不同光照条件对虎皮兰气孔特征及气体交换参数的影响
2018-03-14郭丽丽张运鑫郝立华胡晔晖郑云普
郭丽丽, 张运鑫, 张 浩, 郝立华, 李 菲, 聂 唯, 杨 彬, 胡晔晖, 曹 旭, 郑云普,3
(1.河北工程大学水电学院,河北邯郸 056038; 2.邯郸学院生命科学与技术学院,河北邯郸 056005; 3.中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101; 4.中国农业大学水利与土木工程学院,北京 100083; 5.中国水利水电科学研究院水利研究所,北京 100048)
气孔为植物叶片上由2个保卫细胞组成的孔状结构[1-2],是植物与大气之间进行CO2和水分交换的重要器官[3],对于调节生态系统碳、水循环过程具有极其重要的作用和意义[4-6]。众多研究结果表明,气孔能够根据环境因素的变化调整气孔开度、气孔大小、气孔频度(气孔密度和气孔指数)以及空间分布格局来最优化其气体交换参数,进一步提高其抗逆能力[7-8]。植物叶片气孔的发育过程受许多因素的影响,如转录因子、负调控因子、环境因素(温度、CO2、光照、激素等)均参与和影响着气孔的发育过程,且在各个发育阶段承担着关键的调节作用[9-12]。
通常而言,植物叶片的光合作用受光照、二氧化碳浓度、温度、矿质元素、水分等因素影响[13]。光能是植物进行光合作用的驱动力[14],同时光照在叶片气孔的形成和发育过程中也起到极其关键的诱导和调节作用,从而影响植物叶片的光合作用效率[10,12-13]。光照对气孔结构特征的影响机理非常复杂,至今还没有一致的结论[7,13]。相关研究结果发现,冠层浓厚树木叶片的气孔密度随光照的增强而升高,但是树冠稀薄树木的气孔密度随着光照增强而降低[15]。同一叶片在不同轴面上的气孔密度对光照变化的响应也不尽相同[16]。虎皮兰(SansevieriatrifasciataPrain)隶属于百合科虎尾兰属,为多年生常绿草本植物,以其易繁殖、生长速度快、吸收室内甲醛和室外汽车尾气等优点逐渐成为一种集观赏、绿化和人体保健于一体的重要园艺植物[17-18]。近年来,尽管许多学者对于虎皮兰的食用价值和空气甲醛净化能力等方面进行了初步的研究[19-20],但光照如何通过改变虎皮兰的气孔结构及分布状况,影响虎皮兰的气体交换过程至今没有得到较好的结论。本试验通过不同遮阴处理研究光照对虎皮兰气孔特征及空间分布格局的影响,旨在阐明光照条件下气孔特性和气体交换参数之间的关系。
1 材料与方法
1.1 试验材料
在花卉市场购买长势相同、生长健壮的虎皮兰9盆(每盆3棵)。塑料花盆高10 cm,直径14 cm,盆底铺无纺布以防浇水时基质外渗,盆栽基质为花卉栽培通用营养土,主要成分配比为氮(N)600 mg/L、磷酐(P2O5)270 mg/L、氧化钾(K2O)360 mg/L。
1.2 试验设计与实施
将塑料遮阳网剪成1 m×1 m的正方形用于虎皮兰的遮阴处理,将上述购买的虎皮兰分别置于不同的光照条件下进行处理。于2016年3月10日至4月10日,试验期间每隔 4 d 测定1次,观测时间为09:00—11:00,每株虎皮兰取3张叶片观测。利用光度计(型号759S,棱光科技有限公司,中国上海)测定不同遮阴条件下虎皮兰冠层的光照度,即无遮挡的自然光强度代表100%光照[约1 000 μmol/(m2·s)PAR],利用1层遮阳网遮挡的自然光强度为50%光照[500 μmol/(m2·s)PAR],2层遮阳网遮挡的自然光强度为25%光照[250 μmol/(m2·s)PAR]。在培养期间利用温度计和湿度计每天记录试验处理区的日平均气温和相对空气湿度,保证不同光照处理间的温度、相对空气湿度一致。培养期间每3 d浇水1次(每盆约500 mL),以防止干旱胁迫的发生。
1.3 印记法取样及光学显微镜观察
随机从每株上选取3张叶片,利用无色透明的指甲油涂于虎皮兰叶片的中部,采集气孔印记样品用于叶片气孔参数的测量。将叶片印迹玻片在装备有照相机的显微镜(DM2500,LeicaCorp,Germany)下观察并拍照。随机选择3个不同的显微视野,每个视野下拍3张照片,即得到9张气孔的显微照片,再从9张照片中选取5张来计算气孔密度。利用AutoCAD 2010软件分别测量气孔的长度、宽度、周长、面积以及形状指数。形状指数是指通过计算单一气孔形状与相同面积的圆之间的偏离程度来测量,当叶片气孔的形状为圆形时,形状指数为1,即气孔的形状越扁长,则气孔形状指数就越大。形状指数的计算公式为:

(1)
式中:S表示叶片气孔形状指数;P表示气孔周长;A表示气孔面积。
1.4 气孔的空间格局分析
随机选取3个光学显微照片(放大5倍)用于研究遮阴对叶片气孔空间分布格局的影响,每个气孔都是叶片表面上分布的单点,气孔开口的最中间位置为该单点的位置。首先利用空间分布软件ArcGIS 10.0将显微照片相同的坐标下进行数字化处理,得到每个气孔的坐标值。再运用空间统计分析方法Ripley’s K方程对表征气孔分布状况的点进行空间分析[21]。Ripley’s K方程是一个分布累加的函数,该函数利用所有单点距离的二阶矩阵探究这些点在不同尺度上的二维空间分布格局。分析结果由最小邻域距离Lhat(d)值来表达:
(2)
式中:K(d)表示任何个体在一定空间尺度内对其他个体的期望值;d表示空间尺度。
当分布格局为随机分布时,所有的d值到的距离均相等。为了确定95%的可信任区间,采用蒙特卡洛算法模拟随机分布点1 000次。假如叶片表面的气孔在给定尺度d下为随机分布,则计算出来的值应该位于95%可信任区间之内。假如值大于95%可信任区间,则气孔在该尺度为簇状分布。否则,当值小于95%可信任区间时,气孔在该尺度下为规则分布[21]。
1.5 气体交换参数测定
利用便携式光合测定系统(Lcpro-SD,ADC BioScientific Ltd. Hoddesdon,UK)测定虎皮兰叶片的气体交换参数即叶片的净光合速率(Pn)、气孔导度(Gs)、细胞间CO2浓度(Ci)、蒸腾速率(Tr)。利用光合测定系统分别测量不同光照处理下3盆虎皮兰叶片的气体交换参数,共测8次,并分别对8组数据求出平均值代表气体交换参数。
1.6 统计分析
试验数据均利用单因素方差分析,然后在利用Duncans Multiple Range Test比较处理间的差异显著性,统计分析均利用SPSS13.0(Chicago,IL)统计软件完成。
2 结果与分析
2.1 不同光照条件对气孔密度的影响
从表1可以看出,不同的光照度对虎皮兰的气孔密度产生很大影响,随着光照的减弱,虎皮兰的气孔密度逐渐增加,在25%光照下气孔密度最高。50%光照比100%光照时虎皮兰叶片的气孔个数增加15.9%,而25%光照比100%光照的气孔个数增加63.5%。结果表明,不同光照处理对虎皮兰叶片上气孔个数产生了显著影响。25%光照与100%光照下的气孔密度差异显著,但在50%光照与25%、100%光照处理间的气孔密度差异不显著。

表1 不同光照条件对虎皮兰叶片气孔特征的影响
注:同列数据后不同小写字母表示差异显著(P<0.05)。
2.2 不同光照条件对气孔开度和形状的影响
虎皮兰叶片上气孔的长度、宽度、面积、周长均随光照的减弱呈先上升后降低的趋势,而虎皮兰叶片上的气孔形状指数随光照的增加呈先降低后上升的趋势(表1)。在50%光照下的气孔长度和25%下的气孔长度有显著差异,而100%光照下的气孔长度与50%、25%光照处理间差异不显著。不同光照处理对气孔宽度、气孔面积、气孔周长及形状面积处理间差异不显著。气孔面积指数(SAAI)随着光照程度的加深而增大,25%光照下的SAAI最高,比100%光照下的SAAI增加了59%,虎皮兰叶片在不同光照处理间SAAI差异显著。
2.3 不同光照条件对气孔空间分布格局的影响
不同光照处理,虎皮兰的气孔在小尺度范围80~130 μm为规则分布,而在大尺度范围400~500 μm呈现随机分布(图1)。在100%光照下,虎皮兰的气孔分布比50%、25%光照的气孔分布更加规则,因为在相同尺度下100%光照的Lhat(d)值更小。25%光照下的气孔分布格局在小尺度约100 μm时为随机分布,50%光照下的气孔分布在尺度 230 μm 左右为随机分布,而后随着尺度的不断增大,25%、50%光照下的气孔在随机分布和规则分布之间切换。而100%光照下的气孔分布在更大尺度400 μm左右显示为随机分布。在尺度为430~500 μm时不同的光照处理结果,气孔的空间分布均为随机分布。对照不同光照处理下虎皮兰气孔分布格局最规则的空间分布格局发生在尺度约170 μm处,此时100%光照的最小Lhat(d)为-18,50%光照的最小Lhat(d)为-14.20,25%光照的最小Lhat(d)为-9.82。

2.4 不同光照条件对叶片气体交换参数的影响
虎皮兰叶片的净光合速率在25%光照下最高,其次是50%光照,而在100%光照下最低,且差异显著。随着光照度的加深,虎皮兰叶片的细胞间CO2浓度、蒸腾速率均是先升高后降低,在50%光照下达到最高,胞间CO2浓度50%光照处理与25%、100%光照处理间差异显著,蒸腾速率25%、50%、100%光照处理间差异显著。而虎皮兰在3种不同的光照处理下气孔导度差异不显著(表2)。表明蒸腾速率、净光合速率的改变并不受气孔导度影响,而是受气孔宽度、气孔长度等其他参数的影响。
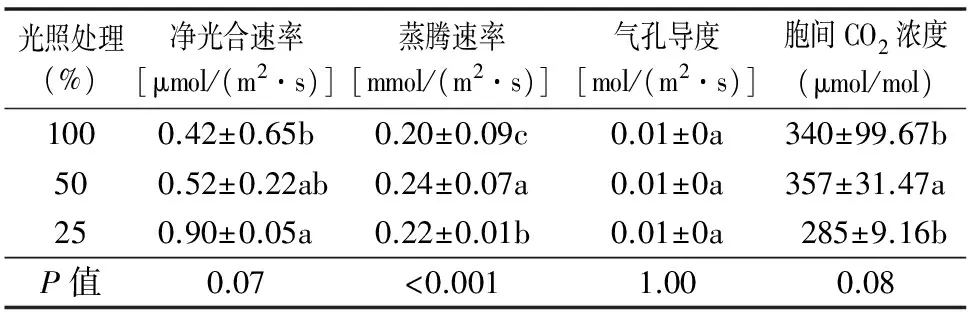
表2 不同光照条件对虎皮兰叶片气体交换参数的影响
注:同列数据后不同小写字母表示差异显著(P<0.05)。
3 讨论与结论
3.1 不同光照处理对虎皮兰叶片气孔特征的影响
气孔是植物叶片上许多小的开口[22],它是植物与外界环境进行CO2气体和水分交换的主要通道[9,23]。植物通过改变气孔开度来响应短期的环境变化,这种响应方式也被称为气孔运动[24-26]。除了对短期环境变化响应的气孔运动外,长期的环境变化如气候变暖也可能会影响到单个气孔的大小、气孔频度以及气孔空间分布格局[27-28]。目前,有关光照对植物叶片气孔频度及大小等方面影响研究还没有一致的结论。前人研究结果表明,气孔密度随着光照度的增加而升高,且冠层顶部叶片的气孔密度高于冠层底部叶片[29-30]。孟雷等的研究发现,光照度导致水稻叶片气孔密度减少,遮阴处理使水稻叶片气孔密度显著减少[31]。张大鹏研究指出,弱光可能有利于植物叶原基阶段气孔发育和形成,从而提高植物叶片的气孔密度[32]。相关研究还发现,同一物种不同轴面气孔密度对光照的响应也不尽相同。Mott等研究发现,豚草(AmbrosiaartemisiifoliaL.)在遮阴环境下近轴面气孔密度会随着光照的增强而降低,而远轴面气孔密度随着光照的增强而增加[33]。此外,光照对气孔密度的影响还同植物的生长状况存在密切的关系。马冬雪等对不同光照条件下HB柚(CitrusmaximaMerr)气孔的特征进行研究,发现气孔密度与树木的生长状况存在密切关系,生长状况较好树木的气孔密度会随着光照的增强而增加,生长状况较差树木的气孔密度则随着光照的增强而减少[15]。本研究通过对虎皮兰不同的遮阴程度发现,虎皮兰叶片气孔密度随着光照的减弱而增加,表明光照是虎皮兰气孔密度的负调控因子。前人研究结果发现,植物叶片上气孔个数由气孔发育过程所决定,而气孔的发育和形成过程又会受多种转录因子的调控,包括bHLH转录因子、MYB转录因子、Dof转录因子等[9-10]。同时,SPCH、MUTE、FAMA等3种基因均决定着气孔的发育和形成过程[34-35]。总之,光照通过影响转录因子、受体、激酶和蛋白等来调控气孔的形成和发育过程。然而,光因子对气孔发育过程产生影响的分子调控机制至今还并不清楚,将成为未来气孔发育学亟待解决的热点和难点问题之一。
3.2 不同光照处理对虎皮兰叶片气孔空间分布格局的影响
气体交换效率除受气孔大小、气孔宽度以及气孔频度以外,还与气孔分布格局有关[3,36]。前人研究发现,细胞分裂和分化过程影响和改变植物叶片上气孔的空间分布格局。同时,细胞的分裂和分化过程也会受到遗传信号和环境因素的影响[37-40]。基因学方面关于气孔的研究发现,SDDI、EPFI、TMM和ERECTA等基因共同决定着气孔的空间分布格局[9,34]。光照不仅会改变气孔的大小、形状指数以及气孔密度,同时还会影响气孔的空间分布格局。本研究发现,光照度越高,气孔分布越规则,100%光照下的气孔分布格局在大尺度(约400 μm)时为随机分布,而25%光照下的气孔分布格局在小尺度内(约100 μm)呈现为随机分布,表明随着光照的增强可能会提高虎皮兰叶片的气孔交换效率,这主要是由于气孔在植物叶片上分布得越规则,叶片的气体交换效率可能会越高。本研究中25%、50%光照条件下,气孔在100~400 μm 空间尺度内以随机和规则2种分布格局交替出现,这可能是虎皮兰在通过改变自身的气孔分布来逐渐适应不同的光照环境,从而提高其气体交换的效率的原因。
3.3 不同光照处理对虎皮兰气体交换参数的影响
众所周知,植物的光合作用过程会受到光照度的影响[39-40],光照度太高或太低都会降低植物光合反应速率,即植物的光合反应速率只有在最佳光照度下才能达到最大值[41-43]。吴安娜等研究发现,光照度增强有利于月季试管苗干物质的积累,但该效应仅在一定光照度范围内产生,若超过该光照范围就会对植物生长产生负效应[44]。Lees等研究发现,铁线莲试管苗随着光照的增强,光合作用和量子小效率均降低,可能是太高的光照对植物进行光合作用的器官造成损伤[45]。另外,光照对植物光合作用过程的影响还会通过气孔导度的变化来实现[46-47]。Whitehead等的研究发现,对火炬松进行遮光处理导致其气孔导度降低39%,从而使光合反应速率也下降58%[48]。马书荣等的研究也发现,裂叶沙参(AdenophoralobophyllaHong)光合速率与气孔导度的相关系数极显著[49]。本研究却发现不同光照度对虎皮兰的气孔导度没有产生影响。这可能是由于气孔面积指数(单位叶面积上气孔的总面积)的增加同气孔空间分布格局变得不规则的效应相抵消,以至于气孔结构和分布状况并未对气孔导度产生影响。
参考文献:
[1]Woodward F I. Stomatal numbers are sensitive to increases in CO2from pre-industrial levels[J]. Nature,1987,327(6123):617-618.
[2]Bergmann D C,Sack F D. Stomatal development[J]. Annual Review of Plant Biology,2007,58(1):163-181.
[3]Hetherington A M,Woodward F I. The role of stomata in sensing and driving environmental change[J]. Nature,2003,424(6951):901-908.
[4]Franks P J,Beerling D J. Maximum leaf conductance driven by CO2effects on stomatal size and density over geologic time[J]. Proceedings of the National Academy of Sciences of the United States of America,2009,106(25):10343-10347.
[5]Haworth M,Heath J,Mcelwain J C. Differences in the response sensitivity of stomatal index to atmospheric CO2among four genera ofCupressaceaeconifers[J]. Annals of Botany,2010,105(3):411-418.
[6]Taylor S H,Franks P J,Hulme S P,et al. Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses[J]. New Phytologist,2012,193(2):387-396.
[7]Kouwenberg L L R,Kürschner W M,Mcelwain J C. Stomatal frequency change over altitudinal gradients:prospects for Paleoaltimetry[J]. Reviews in Mineralogy & Geochemistry,2007,66(12):215-241.
[8]Fraser L H,Greenall A,Carlyle C,et al. Adaptive phenotypic plasticity ofPseudoroegneriaspicata:response of stomatal density,leaf area and biomass to changes in water supply and increased temperature[J]. Annals of Botany,2009,103(5):769-775.
[9]崔国新,韩宝达,赵潇男,等. 气孔发育及其调控[J]. 植物生理学报,2012,48(9):829-836.
[10]吴 丁. 气孔的构造及类型在生理上的意义[J]. 九江学院学报(社会科学版),1997(6):39-43.
[11]刘 俊,郭志富. 植物气孔发育分子机制研究进展[J]. 安微农业科学,2015,43(35):12-15.
[12]刘 婧,王宝山,谢先芝. 植物气孔发育及其调控研究[J]. 遗传,2011,33(2):131-137.
[13]Berry J,Bjorkman O. Photosynthetic response and adaptation to temperature in higher plants[J]. Plant Physiology,1980(31):491-543
[14]Pallardy S G,Kozlowski T T. Stomatal response of populus clones to light Intensity and vapor pressure deficit[J]. Plant Physiology,1979(64):112-114.
[15]马冬雪,刘仁林. 不同光照条件下HB柚气孔数量特征研究[J]. 安微农业科学,2011,39(19):6983-6985.
[16]蔡志全,齐 欣,曹坤芳. 七种热带雨林树苗叶片气孔特征及其可塑性对不同光照强度的响应[J]. 应用生态学报,2004,15(2):201-204.
[17]李泽鸿,刘树英,安佰明,等. 虎皮兰色素提取条件的研究[J]. 北方园艺,2008(1):134-136.
[18]郭铭伟,垄 萍,陶荣琴,等. 盆栽芦荟和虎皮兰对室内空气甲醛净化及耐受能力的研究[J]. 实用预防医学,2012,19(10):1462-1464.
[19]李泽鸿,刘树英,李 闯,等. 虎皮兰营养成分的测定[J]. 北方园艺,2008(2):131-132.
[20]熊 纓,苏志刚. 五种常见装饰植物对甲醛的吸收能力比较研究[J]. 环境科学与管理,2009,34(1):45-48.
[21]Zheng Y,Xu M,Hou R,et al. Effects of experimental warming on stomatal traits in leaves of maize (ZeaMayL.)[J]. Ecology and Evolution,2013,3(9):3095-3111.
[22]Xu M. The optimal atmospheric CO2concentration for the growth of winter wheat (Triticumaestivum)[J]. Plant Physiology,2015(184):89-97.
[23]Young J J,Mehta S,Israelsson M,et al. CO2signaling in guard cells:calcium sensitivity response modulation,a Ca2+-independent phase,and CO2insensitivity of thegca2 mutant[J]. Proceedings of the National Academy of Sciences of the United States of America,2006,103(19):7506-7511.
[24]Shimazaki K I,Doi M,Assmann S M,et al. Light regulation of stomatal movement[J]. Annual Review of Plant Biology,2007(58):219-247.
[25]Shang Z,Laohavisit A,Davies J M. Extracellular ATP activates an Arabidopsis plasma membrane Ca2+-permeable conductance[J]. Plant Signaling & Behavior,2009,4(10):989-991.
[26]Anderson V J,Brisk D D. Stomatal distribution,density and conductance of three perennial grasses native to the southern true trairie of Texas[J]. American Midland Naturalist,1990(123):152-159.
[27]Lammertsma E I,de Boer H J,Dekker S C,et al. Global CO2rise leads to reduced maximum stomatal conductance in Florida vegetation[J]. Proceedings of the National Academy of Sciences of the United States of America,2011,108(10):4035-4040.
[28]山本昭夫. 土壤水分和光照强度对气孔密度的影响[J]. 日本作物学会纪事,1984,53(4):463-471.
[29]Ciha A J,Bron W A. Stomatal size and frequency in soybeans[J]. Crop Science,1975(15):309-313.
[30]Chen W,Cheng H,Liu L,et al. A new method for studying the stomatal character of rice leaf[J]. Review of China Agricultural Science of Technology,2000,2(2):58-62.
[31]孟 雷,陈温福,李磊鑫,等. 减弱光照强度对水稻叶片气孔性状的影响[J]. 沈阳农业大学学报,2002,33(2):87-89.
[32]张大鹏. 水稻叶片气孔的研究Ⅱ.不同生态条件下的气孔动态[J]. 福建农学院学报,1989,18(3):302-307.
[33]Mott K A,Gibson A C,O’leary J W. The adaptive significance of atmphistomatic leaves[J]. Plant,Cell & Environment,1982(5):455-460.
[34]朱燕化,康宏樟,刘春江. 植物叶片气孔性状变异的影响及其研究方法[J]. 应用生态学报,2012,2(1):250-256.
[35]林植芳,李双顺,林桂珠. 叶片气孔的分布于光合途径[J]. 植物学报,1986(4):153-160.
[36]Wang J C,Duan B L,Zhang Y B. Effects of experimental warming on growth,biomass allocation,and needle chemistry ofAbiesfaxonianain even-aged monospecific stands[J]. Plant Ecology,2012,213(1):47-55.
[37]Shpak E D,Mcabee J M,Pillitteri L J,et al. Stomatal patterning and differentiation by synergistic interactions of receptor kinases[J]. Science,2005,309(5732):290-293.
[38]Nadeau J A,Sack F D. Control of stomatal distribution on the Arabidopsis leaf surface[J]. Science,2002,296(5573):1697-1700.
[39]Juarez M T,Twigg R W,Timmermans M C. Specification of adaxial cell fate during maize leaf development[J]. Development,2004,131(18):4533-4544.
[40]Wang H,Ngwenyama N,Liu Y,et al. Stomatal development and patterning are regulated by environmentally responsive mitogen-activated protein kinases inArabidopsis[J]. The Plant Cell,2007,19(1):63-73.
[41]Hunt L,Bailey K J,Gray J E. The signalling peptide EPFL9 is a positive regulator of stomatal development[J]. New Phytologist,2010,186(3):609-614.
[42]何松林,蒋要卫,王 政,等. 外施CO2时光照度对大花蕙兰试管苗生长和叶片气孔特征的影响[J]. 河南农业大学学报,2009,43(6):625-629.
[43]王 政,刘艺平,张春丽,等. LED不同光照度对大花蕙兰试管苗生长和叶片气孔特征的影响[C]// 中国观赏园艺研究进展. 2013:324-328.
[44]吴安娜,朱建镛. 光和蔗糖对玫瑰花组织培养培殖体光自养生长的影响[J]. 中国园艺,1999,45(4):345-352.
[45]Lees R P,Evanse H,Nicholas J R. Effects of the light environment on photosynthesis and growthinvitro[M]. Netherlands:Springer,1994.
[46]Woods D B,Turner N C. Stomatal response to changing light by four tree species of varying shade tolerance[J]. New Phytologist,1971(70):77-84.
[47]Liu T,Ohashi-Ito K,Bergmann D C. Orthologs ofArabidopsisthalianastomatalbHLHgenes and regulation of stomatal development in grasses[J]. Development,2009,136(13):2265-2276.
[48]Whitehead D,Teskey R O. Dynamic response of stomata to changing irradiance in loblolly pine (PinustaedaL.)[J]. Tree Physiology,1995,15(4):245-251.
[49]马书荣,杨红艺,阎秀峰,等. 不同光照处理裂叶沙参和泡参气孔行为与光合特性的关系[J]. 东北林业大学学报,2000,28(6):36-39.
