT活力的升高,说明日粮中添加50 mg/kg的OCT可以缓解LPS对肝脏的损伤作用,从而对仔猪肝脏有保护作用,这与Wang等[15]研究结果一致。
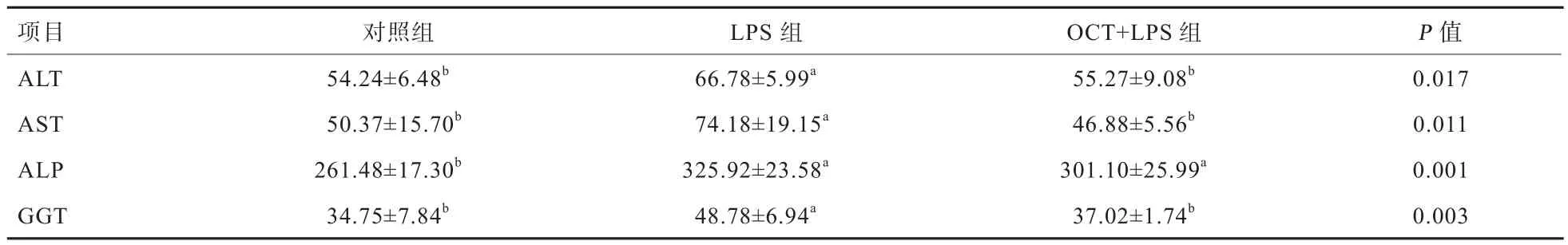
表3 OCT对LPS刺激仔猪血液生化指标的影响 U/L
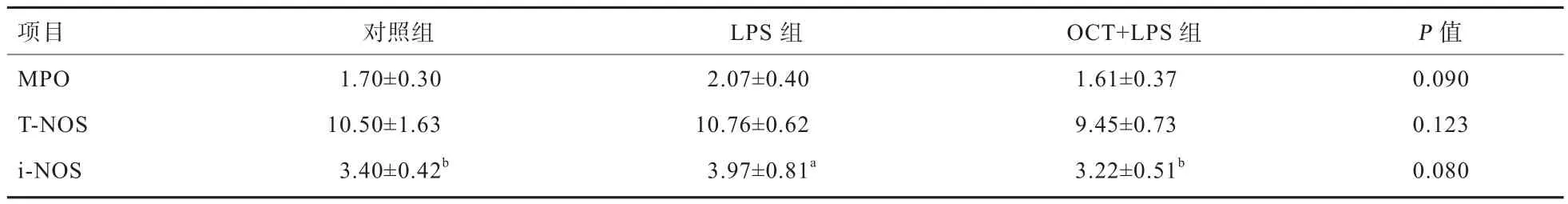
表4 OCT对LPS刺激仔猪肝脏MPO、T-NOS和i-NOS含量的影响 U/mg prot
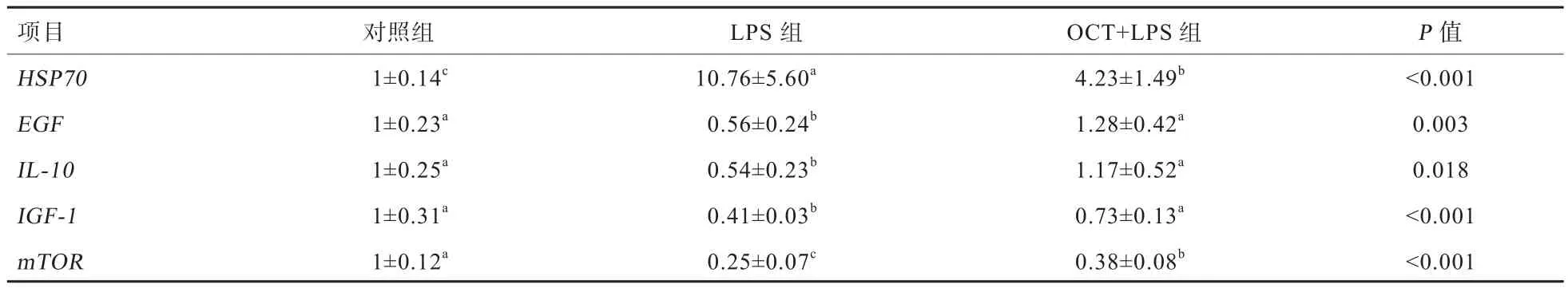
表5 OCT对LPS刺激仔猪肝脏相关基因mRNA水平的影响
MPO是一种含铁溶酶体,其活性与中性粒细胞的功能是否正常以及激活有关,当机体受到外界刺激或者疾病感染后,其含量会升高[16]。NOS分为T-NOS和i-NOS,NOS的作用主要是调控机体一氧化氮(NO)的水平。机体受到应激后,i-NOS被活化,其含量显著升高产生大量NO,高浓度的NO易结合O2-形成可诱导脂质过氧化损伤的ONOO-[17]。本试验发现,50 mg/kg的OCT可缓解LPS导致的仔猪肝脏i-NOS和MPO升高。研究表明,LPS刺激导致机体产生炎症反应,可能是LPS刺激机体中性粒细胞发挥免疫功能,同时i-NOS被活化产生大量的NO,缓解机体氧化损伤[18]。OCT可以缓解LPS刺激造成的仔猪肝脏氧化损伤,有研究表明,肉桂油有抗氧化能力,可以降低溃疡性损伤大鼠脂质过氧化水平[19];另有研究表明,肉桂油可以缓解LPS刺激导致的仔猪空肠GSH-Px的降低和过氧化氢(H2O2)的升高[15]。本研究还发现,日粮中添加OCT可以缓解LPS刺激引起的HSP70基因相对表达量的升高。HSP70通过其分子伴侣作用参与蛋白质的折叠、转运、细胞保护、抗原呈递及肿瘤免疫等许多生理活动过程;HSP70可以阻断诱导细胞凋亡的应激酶(JNK)从而减少细胞凋亡表皮生长因子[20]。HSP70也是氧化应激的标志,LPS刺激会提高仔猪肝脏HSP70的表达[21]。Wang等[15]研究表明,肉桂油可以降低LPS刺激仔猪小肠HSP70的mRNA水平。以上研究进一步说明了OCT可以通过降低LPS刺激造成的氧化应激从而发挥对肝脏的保护作用,可能是复合植物精油中的肉桂油发挥主要作用。
本研究发现,OCT缓解了LPS刺激导致的仔猪肝脏EGF、IL-10、IGF-1和mTOR基因相对表达的降低,说明日粮中添加OCT可以降低LPS刺激引起的炎症反应,提高仔猪肝脏细胞的生长和增殖。EGF主要生理活性是诱导细胞增殖、分裂分化,促进细胞生长[22]。EGF是一个庞大的族群,这个族群都是一些功能性小分子,它们通过与受体结合,打开细胞内的Ras信号通路,从而促成DNA合成并刺激细胞的增殖[23]。IL-10是抗炎因子,可抑制T细胞的活化、刺激B细胞的增殖,会阻断炎性介质NO和H2O2的产生[24]。IGF对肌肉有很强的合成代谢效应,能抑制蛋白质的分解,提高氨基酸的摄取和细胞增生,是合成代谢的直接效应物质,当机体肝脏功能受损时,IGF-1分泌显著降低。IGF对动物胚胎以及出生后的生长、发育有重要作用[25]。mTOR在感受营养信号、调节细胞增长与增殖中起关键作用,可以加快细胞的生长与增殖[26]。LPS刺激使肝脏处于一种炎症状态,炎性因子分泌增加,此外,LPS刺激后IGF-1分泌显著降低,而IGF-1位于mTOR信号通路上游,LPS刺激下调mTOR及EGF基因的表达,说明LPS也可能影响了肝细胞的增殖与分化,加快了蛋白的分解,抑制了细胞生长相关的信号通路,如mTOR和EGF等信号通路。而日粮中添加50 mg/kg的OCT缓解了LPS刺激造成的仔猪肝脏EGF、IL-10、IGF-1、mTOR基因相对表达量的降低,表明OCT对仔猪肝脏有保护作用可能与调控肝脏炎性基因的表达、mTOR和EGF信号通路有关。
4 结 论
LPS刺激使肝脏处于炎症应激状态,导致肝脏损伤,日粮中添加50 mg/kg的OCT可缓解LPS刺激导致的仔猪肝脏损伤,这可能与mTOR和EGF信号通路有关。基于此研究,未来可以对mTOR和EGF信号通路进行更深入的探究,为饲料中天然植物精油的使用提供更可靠的理论基础。
[1] 刘玉兰. 仔猪免疫应激及其营养调控研究进展[J]. 中国畜牧杂志, 2017, 53 (10): 1-3,11.
[2] Brence A, Roura E. Essential oil in poultry nutrition : Main effects and modes of action[J]. Anim Feed Sci Tech, 2010, 158(1): 1-14.
[3] 宋洪涛, 郭涛, 颜秀涛, 等. 肉桂油β-环糊精包合物的制备工艺研究[J]. 中草药, 2000, 31 (11): 818-820.
[4] 邱楚武. 牛至油在仔猪饲料中的应用试验[J]. 粮食与饲料工业, 2003 (7): 32-33.
[5] 王梓贞, 李琪. 百里香精油化学成分分析及其抗菌、抗氧化活性的研究进展[J]. 特种经济动植物, 2013, 16 (8): 35-36.
[6] 中国香料香精化妆品协会. 欧盟批准百里香酚用作植保产品[J]. 国内外香化信息, 2013(7): 518.
[7] Schmocker C, Weylandt K H, Kahlke L,et al. Omega-3 fatty acids alleviate chemically induced acute hepatitis by suppression of cytokines[J]. Hepatology, 2007, 45 (4): 864-869.
[8] 汪中兴, 侯永清, 丁斌鹰, 等. 复合植物精油对脂多糖刺激断奶仔猪生长性能和生长相关基因mRNA表达的影响[A].第七届中国饲料营养学术研讨会论文集[C].郑州: 中国畜牧兽医学会动物营养学分会, 2014.
[9] Fu W J, Stromberg A J, Viele K,et al. Statistics and bioinformatics in nutritional sciences:analysis of complex date in the era of systems biology[J]. J Nutr Biochem, 2010, 21 (7):561-572.
[10] Liu Y L, Li D F, Gong L M,et al. Effects of fish oil supplementation on the performance and the immunological,adrenal, and somatotropic responses of weaned pigs after an Escherichia coli lipopolysaccharide challenge[J]. J Anim Sci,2003, 81: 2758-2765.
[11] Collins T F X, Sprando R L, Black T N,et al. Effects of zearalenone on in utero development in rats[J]. Food Chem Toxicol, 2006, 44(9): 1455-1465.
[12] 张弛, 刘瑛, 李华珠, 等. 血清谷丙转氨酶在正常范围内对非酒精性脂肪性肝病的预测[J]. 世界华人消化杂志, 2011,19(8): 841 844.
[13] 姜晓丹, 小林俊博, 宋文光, 等. 脂多糖诱发急性肺炎时中性粒细胞碱性磷酸酶细胞化学定位观察[J]. 解剖学报,2000, 31(1): 52-55.
[14] 朱陈, 孙源源, 金燕. 血清甲胎蛋白、γ-谷氨酰转肽酶、高尔基体蛋白73检测在原发性肝癌早期诊断中的价值[J]. 临床肝胆病杂志, 2014, 31(10): 1064.
[15] Wang L, Hou Y Q, Yi D,et al. Beneficial roles of dietary oleum cinnamomi in alleviating intestinal injury[J]. Front Biosci,2015, 20(5): 814-828.
[16] 陈柏荣, 危小良, 彭湖, 等. 髓过氧化物酶与冠状动脉慢血流的相关性[J]. 中国动脉硬化杂志, 2015, 23(2): 185-187.
[17] 张伟. N-乙酰半胱氨酸对脂多糖刺激仔猪肠道屏障功能及抗氧化能力的影响[D]. 武汉: 武汉轻工大学, 2011.
[18] 张伟, 杨震国, 侯永清, 等. N-乙酰半胱氨酸对脂多糖刺激仔猪空肠黏膜抗氧化能力的影响[J]. 动物营养学报, 2011,23 (5): 842-847.
[19] Ozbayer C, Kurt H, Ozdemir Z,et al. Gastroprotective,cytoprotective and antioxidant effects of Oleum cinnamomi on ethanol induced damage[J]. Cytotechnology, 2014, 66(3): 431-441.
[20] 程维杰, 李秋玲, 孙延鸣, 等. 热休克蛋白70(HSP70)研究进展[J]. 畜牧兽医杂志, 2008, 2(6): 55-57.
[21] Yi D, Hou Y Q, Wang L,et al. Dietary N-acetylcysteine supplementation alleviates liver injury in lipopolysaccharidechallenged piglets[J]. Brit J Nutr, 2014, 111: 46-54.
[22] Hou Y Q, Wang L, Yi D,et al. N-acetylcysteine reduces inflammation in the small intestine by regulating redox, EGF and TLR4 signaling[J]. Amino Acids, 2013, 45: 513-522.
[23] 席全胜, 钱欣国, 甘人宝. EGF家族研究进展[J]. 生命化学,1999, 19(3): 135-137.
[24] 李枚涓, 王琰, 孔艳. 白介素10在粥样斑块形成机制中的研究进展[J]. 心血管进展, 2009(30): 11-16.
[25] 高萍, 傅伟龙, 朱晓丹, 等. 蓝塘仔猪IGF-1水平与组织IGF-1、GHR基因的表达[J]. 畜牧兽医学报, 2005, 36(1):38-42.
[26] 潘智, 张令强, 蒋继志, 等. mTOR的研究进展[J]. 细胞生物学杂志, 2006(28): 395-398.
