G0S2基因在食管癌组织中表达变化及对癌细胞增殖、凋亡的影响
2018-03-19祝铖李莲唐俊明张蕾郑飞
祝铖,李莲,唐俊明,张蕾,郑飞
(湖北医药学院附属人民医院,湖北十堰442000)
G0S2基因最早发现于血液单核细胞[1]。早期研究认为,G0S2基因在调节细胞周期及细胞增殖方面有重要作用[2]。研究发现,G0S2基因能够与其他蛋白相结合来发挥自身生物学效应,如通过结合B淋巴细胞瘤-2基因促进肿瘤细胞凋亡[3]。G0S2基因可能具有抗癌作用,但尚未得到验证。食管癌组织中存在G0S2基因甲基化现象,其因甲基化而沉默[4],提示G0S2基因消除甲基化恢复表达可能成为治疗食管癌的新途径。本研究观察食管癌组织中G0S2基因的表达情况,并观察G0S2基因高表达对食管癌细胞增殖和凋亡的影响,为防控食管癌提供新的方向。
1 材料与方法
1.1 材料来源 2016年3月~2017年4月本院手术切除的45例份食管癌组织(患者男40例、女5例,年龄47~70岁、中位年龄59岁,术前均未行放化疗)及其相应的癌旁正常组织(距癌组织边缘>5 cm),均经病理检查证实,于液氮中冻存。TRIzol试剂、DEPC水购于美国Sigma公司;RNA逆转录试剂盒购于美国赛默飞公司;RT-PCR上、下游引物及GAPDH引物均购于武汉金开瑞生物工程公司;琼脂糖粉购于Lonza公司;食管癌KYSE-70细胞购于中国科学院上海细胞所;G0S2腺病毒裂解液与空载体由本院临床医学研究所提供;甲基化抑制剂5-氮杂-2-脱氧胞苷试剂购于Sigma公司;RPMI-1640培养基、胎牛血清、胰酶等均购于美国Gibco公司。
1.2 食管癌及癌旁正常组织中G0S2基因表达检测 采用qRT-PCR法。按照TRIzol试剂说明书提取组织RNA,根据RNA逆转录试剂盒说明书对已提取的RNA进行逆转录。G0S2行qRT-PCR扩增,上游引物:5′GGCCTGATGGAGACTGTGTG3′、下游引物:5′CTTGCTTCTGGAGAGCCTGT3′,全长115 bp。GAPDH上游引物:5′GAAGGTGAAGGTCGGAGTC3′,下游引物:5′GAAGATGGTGATGGGATTTC3′。qRT-PCR反应体系10 μL,反应条件:95 ℃预变性10 min,95 ℃变性15 s,60 ℃退火延伸32 s,共40个循环。读取各样本Ct值,用2-ΔΔCt法计算基因相对表达量。
1.3 KYSE-70细胞培养及处理 用含10%胎牛血清的RPM 1640培养基培养KYSE-70细胞,置于37 ℃、5% CO2、湿度适宜的培养箱中。待细胞长至70%~80%时,随机分为空白组、Ad-Null组、Ad-G0S2组及0、25、50、100 μmol/L 5-氮杂-2-脱氧胞苷组。Ad-Null组、Ad-G0S2组分别转染Ad-G0S2、Ad-Null,0、25、50、100 μmol/L 5-氮杂-2-脱氧胞苷组分别给予0、25、50、100 μmol/L浓度的甲基化抑制剂5-氮杂-2-脱氧胞苷进行处理。将各组细胞继续培养24 h。用 qRT-PCR技术检测各组G0S2mRNA表达,方法同1.2。转染后及经5-氮杂-2-脱氧胞苷处理后细胞内G0S2mRNA相对表达量升高,说明实验成功。
1.4 细胞增殖能力检测 采用CCK-8法。收集各组细胞接种于96孔板,每孔1.0×104/mL,每组设置5个复孔。培养结束后向每孔滴加CCK-8试剂10 μL,孵育2 h。用酶标仪测定450 nm波长下各个孔的吸光度值。
1.5 细胞凋亡率检测 采用Annexin V-FITC/PI双染法。取各组细胞用4 ℃预冷无菌PBS洗涤2次,并用0.25%的胰酶(含0.02%EDTA)消化离心,用Binding Buffer 200 μL重悬细胞,调整细胞为(2~5)×105/mL,取细胞悬液195 μL,加入Annexin V-FITC 5 μL,轻轻混匀后间隔3 min再加入PI 10 μL,混匀,室温避光孵育10 min。在反应管中加入无菌PBS 300 μL,混匀。上机检测细胞凋亡情况,计算细胞凋亡率。
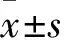
2 结果
2.1 G0S2基因在食管癌组织和癌旁组织中的表达比较 G0S2基因在食管癌组织和癌旁组织中的相对表达量分别为31.9±7.8、1,二者比较差异有统计学意义(P<0.05)。
2.2 G0S2对KYSE-70细胞增殖的影响 空白组、Ad-Null组、Ad-G0S2组及0、25、50、100 μmol/L 5-氮杂-2-脱氧胞苷组吸光度值分别为0.99±0.05、0.91±0.12、0.56±0.17、0.87±0.08、0.75±0.03、0.68±0.06、0.60±0.02。Ad-G0S2组吸光度较其他两组低(P均<0.05),空白组与Ad-Null组吸光度值比较差异无统计学意义(P>0.05);随5-氮杂-2-脱氧胞苷浓度升高,吸光度值逐渐降低,细胞增殖活力下降(P均<0.05)。
2.3 G0S2对食管癌KYSE-70细胞凋亡的影响 空白组、Ad-Null组、Ad-G0S2组及0、25、50、100 μmol/L 5-氮杂-2-脱氧胞苷组细胞凋亡率分别为3.2%±2.8%、3.6%±2.1%、12.1%±1.8%、1.3%±0.4%、4.1%±1.7%、7.3%±1.9%、9.5%±1.1%。Ad-G0S2组细胞凋亡率较其他两组高(P<0.01),空白组与Ad-Null组凋亡率比较差异无统计学意义(P>0.05);随5-氮杂-2-脱氧胞苷浓度升高,细胞凋亡率升高(P<0.05)。
3 讨论
研究表明,G0S2基因能调节细胞增殖分化,促进癌细胞凋亡,抑制肿瘤发生发展等;调节细胞由G0期向G1期转化,细胞进入G1期后诱导G0S2基因瞬时表达,G0S2反过来抑制细胞由G0期进入G1期,从而能使细胞稳定增殖和分化[5]。细胞只有正常地表达G0S2基因,才能够正常、稳定地进入细胞分化的过程[6]。最近有研究指出,接受造血干细胞移植的患者,其外周血单核细胞中的G0S2基因表达明显上调[7]。研究发现,胶质瘤患者中癌细胞G0S2基因低表达,与胶质瘤细胞的发生发展密切相关[8]。以上说明G0S2基因对细胞增殖有重要的稳定和调控作用。抑癌基因失活是癌症发生的重要因素之一[9],而抑癌基因的启动子区发生甲基化是其失活的重要环节[10]。癌组织与正常组织中的G0S2基因甲基化程度存在明显差异,因此G0S2基因的表达水平亦存在明显差异[11]。近年来研究发现,G0S2基因的启动子区因发生高甲基化而沉默,呈低表达,从而导致癌症的发生,在头颈部癌、肺癌、乳腺癌等多种癌症中均已被发现并证实[12]。最近在皮肤鳞癌组织中也发现G0S2基因因甲基化而沉默的现象[13]。研究发现,人慢性髓系白血病细胞的G0S2基因调控序列和外显子区因处于高甲基化的状态而沉默,使用特定的去甲基化试剂后发现,细胞增殖活性降低[14];相反利用特定技术使G0S2基因沉默,则促进癌细胞增殖。有研究用shRNA使大肠癌细胞的G0S2基因沉默,发现大肠癌细胞增殖活力明显提高[15]。由此可见,一方面甲基化现象在癌症中广泛存在,如何消除甲基化使G0S2基因正常、稳定地表达从而发挥抑癌作用,是癌症高效治疗的关键所在。另一方面说明G0S2基因高表达治疗食管癌具有一定的理论依据;且食管癌的传统治疗方案效果欠佳,食管癌患者5年生存率较低,生活质量差。目前对G0S2基因在食管癌治疗中作用的研究较少,以此为研究方向可能成为食管癌高效治疗的突破口。本研究结果显示,食管癌组织中G0S2基因呈低表达,证明食管癌的发生可能与G0S2基因低表达有密切关系;通过转染及甲基化转移酶抑制剂5-氮杂-2-脱氧胞苷使G0S2基因过表达后两种方法,G0S2基因能够抑制食管癌KYSE-70细胞的增殖,促进其凋亡。
G0S2基因正常表达是抑制食管癌细胞增殖、促进其凋亡的重要因素;如何消除甲基化从而使G0S2基因正常、稳定地表达可能是预防和治疗食管癌的研究热点。G0S2基因的甲基化修饰及其在食管癌发生、发展中如何发挥作用有待进一步的研究。
[1] Siderovski DP, Blum S, Forsdyke RE, et al. A set of human putative lymphocyte G0/G1 switch genes includes genes homologous to rodent cytokine and zinc finger protein-encoding genes[J]. DNA Cell Biol, 1990,9(8):579.
[2] Russell L, Forsdyke DR. A human putative lymphocyte G0/G1 switch gene containing a CpG-rich island encodes a small basic protein with the potential to be phosphorylated[J]. DNA Cell Biol, 1991,10(8):581-591.
[3] Oh SA, Suh Y, Pang MG, et al. Cloning of avian G0/G1 switch gene 2 genes and developmental and nutritional regulation of G0/G1 switch gene 2 in chicken adipose tissue[J]. J Anim Sci, 2011,89(2):367-375.
[4] Wan SG, Doong TJ, Chen H, et al. Zinc deficiency and G0S2DNA methylation in rat esophageal carcinogenesis[J]. Cancer Res, 2010,70(8s):146.
[5] Heckmann BL, Zhang X, Xie X, et al. The G0/G1 switch gene 2 (G0S2): regulating metabolism and beyond[J]. Mol Cell Biol L, 2013,1831(2):276-281.
[6] Choi H, Lee H, Kim TH, et al. G0/G1 switch gene 2 has a critical role in adipocyte differentiation[J]. Cell Death Differ, 2014,21(7):1071-1080.
[7] Verner J, Kabathova J, Tomancova A, et al. Gene expression profiling of acute graft-vs-host disease after hematopoietic stem cell transplantation[J]. Exp Hematol, 2012,40(11):899.
[8] Yim CY, Sekula DJ, Freemantle SJ, et al. G0S2functions as a tumor suppressor gene through inhibition of c-Myc[J]. Cancer Res, 2015,75(15 s):2026.
[9] Barreau O, Assié G, Wilmot-Roussel H, et al. Identification of a CpG island methylator phenotype in adrenocortical carcinomas[J]. J Clin Endocr Metab, 2013,98(1):174-184.
[10] Filiz O, Julide A, Ates K, et al. Methylation of tumor suppressor genes in ovarian cancer[J]. Exp Ther Med, 2012,4(6):1092-1096.
[11] Huang WY, Hsu SD, Huang HY, et al. Meth HC: a database of DNA methylation and gene expression in human cancer[J]. Nucleic Acids Res, 2015,43:D856-D861.
[12] Kusakabe M, Watanabe K, Emoto N, et al. Impact of DNA demethylation of the G0S2gene on the transcription of G0S2in squamous lung cancer cell lines with or without nuclear receptor agonists[J]. Biochem Bioph Res Co, 2009,390(4):1283-1287.
[13] Nobeyama Y, Watanabe Y, Nakagawa H. Silencing of G0/G1 switch gene 2 in cutaneous squamous cell carcinoma[J]. PLoS One, 2017,12(10):e0187047.
[14] Yamada T, Park CS, Ye S, et al. G0S2inhibits the proliferation of K562 cells by interacting with nucleolin in the cytosol[J]. Leukemia Res, 2014,38(2):210-217.
[15] Rachid Z, Wissal EA, Isabelle G, et al. Inhibition of adipose triglyceride lipase (ATGL) by the putative tumor suppressor G0S2or a small molecule inhibitor attenuates the growth of cancer cells[J]. Oncotarget, 2015,6(29):28282-28295.
