基于生物光谱技术的污染物毒性效应研究进展
2018-03-16徐笠蔚青李君逸韩丽花殷敬伟陆安祥
徐笠,蔚青,3,李君逸,韩丽花,3,殷敬伟,陆安祥,*
1. 北京农业质量标准与检测技术研究中心,北京市农林科学院,北京100097 2. 农产品产地环境监测北京市重点实验室,北京 100097 3. 中北大学理学院化学系,太原 030051 4. 清华大学摩擦学国家重点实验室,北京 100084
应用生物光谱技术研究污染物毒性效应是毒理学研究中的一个新兴领域。最常用的生物光谱技术包括红外光谱技术(Infrared spectrometry)和拉曼光谱技术(Raman Spectroscopy)[1-2]。红外光谱技术进行生物样品的无损分析是一个快速发展的研究领域,而且生物大分子在构型和构象上的变化可以用来表征环境污染物对生物细胞、组织或者器官上的毒性效应,因此运用红外光谱结合多元统计的手段,不仅可以灵敏、有效地评价污染物的毒性效应,还可以快速、准确地推断其毒性作用机制[3-4]。拉曼光谱学是一种强大的、非侵入性的、高通量的方法,它被用来在样本中产生化学实体的光谱代表,并且能够检测细胞的变化[5-6]。拉曼光谱是散射光谱,而红外光谱则是吸收光谱,但两者均是研究分子振动的重要手段。红外和拉曼光谱在污染物毒性效应研究中具有灵敏度较高、检测时间短、仪器操作简单、所得生物信息丰富、光谱信息特征性较强和样品前处理简单且不造成二次污染等优势,并且应用范围非常广泛,在细胞、组织和微生物等方面都可以应用,且呈现出其独特的优势[7-11]。本文简要介绍了这2种技术的原理以及数据处理流程,重点介绍了这2种光谱技术在污染物毒性效应研究中的进展情况,并对未来主要的研究方向进行了展望。
1 生物光谱法概述(Overview of biospectroscopy)
生物光谱学是光谱技术在生物检测分析中的应用,该技术不仅可以获得样品形态学上的结果,而且可以得到元素或分子层面的信息,成为一个新的领域学科。生物光谱法包括红外光谱法、拉曼光谱法、荧光光谱法和X射线光谱法。目前,国内外研究学者主要是利用红外光谱法和拉曼光谱法2种方法在污染物毒性效应方面开展了一系列研究。
1.1 红外光谱
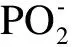
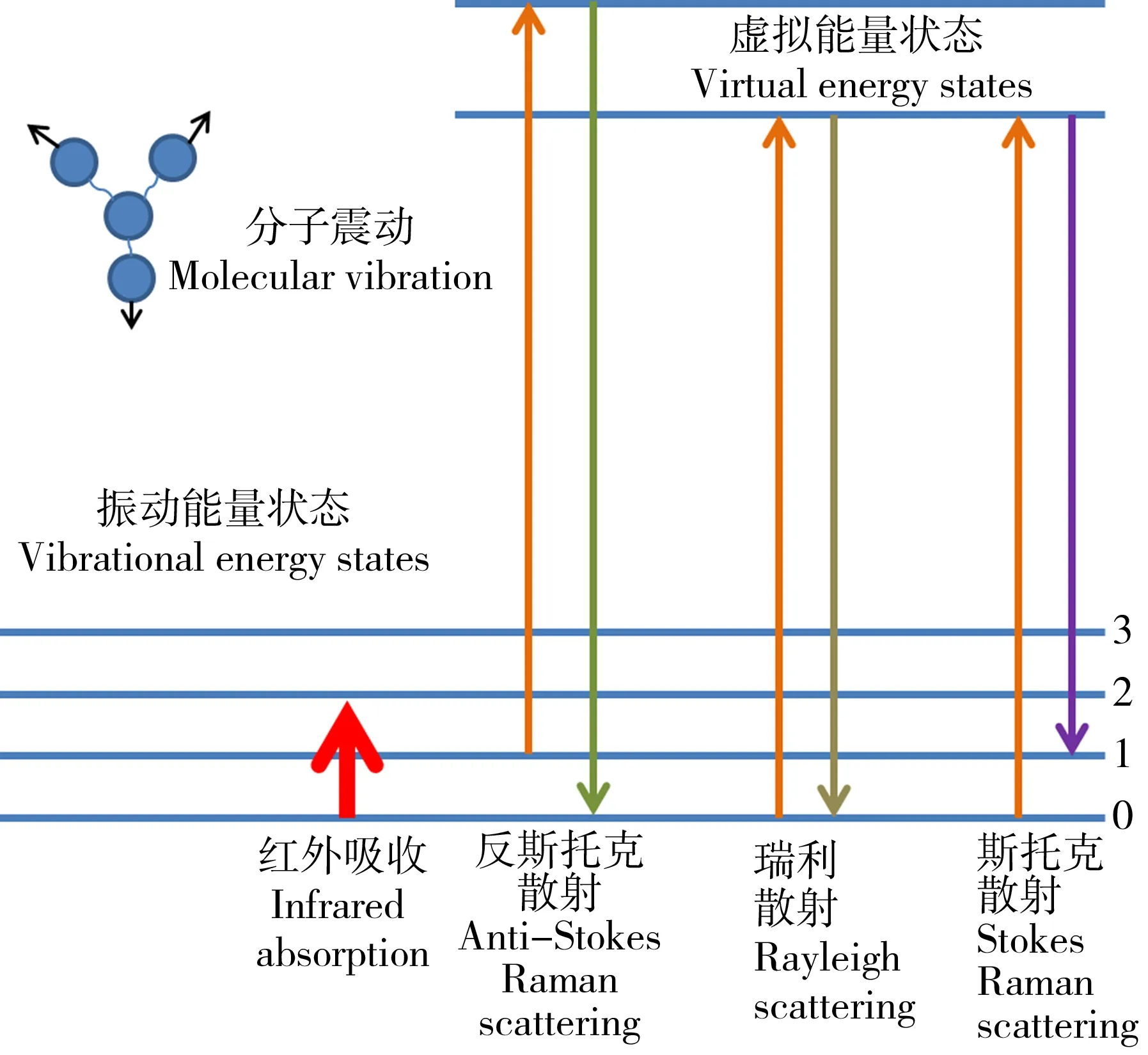
图1 红外吸收和拉曼散射分子振动原理Fig. 1 Molecular vibration principle of infrared absorption and Raman scattering
1.2 拉曼光谱
拉曼是一种单色光源的光子被吸收并释放回来
后,分子的振动能量也随之增加和减少的散射技术。通常情况下,有时分子不会返回到它原来的能量状态,为了保持能量平衡,在释放的光子中会发生频率偏移。这种位移现象称为非弹性散射或拉曼散射(图1)[26]。散射分为瑞利散射与拉曼散射2种,拉曼散射包括斯托克拉曼散射(散射光频率小于入射光频率)和反斯托克拉曼散射(散射光频率大于入射光频率),斯托克的强度远大于反斯托克的强度,在样品分子的拉曼光谱测试中都是以斯托克散射峰为检测峰,忽略反斯托克散射的信号[27]。拉曼强度、拉曼频移以及线宽都可以反映出拉曼样品池中样品物质的结构特征,与物质分子的化学键、官能团的振动和转动有关。拉曼光谱可分为普通拉曼光谱和表面增强拉曼光谱,在使用普通拉曼光谱时,经常会遇到强烈的荧光背景(固有的或杂质引起的),这会降低光谱的质量,并对信噪比(S/N)产生不利影响[28]。表面增强拉曼光谱(SERS)由于其增强拉曼横截面的性质,可以提供强烈的光谱,并帮助我们克服普通拉曼的这些缺点[29]。为生物研究中的应用提供了一种快速、高灵敏度的化学结构信息检测工具[30-32]。通常,在生物样本中观察到的拉曼光谱的重要区域在400~2 000 cm-1波数内,与蛋白质(1 500~1 700 cm-1)、碳水化合物(470~1 200 cm-1)、磷酸基团DNA(980、1 080和1 240 cm-1)和其他细胞生物分子的键合振动相关。在更高的波数(2 700~3 500 cm-1)中也可以观察到与脂质和蛋白质中的CH、NH和OH对应的更高频率的伸缩振动峰[33]。
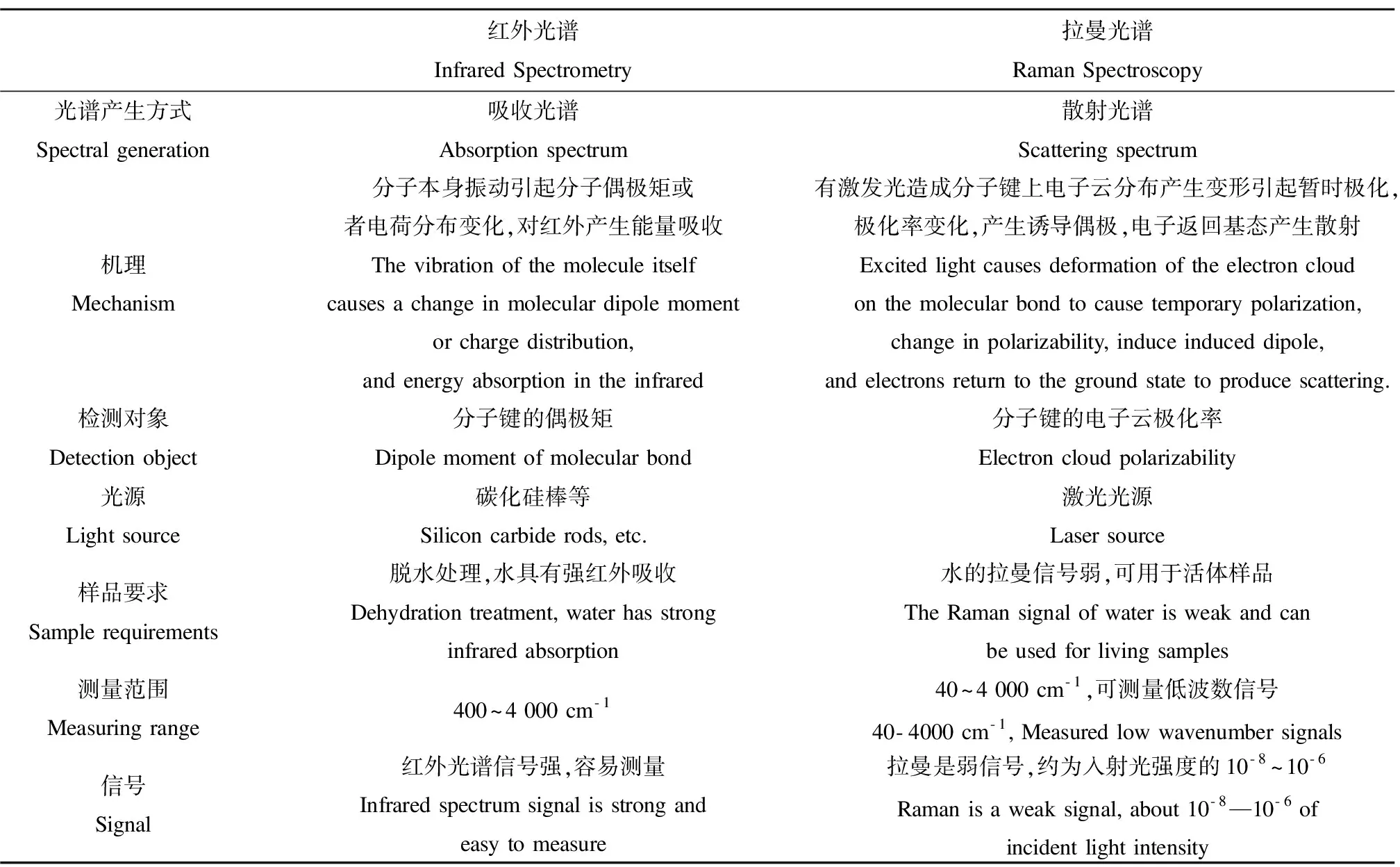
表1 红外和拉曼光谱的原理、特点及差异Table 1 Principle, characteristic and difference of Infrared Spectroscopy and Raman Spectroscopy
拉曼光谱和红外光谱类似,同样可以对物质的结构和性质进行定量、定性分析,不同的拉曼峰对应了不同的官能团,与红外相比拉曼的一个突出优点是:水分对生物指纹区的拉曼信号没有影响,因此拉曼光谱还可以用于活体成像。此外,受限于红外光的波长衍射极限,红外光谱显微镜的空间分辨率受限,一般都是微米级别以上的,而拉曼光谱可以使用短波长的激光器做激发,并结合共聚焦显微系统,可以达到百纳米级别的空间分辨率。红外和拉曼光谱的特点及差异详见表1。
2 生物光谱数据处理方法(Biospective data processing method)
通常生物光谱实验采集复杂生物化学信息数据集的成百上千个差异细微的图谱,而且由于生物样本本身的复杂性使得图谱中存在着很多的谱峰重叠性,因此需利用化学计量学方法对图谱进行分析。光谱数据分析大致分为3个部分:光谱数据预处理、提取光谱信息特征、以及信息分类和光谱特征峰解析(图2),可以利用一些软件对数据进行处理(表2)。
2.1 光谱数据预处理
预处理旨在提高后续多变量分析的稳健性和准确性,并通过纠正与光谱数据采集相关的问题来提高数据的可解释性。光谱数据前处理包括剔除异常值、剪切、降噪、基线校正以及归一化[34]。在某些情况下,光谱的目视检查显示出明显的异常值,包括大量的光谱污染、荧光或非常差的S/N,可以要剔除这些明显的异常值。样品制备、背景荧光以及电荷耦合器件的热波动会产生噪声和影响光谱基线。因此,需要降低噪声和基线校正以提高光谱质量,降噪和基线校正的第一种方法就是改变样品制备方法以及光谱仪器的采集设置,其次还可以通过一些相应的方法进行处理。比如常用的基线校正、降噪处理方法包括直接基线校正(rubber-band correction)、多项式拟合基线校正(polynomial-correction)、一阶微分(1st order differentiation)、小波降噪(wavelet-deniose),其中直接基线校正常用于谱图背景相对简单的情形中,多项式拟合主要针对于复杂背景条件下的基线校正,常用于拉曼谱图的校正,但是存在一定误差,一阶微分的方法则适用于复杂谱图背景条件下,例如像包含荧光背景的拉曼谱图,但是也会引入过多的噪音,而小波降噪则正好适用于含大量噪音信号成分的谱图中,进行降噪处理[35]。归一化红外和拉曼光谱至关重要,以解决混淆因素,可以运用酰胺I、酰胺II、矢量归一化、最大-最小值归一化、标准化和中心化等方法对光谱数据进行归一化[36]。
2.2 提取光谱信息特征
特征提取大致分为特征构造和特征选择2种方法。特征构造是在数据中创造新特征,从而推断出其他不太清楚的信息[37]。这对于其他同质数据集中的诊断,生物标记提取和模式识别尤为重要,并且它在高光谱成像中具有重要作用,因为单个像素可以减少与光谱强度或方差有关的单个值[38]。特征选择是从数据集推断现有特征,例如特定波数,可用于确定光谱生物标志物和提供诊断框架。光谱信息的特征提取包括分类、回归、聚类和降维等相关的处理方法,常用的提取方法包括主成分分析(PCA)[39]、偏最小二乘回归(PLS)[40]、线性判别分析(LDA)[41]、支持向量机分析(SVM)[42-43]及主成分结合线性判别分析(PCA-LDA)[1]。
2.3 信息分类和光谱特征峰解析
在成像和诊断研究中,通常需要基于光谱对样品进行分类,因为光谱可以基于先前的用户输入(监督分类)或单独的光谱方差(无监督)进行分类。无监督分类通常依赖于聚类技术,其中层次聚类分析,k均值聚类和模糊C均值聚类是3种常用的方法[44],监督分类常见的方法包括线性判别分类器(LDCs)、人工神经网络和支持向量机(SVMs)等[45-46]。生物样本主要组成成分包括脂质、蛋白质、核酸和糖类等。所以解析光谱特征峰主要包括:分析脂质特征吸收峰(烷基链结构(CH2和CH3)、羰基及磷酸基团的振动信息),得出与细胞膜脂质结构及膜脂过氧化程度的信息;分析蛋白质分子相关的振动信息主要包括酰胺I、酰胺II、游离氨基酸残基及磷酸化蛋白相关的振动信息;分析酰胺I的结构可以得到蛋白质二级结构信息,分析游离氨基酸可以得到细胞内氨基酸侧链信息及蛋白质翻译后修饰的信息等;分析与核酸振动信息相关的吸收峰可以得到与DNA二级结构的变化情况。
3 生物光谱法应用研究进展(Progress in the application of biospectroscopy)
3.1 红外光谱在污染物毒性效应研究进展
3.1.1 细胞毒性效应研究进展


表2 红外和拉曼光谱数据分析软件Table 2 Data analysis software for Infrared spectroscopy and Raman spectrum
上述研究大都侧重于单一污染物在同一细胞周期的毒性效应,也有部分研究开展了同一污染物对处于不同周期细胞的毒性效应以及复合污染的细胞毒性效应,如Li等[51]运用ATR-FTIR研究了不同类型碳纳米材料(C60、长多壁碳纳米管、短多壁碳纳米管)的细胞毒性效应,结果表明:G0/G1期细胞的抗氧化酶的活性要高于S期,G0/G1期细胞对这3种碳纳米材料的毒性效应更加敏感。Li等[4]将鱼鳃细胞和MCF-7共同暴露在C60和苯并芘环境下,然后用显微傅立叶变换红外光谱法(Micro-FTIR)分析这2种细胞的生物结构信息。结果显示低浓度C60可以促进苯并芘对细胞的氧化损伤,高浓度C60却减弱这种影响效果;尽管低浓度C60可以促进苯并芘氧化损伤水平,但是若这2种物质共同处理细胞,却减弱了任一种污染物单独诱导的细胞氧化损伤水平。Llabjani等[52]运用ATR-FTIR研究多氯联苯和多溴二苯醚对MCF-7的复合毒性作用时发现,通过相似调节机制作用的不同污染物可能产生抑制作用,而具有独立作用机制的污染物可能增强该毒性作用。
3.1.2 植物毒性效应研究进展
红外光谱在植物毒性效应方面的研究主要集中在浮萍和藻2种植物上。Hu等[14]使用FTIR结合多元统计的方法监测4种化学物质(Cu、Cd、阿特拉津和乙草胺)和4种金属工业废水对浮萍的蛋白质、脂质、DNA/RNA和碳水化合物等生物化学分子变化的影响情况,从而鉴定其对浮萍的毒性效应,结果表明以光谱信息为工具的毒性评价方法不但准确、可靠,而且能够提供多效应终点的评价结果。Xin等[53]利用SR-FTIR研究了三氯生和卡马西平对绿藻的毒性机制,结果显示三氯生对绿藻的毒性要强于卡马西平。三氯生通过抑制脂肪酸合成进而影响蛋白质合成,这种毒性作用在高浓度(100 μmol·L-1)下是不可逆转的,但是随着时间的延长它的浓度会减弱到0.391 μmol·L-1;卡马西平可以产生疏水性的相互作用,影响磷脂双层结构,并对特定的蛋白质起作用,使细胞膜失去功能。但是在延长暴露时间的同时,暴露在卡马西平下的绿藻细胞会产生耐药性。Dao等[54]利用ATR-FTIR研究了Pb对2种绿藻(小球藻和栅藻)的毒性效应,结果表明:在Pb暴露条件下,绿藻的多糖和脂质含量增加,但是蛋白质和磷酸分子含量减少,红外光谱所表现的结果与光合参数与常规脂质测定的结果相吻合。
3.1.3 动物毒性效应研究进展
Li等[55]将斑马鱼分别暴露于C60、长多壁碳纳米管、短多壁碳纳米管和单壁碳纳米管21 d,ATR-FTIR结果显示:高剂量的碳纳米材料(CNPs)对斑马鱼的鳃和大脑的毒性更大,而低剂量的CNPs对性腺和肝脏的毒性更强;CNPs对斑马鱼的脂质、蛋白质和DNA/RNA产生显著变化;脑和性腺对CNPs的响应更敏感,与对照组相比,长多壁碳纳米管对大脑中脂质与蛋白质的比值没有明显的影响,而C60、短多壁碳纳米管和单壁碳纳米管会导致脂质与蛋白质的比率显著升高,CNPs对雄性大脑的DNA/RNA光谱区域的影响比雌性更大。CNPs对大脑的干扰可能扰乱下丘脑-垂体-性腺轴,并进一步影响性腺;所以CNPs对斑马鱼具有一定的神经毒性和生殖毒性。Anusha等[56]将老鼠每天吸入蚊香8 h并持续30 d,结果显示蚊香对老鼠的肝、肾、肺、心、脑等多种组织和功能有损伤情况。与对照组相比,吸入蚊香的老鼠肝脏、肾脏、肺和大脑中的生物大分子都发生了变化,主要表现在蛋白质和脂质相关振动峰降低。生物大分子改变程度在老鼠的肝脏和肾脏最显著,接下来是肺和大脑。Palaniappan和Pramod[57]利用FTIR研究了二氧化钛纳米粒子及其块状材料对斑马鱼大脑生化成分的影响,研究结果显示:纳米二氧化钛的毒性要强于其他块状材料;与对照组相比,暴露在二氧化钛下的斑马鱼,其大脑组织中蛋白质、脂质和核酸等生化成分都发生了改变。大脑组织中脂质被氧化,导致羰基化物增加,因此羰基与-CH2的峰积分面积比升高,进一步证明大脑生化成分的改变主要是因为活性氧的过量产生。Prabha等[58]从印度泰米尔纳德北部的池塘收集了南亚野鲮,然后以不同含量的黑藻为鱼饲料,研究这种淡水鱼的铅毒性试验。结果发现,对比铅中毒和不同浓度黑藻处理过的鱼骨样品,鱼骨中的蛋白质、碳水化合物和脂质的频带面积和峰强都有显著变化。当南亚野鲮喂含20%黑藻的饲料,其鱼骨的铅含量显著降低,说明黑藻具有对南亚野鲮骨组织中铅诱导毒性的抗骨质疏松作用。
3.1.4 微生物毒性效应研究进展
Riding等[59]运用FTIR研究了碳纳米材料对革兰氏阴性菌的毒性效应,研究发现C60、长多壁碳纳米管、短多壁碳纳米管和单壁碳纳米管都导致细菌的脂质、酰胺II的DNA改变,改变程度随纳米粒子大小而不同,短多壁碳纳米管比长多壁碳纳米管毒性更强,C60毒性最小。相比于其他,而C60所表现出的光谱信息差异最显著,且主要表现在蛋白质方面。Gupta等[60]利用红外光谱法研究了重金属Ni和Cr对大肠杆菌的毒性的相互作用,结果表明Ni(II)对大肠杆菌的亲和性要强于Cr(VI),因为它可能会侵入到大肠杆菌蛋白质的S层,所以它对大肠杆菌的毒性也要强于Cr。并且随着Ni的不断加入,会导致Cr对大肠杆菌的毒性增强,二者表现为协同作用。Ni会使大肠杆菌的脂质含量降低,在3 000~2 800 cm-1和~1 455 cm-1的频带峰面积减小,所以比较脂质谱带面积可以去监测金属毒性而导致的脂肪酸变化。Heys等[61]使用ATR-FTIR和SR-FTIR光谱研究碳纳米材料(CBNs)对革兰氏阴性和革兰氏阳性细菌的影响。结果显示革兰氏阳性和革兰氏阴性菌在接触CBNs时表现出不同的变化。革兰氏阳性细菌对这些材料表现出更强的抵抗力,这可能是由于它有更坚固的细胞壁来维持其完整性,但是在接触CBNs后革兰氏阴性和革兰氏阳性细菌的酰胺II都发生了变化。SR-FTIR因其高分辨率能够提供反映纳米毒性变化的详细的、深入的信息,而ATR-FTIR光谱学则提供了被CBNs诱导所导致的细菌改变细节的合理概述。
上述研究已表明,红外光谱技术(FTIR、ATR-FTIR和micro-FTIR)可以广泛用于不同污染物对不同生物的毒性效应研究之中,这些技术有着各自的优缺点,如ATR-FTIR在这3种测定方法中是最灵敏的,但是该方法测定会破坏样品,而且比较耗时;micro-FTIR灵敏度没有ATR-FTIR高,但是可以不破坏样品,并且可以进行面扫描。因此,在运用这些红外技术研究污染物毒性效应时,需根据研究目的和样品选择合适的方法。
3.2 拉曼光谱在污染物毒性效应的研究进展
拉曼光谱技术是以拉曼散射效应为基础,以光子为探针,具有实时在线分析、无损快速检测等优秀特点的分子结构表征技术。近年来,拉曼光谱可以提供生物分子指纹图谱,如蛋白质、DNA、脂类以及氨基酸、嘌呤等代谢中间产物的信息,因此能够从分子水平上研究污染物的毒性效应。
3.2.1 常规拉曼光谱
Li等[62]利用拉曼光谱研究了C60、长多壁碳纳米管、短多壁碳纳米管3种碳纳米材料对MCF-7的毒性效应,结果表明:这3种纳米材料均造成了胱氨酸和蛋白质比值的升高,其中短多壁碳纳米管暴露造成的胱氨酸和蛋白质比值最高,胱氨酸和蛋白质比值增加与活性氧含量增加是成比例的,因此,也说明了短多壁碳纳米管造成了活性氧含量升高的幅度最大。林凡佳等[63]使用拉曼光谱分析不同浓度的凋亡诱导剂(黄连素)对人肝癌细胞(HepG2)内吞Si纳米粒子在520 cm-1的特征拉曼峰的平均信号强度,表征细胞内吞Si纳米粒子的能力。结果表明,随着黄连素浓度的增加,细胞对Si纳米粒子的内吞能力减弱,原因是HepG2细胞凋亡过程中线粒体的活性不断减弱,导致能量代谢受阻。Neugebauer等[64]采用拉曼光谱发现抗生素环丙沙星导致短小芽胞杆菌蛋白质峰减弱和核酸峰增强,从而认为环丙沙星能够结合rRNA,进而抑制蛋白质的合成和核酸含量的增加。Li等[55]将斑马鱼分别与C60、长多壁碳纳米管、短多壁碳纳米管和单壁碳纳米管接触21 d,然后用拉曼光谱分析了斑马鱼的大脑和性腺中的不饱和脂质含量(C=C和CH2的比例),结果显示经过处理的斑马鱼的脂质与对照组的斑马鱼差异很明显,其中雄性斑马鱼中不饱和脂质含量要高于雌性。赵晶[65]利用拉曼光谱研究了4-壬基酚对大鼠睾丸组织的影响,结果表明:4-壬基酚暴露组与对照组之间有显著性差异(P<0.001),主要表现在脂质与蛋白质比率和不饱和脂肪水平等方面。
3.2.2 表面增强拉曼光谱
与常规拉曼相比,SERS可以使接近或吸附于金、铝和银等粗糙金属表面或者纳米结构(如等离子体磁性二氧化硅纳米管[66])的分子的拉曼信号增强,其信号比常规拉曼散射增强106~1014倍[67]。SERS可以极大地提高拉曼信号强度、缩短采谱时间和淬灭荧光干扰,有效地解决常规拉曼所遇到的问题。Cui等[68]利用SERS研究了Ag对大肠杆菌的毒性,结果表明在Ag纳米粒子的作用下大肠杆菌的蛋白质、次黄嘌呤、腺嘌呤和鸟嘌呤核苷拉曼光谱带会发生变化,其主要原因是Ag纳米粒子影响嘌呤的代谢和蛋白质的合成过程,并且粒径小的Ag纳米粒子要比大的毒性更大。Zhang等[69]借助无纳米毒性和高SERS活性纳米金方法,研究了不同浓度纳米ZnO(0、0.6、3、15、60、240、480 mg·L-1)作用于大肠杆菌不同时间(0.5、1.5、3、6、12、24 h)后细菌的特征生物变化和毒性动力学过程。结果表明,纳米ZnO的毒性机制在于产生活性氧基团,导致损伤核酸和改变了膜的通透性,而细菌通过合成相关的防御蛋白来降低这种不利影响。纳米Zn在0.5 h即可发挥毒性作用,且低剂量长期暴露能够发挥与高剂量暴露相同的毒性作用。Walter等[5]通过使用SERS研究金属纳米粒子对链霉菌的毒性效应,结果显示随着Ni2+浓度的增加链霉菌中蛋白质(1 660和1 448 cm-1)的峰的相对强度增加,而DNA(1 574和1 480 cm-1)的峰强在减弱。El-Zahry等[70]研究了Ag纳米粒子对细菌的毒性效应,再次证明Ag纳米粒子的抗菌机制是影响嘌呤的代谢进而抑制蛋白质的合成。并且通过SERS谱图分析发现,Ag纳米粒子的粒径和浓度对细菌的抗菌性能影响最大。Cui等[71]利用拉曼光谱研究重金属As(V)和四环素对细菌的毒性效应,发现As(V)和四环素之间是交互抗性,As(V)在增强细菌耐药性的同时阻止四环素起作用。Lu等[72]借助SERS和FTIR能够预测细胞损伤类型和程度,然后深入了解大蒜萃取物和二烯丙基硫化物的抗菌机理。Li等[62]利用SERS分析了0.1 mg·L-1、0.01 mg·L-1和0.001 mg·L-1的C60、长多壁碳纳米管、短多壁碳纳米管和单壁碳纳米管对A549肺细胞的毒性效应,研究发现这些碳基纳米离子会导致A549肺细胞的DNA甲基化水平增强,这也是首次发现纳米粒子会改变细胞的甲基化水平。
拉曼光谱主要是针对非极性分子的检测,而红外光谱主要是针对极性分子的检测,因此2种之间是互补的关系。此外,和红外光谱相比,拉曼光谱在污染物毒性效应方面的研究较少,但是拉曼光谱所蕴含的信息更加丰富,应着重发展拉曼光谱在污染物毒性效应方面的应用。
4 研究展望(Research outlook)
(1)加强将红外和拉曼光谱有机联合应用,目前将两者联合用于污染物毒性效应的研究较少。因为许多在红外光谱中较弱的波段是拉曼光谱中最强的波段,所以在污染物毒性效应研究中各有各的优势,可以互补。通过有机地结合红外和拉曼光谱的测定结果,能更好地反映污染物的毒性效应。
(2)拓展红外和拉曼光谱污染物毒性效应研究的应用范围。一方面是样品类型的拓展,如在植物应用领域,目前基本上只运用在绿藻和浮萍等植物中,应进一步拓展到高等植物(农作物、蔬菜等)上;另一方面是目标污染物的拓展,如微塑料等新型污染物的研究。
(3)相对于红外光谱而言,目前拉曼光谱的应用较少,特别是表面增强拉曼光谱的应用,拉曼光谱中所含有的生化特征要远远高于红外光谱,因此,发展新的表面增强技术,特别是具有特异性指标的表面增强拉曼光谱技术,将是一个重要的研究方向。
(4)目前的研究大都侧重于单一污染物的研究,但是环境中的污染大都以复合污染的形式存在,因此,应加强将生物光谱技术应用于复合污染的研究之中。
