不同剂量质子辐射玉米M1代的生物诱变效应
2018-03-14胡金山田安然耿金鹏
胡金山,田安然,耿金鹏*,隋 丽,展 永
(1.河北工业大学生物物理研究所,天津300401;2.华北理工大学 化学工程学院,河北 唐山063210;3.中国原子能科学研究院,北京102413)
玉米(Zea mays L.)原产自南美洲,为1年生禾本科草本植物,是目前全世界产量最高的三大粮食作物之一,其籽粒以黄、白两色为主。玉米素有长寿食品的美称,含有丰富的蛋白质、氨基酸、脂肪酸、维生素、纤维素以及多糖等,经常食用能够预防动脉粥样硬化、心脑血管疾病、高胆固醇血症的发生[1]。玉米不仅作为粮食和饲料,还广泛用于食品、化工、医药、纺织等行业[2]。全球对玉米总量的需求日益增长。现阶段,通过提高玉米种植面积增加产量已经非常困难,因而对提高玉米单产和改善玉米品质提出了更高的要求。除气候、环境、土壤等特定因素,玉米遗传基础狭窄、种质资源相对匮乏已经成为我国玉米育种的瓶颈[3]。
辐射诱变在创新、拓展玉米种质资源方面发挥着重要作用。传统辐射源以χ、γ、β射线和中子为主,但存在育种周期长、定向突变差等问题,限制了辐射诱变育种的应用。近年来,重离子辐射诱变因其束流多样性、突变频谱宽、稳定周期短等特点开创了辐射诱变育种的新途径[4]。质子辐射(PR)尽管原子序数较低但电离密度大,也属高传能线密度辐射(LET),可将自身所携带能量沉积在其径迹上,产生的相对生物学效应(RBE)较χ、γ、e等低LET辐射明显,而其引起的生理损伤却明显小于7Li、12C等重离子[5-6]。20世纪90年代,我国科研人员尝试用加速的高能质子辐射诱变植物种子,相继取得了一系列成果。唐掌雄等[7]用质子辐射麦类,结果发现,M1代苗高和根长的变化分别与辐照剂量及射线能量有关。王彩莲等[8]用质子辐射水稻,获得了矮秆和千粒质量显著增加的突变株。施巾帼等[9]研究了质子辐射对小麦的诱变效应,结果发现,不同剂量质子辐射对小麦M1代的有益变异效果不同。质子辐射突变频率高,可以显著加快育种进程,但现有文献报道对玉米的辐射诱变研究大多采用γ射线[10]、重离子[5-6]以及航天搭载[11]等方式,研究重点主要集中在细胞基因组的生理损伤[12]、生物诱变效应[5-6]、突变体的营养品质[13-14]以及构建理论模型[15]等方面,而关于质子辐射诱变玉米的研究却鲜见报道。鉴于此,利用不同剂量质子辐射处理玉米种子,研究其对发芽率、成苗率、空秆率、株高、穗位高以及千粒质量等生物性状的影响,同时采用随机扩增多态性DNA(RAPD)分子标记技术研究M1代的突变率并进行遗传相似聚类分析,系统研究质子辐射对玉米的生物诱变效应,初步筛选有益变异株,为提高质子辐射效果和选育玉米新品种提供依据。
1 材料和方法
1.1 供试材料及质子辐射处理
供试玉米品种为白马牙玉米,白马牙玉米因籽粒白色、颗粒偏平而得名,为河北迁安当地传统种植品种。采用中国原子能科学研究院的HI-13串列加速器对白马牙玉米干种子进行质子辐射,辐射能量为 13.00 MeV/u,传能线密度为 3.50 keV/μm,辐射剂量分别为 0(CK)、10、20、30、40、50 Gy,每个剂量处理种子50粒,所有辐射试验均在正常大气条件下进行。
1.2 田间试验及表观性状监测
田间试验于2016年在河北省迁安市隆兴农业科技示范场进行,浅耕施基肥,人工开穴单粒点播。不同剂量质子辐射处理分别播种50粒种子(株距0.25 m,行距 0.50 m)。根据植株生长情况定期灌溉,追肥尿素、磷酸二铵一次。观测发芽率、成苗率、空秆率、叶型、株高、穗位高、千粒质量等表观性状以及抗病性。重点统计M1代出苗期第15天的发芽率、拔节期第5天的成苗率以及抽雄吐丝期第7天的空秆率(n=50),随机统计完熟期收获前玉米植株的株高、穗位高、千粒质量等表观性状指标(n=20),以未经质子辐射处理的M1代玉米表观性状指标作为对照(CK)。
1.3 光合速率测定
在玉米抽雄吐丝期的第8天同一时间段内环境温度相对稳定条件下,采用ECA-PB0402光合速率测定仪(北京益康农科技有限公司生产)测定M1代玉米穗位上部第1片叶的光合速率,各辐射剂量处理及CK随机测量20株长势较为接近的植株,取平均值。
1.4 RAPD 分析
DNA提取及相关检测:采用 Tiangen公司的DNA提取试剂盒,根据具体步骤提取CK及各辐射剂量处理M1代玉米抽雄吐丝期叶片DNA,采用凝胶电泳成像系统(1%琼脂糖凝胶)检测。采用UV-765紫外可见分光光度计对样品逐一进行吸光度检测。
PCR扩增:采用美国Bio公司的荧光定量PCR仪进行DNA扩增,扩增程序按95℃ 5 min,37℃1 min,72 ℃ 1.5 min,共 35 个循环,额外 72 ℃10 min用于完全延伸。扩增产物在1.5%琼脂糖凝胶中进行电泳分离,1×TAE缓冲液含有EB,最后用凝胶成像系统拍照。
PCR体系优化及引物筛选:由于PCR体系会对RAPD结果的稳定性产生影响,为保证扩增的准确、可靠以及可重复性,对PCR体系进行一系列优化。优化体系如下:采用 25 μL 体系,含 10.8 μL 超纯水、2.5 μL buffer、2.5 μL Dye、2 μL dNTP、4 μL 引物、1.2 μL Taq 酶、2 μL DNA 模板。以对照 DNA 为模板,选用142对随机引物在优化体系下进行扩增,从中选出条带稳定性好、清晰度高、带型丰富的随机引物。
1.5 数据处理与分析
采用SPSS软件的单因素方差分析和LSD多重比较进行数据分析。各统计量定义如下:
发芽率=(发芽种子粒数/供试种子粒数)×100%,
成苗率=(成苗株数/供试种子粒数)×100%,
空秆率=(空秆株数/成活总株数)×100%,
多态性比率=(多态性条带数/对照扩增条带数)×100%。
根据RAPD电泳结果统计,缺失或增加条带为阳性记为1,与对照条带相同扩增阴性记为0。采用NTsys软件对不同剂量处理及CK进行聚类分析。
2 结果与分析
观察田间表观性状发现,质子辐射诱变的白马牙玉米M1代在发芽、成苗、抽雄、开花、散粉、吐丝、成熟等阶段都有变异现象出现。20 Gy质子辐射处理有3株叶片出现皱缩、卷曲,40 Gy质子辐射处理有5株叶片变宽,但上述2个处理的穗上叶片数和总叶片数与CK没有明显差异。10 Gy质子辐射处理有1株出现雄性不育,2株花药由紫色变为黄色,2株花丝由红色变为粉色。另外,50 Gy质子辐射处理有4株出现同位多穗现象。10 Gy质子辐射处理抗病性整体增强,但20、30 Gy质子辐射处理抗倒伏能力整体减弱,抗旱性没有明显变化。10 Gy质子辐射处理有6株抽雄期、吐丝期均比CK提前3 d,成熟期也相应提前且活秆成熟,而50 Gy质子辐射处理有3株成熟期却比CK晚2 d。
2.1 质子辐射对玉米M1代发芽率、成苗率和空秆率的影响
玉米发芽率能够直观地显示种子的萌发出芽状况,而成苗率则反映植株的成苗情况。如图1所示,不同剂量质子辐射处理发芽率均在86%以上,其中,20 Gy处理出苗情况最好,发芽率高达98%,辐射剂量继续增加,发芽率总体呈降低趋势。质子辐射各组成苗率均在84%以上,整体变化趋势与发芽率相似,但略低于发芽率。空秆率直接影响玉米产量,统计空秆率对准确估算玉米产量有重要意义。与CK相比,10、40 Gy处理诱变的空秆率分别为13.04%、20.93%,而 20、50 Gy 处理空秆率则明显较低(2%左右),不同剂量质子辐射处理M1代空秆率随辐射剂量的变化呈典型的马鞍形双峰曲线。
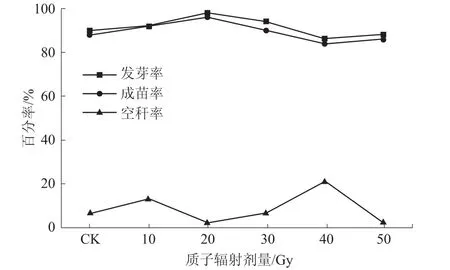
图1 不同剂量质子辐射对玉米M1代发芽率、成苗率和空秆率的影响
2.2 质子辐射对玉米M1代株高、穗位高、千粒质量的影响
相对穗位(穗位高与株高之比)能在一定程度上反映植株的抗倒伏能力,千粒质量则是体现玉米籽粒大小与饱满程度的一项指标,也是田间试验预测产量的重要依据。由表1可知,与CK相比,10、20、40、50 Gy处理玉米株高均显著增加,而30 Gy处理显著降低。穗位高只有20 Gy处理显著增加,整体而言,质子辐射对株高的影响大于对穗位高的影响。20、30 Gy处理玉米植株的相对穗位均明显增高,提示上述2个剂量处理的植株抗倒伏能力减弱。图2所示为不同剂量质子辐射M1代玉米果穗,10 Gy 处理玉米千粒质量高达 654 g,20、40、50 Gy 处理玉米千粒质量也均显著增加(表1),随辐射剂量增加玉米千粒质量也呈典型的马鞍形双峰曲线变化,提示质子辐射对提高白马牙玉米产量有潜在的作用。

表1 不同剂量质子辐射对玉米M1代株高、穗位高和千粒质量的影响

图2 不同剂量质子辐射处理M1代玉米果穗
2.3 质子辐射对玉米M1代光合速率的影响
光合作用能够提供玉米生长发育所需物质和能量,是玉米产量形成的基础。由图3可见,不同剂量质子辐射处理的白马牙玉米M1代光合速率随辐射剂量增加呈典型的马鞍形曲线变化。其中,10 Gy处理白马牙玉米M1代的光合速率最高,达23.48 μmol/(m2·s),其次是 20、40 Gy 处理,分别为22.01、21.87 μmol/(m2·s),3 个处理均显著高于CK[19.72 μmol/(m2·s)],而 30 Gy 处理[18.96 μmol/(m2·s)]略低于 CK,50 Gy 处理[19.82 μmol/(m2·s)]基本与CK持平。
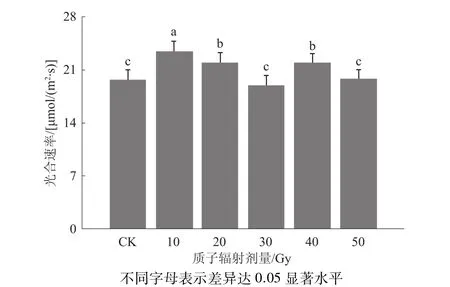
图3 不同剂量质子辐射对玉米光合速率的影响
2.4 玉米突变的RAPD分析
在优化反应体系下,从142对随机引物中筛选出16对(表2),共扩增出124个条带,扩增产物片段大小均在200~3 000 bp。不同剂量质子辐射处理玉米基因组DNA均出现不同程度的多态性,主要表现为电泳条带的增加或缺失。从图4可见,与CK相比,引物A2中箭头所指10、20 Gy处理在800~1 200 bp各增加1个条带,而50 Gy处理在800~1 200 bp增加不同的2个条带;引物A10中箭头所指50 Gy处理在800~1 200 bp缺失1个条带,10、50 Gy处理在500~800 bp增加相同的1个条带,而20、30、40 Gy处理也增加相同的1个条带。

表2 RAPD分析选用随机引物序列
为系统研究质子辐射诱变对玉米基因组的影响,采用RAPD分别统计多态性比率(表3),不同剂量质子辐射诱变的多态性比率为 20.16% ~32.26%,10、50 Gy处理多态性比率明显高于其他剂量处理组,20 Gy处理多态性比率最低,多态性比率呈现非线性的变化趋势,由此可见,质子辐射能够有效引起玉米基因组的变异。利用NTsys软件基于遗传相似度对不同剂量质子辐射处理进行聚类分析可知(图5),整个质子辐射群体在遗传相似系数0.69附近聚为两大类,其中20、30、40 Gy诱变处理与CK聚为一类,10、50 Gy处理诱变效果相近聚为另一类。

图4 随机引物A2、A10扩增结果

表3 不同剂量质子辐射对玉米基因组DNA多态性的影响

图5 不同剂量质子辐射诱变的聚类分析
3 结论与讨论
本试验结果表明,质子辐射能够引起白马牙玉米M1代明显的诱变效应。不同剂量质子辐射处理白马牙玉米M1代的发芽率、成苗率均在84%以上,可见,低剂量质子辐射处理对以上指标影响不大,但质子辐射对玉米植株的空秆率影响明显,空秆率随质子辐射剂量的增加呈马鞍形变化,该指标是否稳定遗传需要逐代跟踪。质子辐射对千粒质量以及光合速率的影响也较为明显,随辐射剂量增加有增加—降低—增加—降低交替出现的马鞍形趋势。耿金鹏等[16]曾报道,不同剂量重离子7Li辐射对玉米自交系永19的株高、穗位高有马鞍形剂量效应。陈学君等[6]证实,重离子12C和36Ar辐射处理玉米自交系郑58对部分表观性状有正负双重作用。韩荣飞等[15]综合能质沉积和电荷协同提出了EMC模型,从理论角度阐述了马鞍形存活率产生的机制。不同辐射试验引起的变异情况比较复杂,因粒子类型、射线能量、辐射剂量以及供试材料不同而反应不一,推测马鞍形剂量效应可能是一种普遍的辐射诱变效应。
Duncan等[17]系统研究了22个杂交玉米品种的光合特性,证明光合速率是一个可以稳定遗传的性状。于康珂等[18]探索了不同成熟度玉米叶片对高温胁迫的光合响应特征,结果发现,成熟叶片叶绿素含量显著下降,且不同玉米自交系的光合速率相差较大。本试验测定光合速率时所选植株的长势与CK相近,不同剂量质子辐射处理引起的植株长势明显增强或减弱的单株未被测量。10、20、40 Gy处理植株的光合速率显著高于CK,这可能是辐射引起玉米叶片细胞叶绿体合成的增加或突变所致,同时,对应剂量处理的千粒质量也有所增加。但唐掌雄等[7]利用质子辐射诱变麦类种子发现,M1代幼苗出现叶绿素缺失的现象。究竟质子辐射如何影响玉米叶绿体合成从而影响光合作用,并最终影响千粒质量需要更进一步研究。如果增强的植株光合速率特性和提高的千粒质量指标能够稳定遗传,那么通过对辐射诱变后代定向选择,则有可能培育出光合能力突出的玉米新品种,这对于培育高产玉米及创新种质资源具有重要意义。
自 Williams等[19]和 Welsh 等[20]提出 RAPD 分子标记以来,该技术因其快速、简便、灵敏等特点逐渐渗透于基因组研究的各个方面[21-22]。李谨等[23]应用RAPD对航天搭载辐射仙客来突变株进行分析,结果显示,太空环境对仙客来当代植株造成了显著诱变。孙晓莉等[24]采用RAPD分析获得了60Co γ射线辐照蝴蝶兰的最佳辐射诱变剂量。DNA分子水平的多态性检测是进行基因组研究的基础,也为探索质子辐射诱变玉米的理论机制提供了依据。本结果也证实,RAPD能有效检测玉米基因组的DNA变化,尤其是片段的插入或者删除。与转基因玉米不同,质子辐射诱变并没有外源基因的导入,主要是自身遗传物质的同源重组[25]。目前尚未确定引起玉米生物诱变效应的相关基因,下一步仍需要在细胞或生物大分子层面进行研究以阐明质子辐射诱变的机制。
综上所述,质子辐射能够有效引起玉米M1代的生物诱变效应,空秆率、千粒质量以及光合速率等生物性状变异明显,且均有典型的马鞍形剂量效应出现。10 Gy质子辐射显著提高了白马牙玉米的千粒质量,结合群体生物性状认为,10 Gy为最优辐射剂量。RAPD和聚类分析表明,质子辐射能够有效引起玉米基因组的变异,从而引起玉米M1代的生物诱变效应。可见,质子辐射是玉米种质创新的一种有效手段。
致谢:承蒙中国原子能科学研究院北京串列加速器核物理国家实验室和河北省迁安市隆兴农业科技示范场航天育种基地提供的帮助,特此感谢!
[1] 张中东,惠国强,张红梅,等.玉米的营养及药用价值研究进展[J].玉米科学,2006,14(3):173-176.
[2] 侯旭光.内蒙古玉米产业化发展现状与对策[J].河南农业科学,2006(11):15-18.
[3] 杨培珠,钟国祥,谢虹,等.玉米种质资源的背景与利用现状[J].中国农学通报,2011,27(5):25-28.
[4] 周章印.玉米诱变育种研究进展[J].河北农业科学,2008,12(7):54-57.
[5] Geng J,Li D,Cao T,et al.Mutagenic effects of maize inbred line irradiated by heavy ion7Li[J].Molecular Plant Breeding,2012,3(13):132-138.
[6] 陈学君,李文建,陈婧,等.不同剂量重离子辐照玉米自交系的生物学效应比较[J].原子核物理评论,2008,25(2):176-181.
[7] 唐掌雄,施巾帼,巩玲华,等.质子辐照麦类种子对苗期生长的影响[J].核农学通报,1997,18(6):260-262.
[8] 王彩莲,陈秋方,慎玫.空间环境和质子对水稻的诱变效应[J].中国核科技报告,1998(6):775-786.
[9] 施巾帼,孙国庆,李桂英,等.质子对小麦的诱变效应及作用机理研究Ⅰ.质子对小麦的诱变效应[J].核农学报,2002,16(2):65-69.
[10] 石海春,李奇,柯永培,等.60Co-γ射线对自交系R08和48-2 的诱变效应[J].西南农业学报,2011,24(5):1636-1641.
[11] 乔晓,石海春,柯永培,等.玉米航天诱变SP3株系的遗传变异分析[J].玉米科学,2012,20(3):15-21.
[12] 罗红兵,孔福全,倪嵋楠,等.重离子辐照玉米种子引起的基因组DNA变异分析[J].激光生物学报,2009,18(6):755-759.
[13] 陈学君,栗孟飞,杨德龙,等.SFE-GC/HPLC对重离子辐照育种的玉米油中五种营养物质的含量分析[J].营养学报,2012,34(5):496-500.
[14] 仲彩萍,李唯,陈学君.重离子辐照对玉米自交系营养品质的影响[J].甘肃农业大学学报,2010,45(5):29-34.
[15] 韩荣飞,吴跃进,卞坡,等.低能重离子剂量-存活率效应及其拟合模型探讨[J].原子核物理评论,2009,26(4):352-355.
[16] 耿金鹏,李多芳,曹天光,等.重离子辐射玉米自交系田间性状的统计分析[J].玉米科学,2013,21(2):66-70.
[17] Duncan W G,Hesketh J D.Net photosynthetic rates,relative leaf growth rates,and leaf numbers of 22 races of maize grown at eight temperatures[J].Crop Science,1968,8(6):670-674.
[18] 于康珂,孙宁宁,齐红志,等.不同成熟度玉米叶片光合生理对高温胁迫的响应特征及基因型差异[J].河南农业科学,2017,46(5):34-38.
[19] Williams J G K,Kubelik A R,Livak K J,et al.DNA polymorphisms amplified by arbitrary primers are useful as genetic markers[J].Nucleic Acids Research,1990,18(22):6531-6535.
[20] Welsh J,McClelland M.Fingerprinting genomes using PCR with arbitrary primers[J].Nucleic Acids Research,1990,18(24):7213-7218.
[21] Liu P,Zhang B,Zhao H,et al.Analysis of genetic relationship among celery germplasms by RAPD[J].Agricultural Biotechnology,2014,3(2):10-13.
[22] 李多芳,田安然,耿金鹏,等.航天搭载诱导百日草的生物学效应[J].北方园艺,2015(23):97-100.
[23] 李谨,耿金鹏,曹天光,等.太空环境对仙客来诱变效应的研究[J].北方园艺,2015(4):112-115.
[24] 孙晓莉,章铁,刘秀清.蝴蝶兰60Co γ射线辐照材料的RAPD 分子标记研究[J].中国农学通报,2009,25(1):156-159.
[25] 崔宁波,张正岩,张曲薇.转基因玉米材料的选育及产业化研究进展[J].河南农业科学,2017,46(6):13-22.
