EYA 3和TSH β在季节性发情和常年发情绵羊中的表达模式
2018-03-13张效生刘秋月王翔宇贺小云郭晓飞胡文萍张金龙储明星
夏 青,张效生,刘秋月,王翔宇,贺小云,郭晓飞,胡文萍,张金龙,储明星*,狄 冉*
(1. 中国农业科学院北京畜牧兽医研究所,农业部动物遗传育种与繁殖重点实验室,北京 100193;2. 天津市畜牧兽医研究所,天津 300381)
大部分绵羊品种具有季节性发情的特征,在长日照季节表现为乏情,短日照季节表现为发情。绵羊季节性发情导致羔羊肉不能实现四季均衡供应,严重制约了肉羊业的生产效率,因此,研究季节性发情调控机制以提高绵羊繁殖效率十分重要。目前关于该机制研究较为清楚的是动物通过甲状腺激素调节性腺生理活动进而实现对季节性发情的调控[1-2],这种机制由垂体结节部(Pars tuberalis, PT)促甲状腺细胞表达的两个长光照因子EYA 3与TSH β 启动[3-7]。
EYA基因编码一类眼缺失家族蛋白,目前已知该家族成员包含EYA 1、EYA 2、EYA 3和EYA 4,EYA因子具有磷酸酶活性,对于视网膜的发育具有重要调控作用[8-10]。其中EYA3作为昼夜节律基因对绵羊季节性发情具有间接调控作用,其参与绵羊季节性发情长光照分子通路:绵羊将接收到的外界不同长度光照信号经下丘脑视交叉上核(Hypothalamic suprachiasmatic nuclei, SCN)传递至松果体,松果体可分泌承载光周期信号的褪黑素(Melatonin, MEL),MEL可与PT区域褪黑素受体(Melatonin receptor 1, MT 1)结合,长日照下,垂体分泌的EYA 3在天黑后12 h达到表达高峰,EYA 3表达峰在早上出现,MEL水平低,EYA 3峰值较高[11-13]。此机制启动了动物机体内部一系列识别长日照信号因子的分泌机制,进而间接调控绵羊性腺轴的关闭。EYA3在不同物种组织中的表达情况已有相关研究报道,EYA3可在绵羊垂体中表达,且在LP下表达量高于SP[14];EYA3在大鼠下丘脑中表达,且LP下表达量也高于SP[15];EYA3在人类的骨髓、胚胎、肝、脊髓、肌肉、大脑和肠中均有表达[10]。已知EYA3可通过垂体间接调控性腺活动,但在不同光照条件和不同繁殖时期,EYA3是否在绵羊垂体外的组织存在表达差异,并发挥调控作用,及由SP变为LP后,绵羊EYA3表达变化趋势目前未见相关报道。
促甲状腺激素(Thyroid stimulating hormone, TSH)由糖蛋白α亚基(Glycoprotein hormone, alpha polypeptide, CGA)和β亚基(TSH-beta, TSH β)构成[16-17]。TSHβ可对光周期进行应答[3,18-19],长光照时EYA3表达升高,使PT分泌TSH β 增多,TSH β 与下丘脑基部的促甲状腺激素受体(Thyroid-stimulating hormone receptor, TSHR)结合介导了DIO 2的升高, DIO 2促使无活性的T4转化为有活性的T3,T3能够影响GnRH神经元的形态和空间位置,导致GnRH 浓度释放产生差异,最终影响垂体激素的分泌,继而调控性腺的生理活动[20-23]。研究发现,TSHβ在多种动物(大鼠、小鼠、猪、山羊、绵羊)的垂体中高表达[24-27];与EYA3类似,在不同光照条件和不同繁殖时期,各组织中TSHβ是否存在表达差异及由SP变为LP后绵羊TSHβ表达变化趋势均鲜有报道。因此,本研究明确了上述两个基因在常年发情和季节性发情绵羊品种中的表达模式,并对各组织中两基因在不同光照条件下和不同繁殖时期间的表达差异进行研究,分析了由SP转变为LP后42 d内,EYA3与TSHβ在绵羊垂体中的表达变化趋势,研究结果有助于进一步揭示两个基因在绵羊发情性状中的作用。
1 材料与方法
1.1 试验材料
1.1.1 试验动物组织样品采集 不同时期目标基因组织表达谱试验:试验羊为天津市畜牧兽医研究所畜禽繁育基地人工控光条件下饲养的健康空怀苏尼特母羊;山东郓城小尾寒羊保种场的健康空怀小尾寒羊母羊。选取短光照(人工模拟配种季节)第21 天和长光照(模拟休情季节)第49天的苏尼特羊成年母羊各3只,卵泡期和黄体期的小尾寒羊成年母羊各3只。屠宰绵羊后,迅速采集垂体、松果体、大脑、小脑、下丘脑、输卵管、子宫、卵巢、肾上腺和肾共10种新鲜组织样品,装入2 mL冻存管,置于液氮,之后储存在-80 ℃冰箱中备用。
季节性发情绵羊不同光照条件下,垂体中目标基因表达模式试验:苏尼特母羊在短光照(白天8 h:黑夜16 h)下,饲养42 d后,转至长光照(白天16 h:黑夜8 h)饲养49 d,并根据文献报道[11-12,22,28]与本实验室预试验结果,分别在SP 21、LP 3、LP 21、LP 33、LP 42共5个时间点(短光照第21天和长光照第3、21、33和42 天)屠宰绵羊,每个时间点屠宰3只羊,迅速采集垂体组织置于液氮中短暂保存后,置于-80 ℃冰箱中保存备用。
1.1.2 试剂与仪器 RNAprep pure动物组织总RNA提取试剂盒(天根生化科技北京有限公司)、PrimeScriptTMRT Reagent Kit及SYBR@Premix ExTaqTMⅡ(TaKaRa, Japan)。 PCR仪(T100型,美国Bio-rad)、超低温冰箱(美国Thermo)、电泳成像仪(Chemi Doc XRS+, 美国Bio-rad)、电泳槽(JY-SPFT,北京JUNYI)、离心机(5417型,德国Eppendorf)、荧光定量仪(罗氏480)。
1.2 方法
1.2.1 引物设计 根据NCBI数据库中绵羊EYA3基因序列(GenBank登录号:NM_001161733)与TSHβ基因序列(GenBank登录号:XM_004002368),用Primer 3软件各设计1对荧光定量引物,内参基因选用β-actin,引物信息见表1。
表1 荧光定量PCR扩增引物序列
Table 1 Primer sequences of real-time PCR
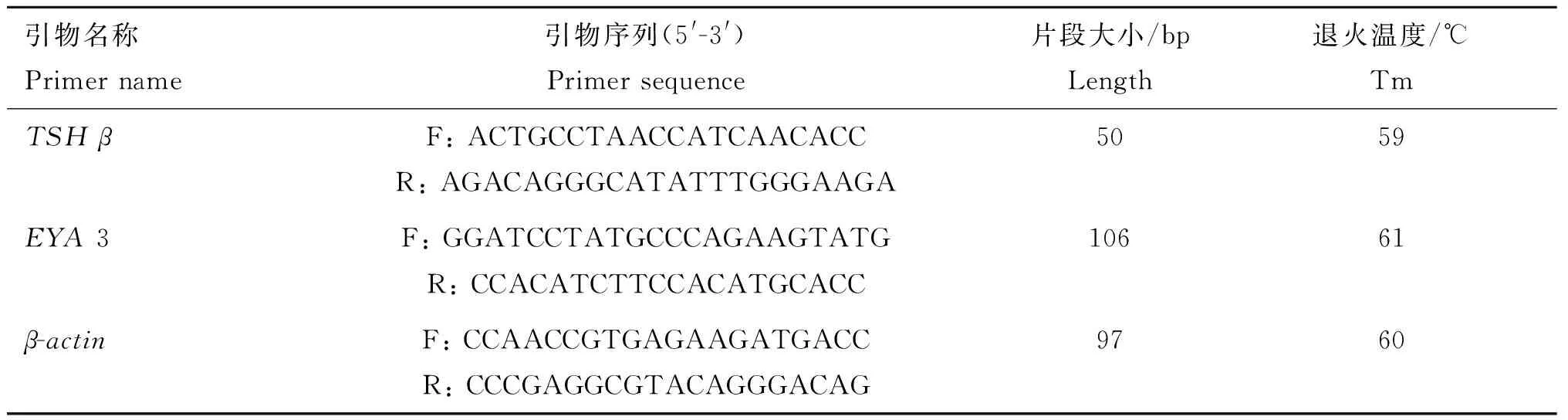
引物名称Primername引物序列(5'-3')Primersequence片段大小/bpLength退火温度/℃TmTSHβF:ACTGCCTAACCATCAACACCR:AGACAGGGCATATTTGGGAAGA5059EYA3F:GGATCCTATGCCCAGAAGTATGR:CCACATCTTCCACATGCACC10661β-actinF:CCAACCGTGAGAAGATGACCR:CCCGAGGCGTACAGGGACAG9760
1.2.2 反转录 使用PrimeScriptTMRT Reagent Kit反转录试剂盒合成cDNA第一链,按照说明书进行操作。反转录产物稀释后,用内参基因β-actin进行PCR检测,检测合格后,-20 ℃保存。
1.2.3 荧光定量PCR 用荧光定量PCR方法检测苏尼特羊与小尾寒羊不同组织及不同光照时间点苏尼特羊垂体中EYA3与TSHβ表达量,每个样品3个重复,以β-actin为内参基因,设置阴性对照。反应体系总体积为20 μL:SYBR Premix ExTaqⅡ 10 μL,RNase Free ddH2O 6.4 μL,上、下游引物(10 pmol·L-1)各0.8 μL,cDNA 2 μL。反应程序:95 ℃预变性5 s;95 ℃变性5 s,60 ℃ 30 s,40个循环;反应结束后,进行熔解曲线分析。以2-ΔΔCT法[29]计算目的基因相对表达量。
1.2.4 数据分析 使用SPSS13.0软件对数据进行统计分析。组间比较采用单因素方差分析(ANOVA)检验。P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1 EYA3在SP和LP条件下SNT中的表达谱
荧光定量分析发现,EYA3在苏尼特羊各组织广泛表达,且不同光照条件下,EYA3均在松果体、输卵管和小脑中高表达,其中松果体表达量最高,其次是输卵管,然后是小脑,其它组织表达量较低,暗示EYA3可能在绵羊松果体、输卵管和小脑组织中发挥作用;另外,LP下,EYA3在各组织表达量均高于SP,且在松果体、大脑和输卵管中的表达量显著高于其它组织(P<0.05,图1),暗示该基因在松果体、大脑和输卵管中的表达差异可能与季节性发情相关。

1~10.垂体、松果体、大脑、小脑、下丘脑、输卵管、子宫、卵巢、肾上腺和肾。下同。*.P<0.051-10.Pituitary,pineal gland,brain,cerebellum,hypothalamus,fallopian tube,uterus,ovary,adrenal gland and kindey.The same as below. *.P<0.05图1 EYA 3在LP和SP下苏尼特羊不同组织中的表达水平Fig.1 The expression levels of EYA 3 in different tissues of Sunite sheep under LP and SP conditions
2.2 EYA 3在黄体期和卵泡期STH中的表达谱
荧光定量分析发现,EYA3也在小尾寒羊各组织广泛表达,且不同繁殖时期EYA3均在垂体、松果体和小脑中高表达,且垂体和松果体中,黄体期高于卵泡期,但并无显著差异,其它组织表达量较低(图2)。这一结果表明:除了垂体,小尾寒羊中EYA3可能也在松果体和小脑中发挥作用。
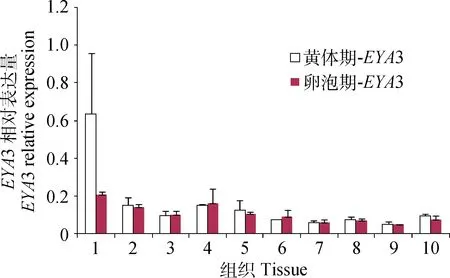
图2 EYA 3在小尾寒羊黄体期和卵泡期不同组织中的表达水平Fig.2 The expression levels of EYA 3 in different tissues of Small Tail Han sheep at luteal phase and follicular phase
2.3 TSH β 在LP和SP下,SNT中不同组织的表达
荧光定量分析发现,TSHβ在苏尼特羊垂体中高表达,其它组织几乎不表达,且LP下垂体中表达量极显著高于SP表达量(P<0.01)(图3)。
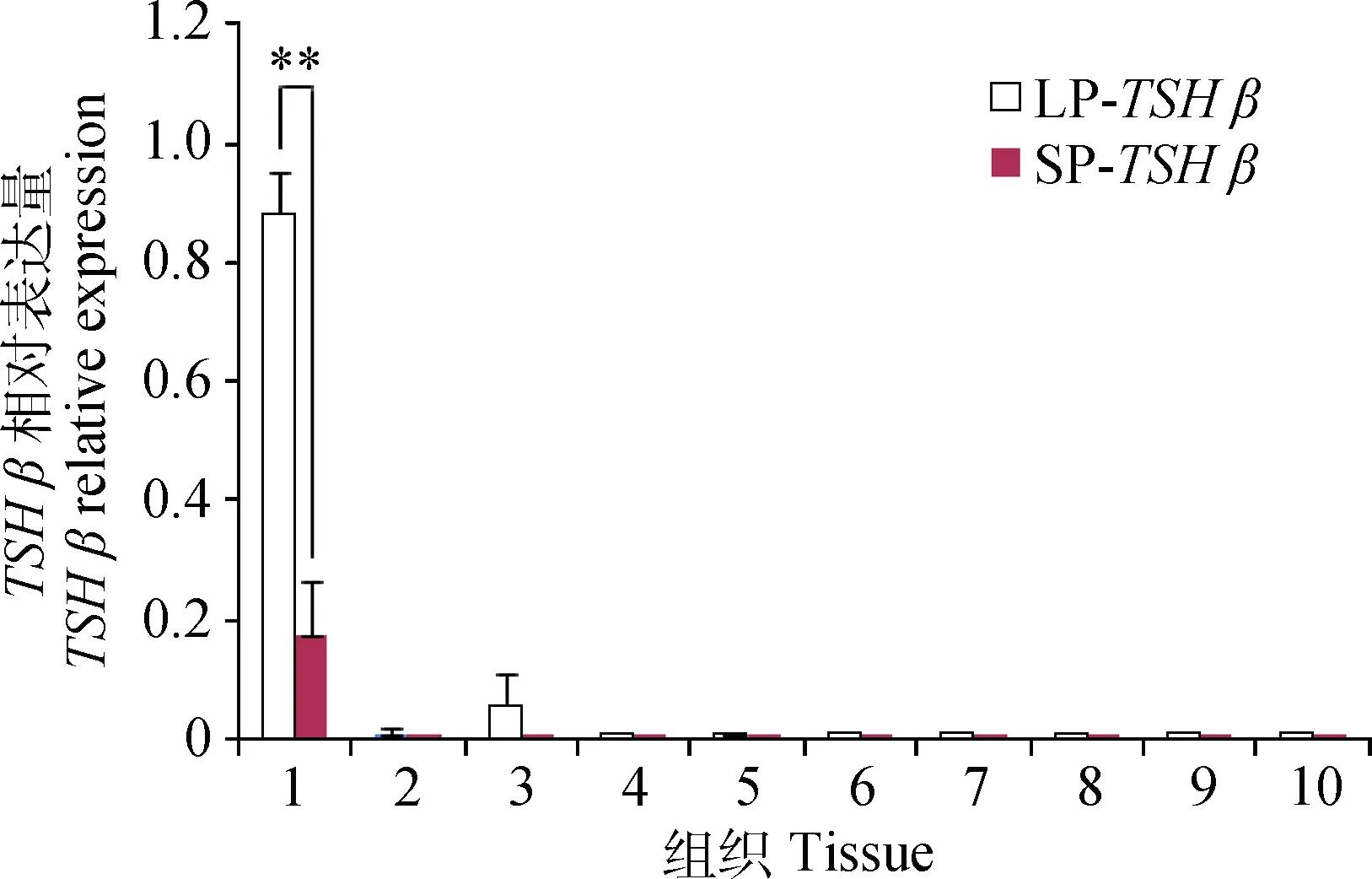
**.P<0.01。下同**.P<0.01.The same as below图3 TSH β 在LP和SP下苏尼特羊不同组织的表达水平Fig.3 The expression levels of TSH β in different tissues of Sunite sheep under LP and SP conditions
2.4 TSH β 在STH黄体期和卵泡期不同组织的表达
荧光定量分析发现,TSHβ在小尾寒羊垂体中高表达,其它组织几乎不表达,且黄体期垂体中表达量极显著高于卵泡期(P<0.01)(图4)。
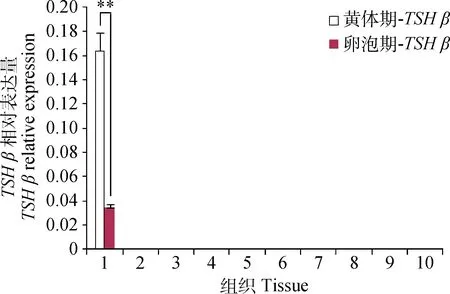
图4 TSH β 在小尾寒羊黄体期和卵泡期不同组织中的表达水平Fig.4 The expression level of TSH β in different tissues of Small Tail Han sheep at luteal phase and follicular phase
2.5 SP转变为LP后,SNT垂体中,EYA 3与TSH β的表达变化模式
荧光定量分析发现,不同时间点苏尼特羊垂体中EYA3与TSHβ均在SP转至LP后表达量升高,其中EYA3在LP 3达到表达高峰,之后逐渐降低;TSHβ在LP 21达到表达高峰,之后逐渐降低。EYA3表达量提前于TSHβ开始降低,表明EYA3发挥调控作用的时期早于TSHβ,主要在LP 3之前;而TSHβ主要在LP 21之前发挥调控作用。
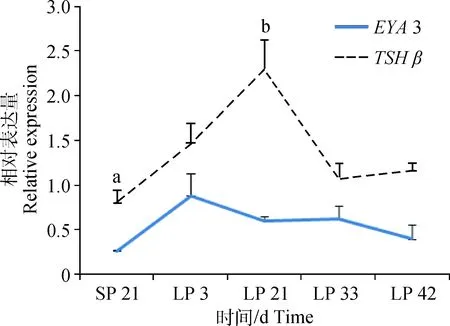
SP 21、LP 3、LP 21、LP 33、LP 42分别代表短光照第21天和长光照第3、21、33和42天SP 21, LP 3, LP 21, LP 33, LP 42 represent short photoperiod day 21, long photoperiod day 3, 21, 33 and 42图5 SP转变为LP后不同时间点苏尼特羊垂体中EYA 3与TSH β 的表达模式Fig.5 The expression pattern of EYA 3 and TSH β during the transition from SP to LP in pituitary of Sunite sheep
3 讨 论
3.1 常年发情与季节性发情绵羊不同时期EYA 3和TSH β的表达谱分析
S.H.WOOD等[30]以绵羊为研究对象发现,EYA3与TSHβ均可在PT区域的促甲状腺细胞中表达,两基因通过TSH β-DIO 2-TH通路激活或抑制下丘脑-垂体-性腺轴,达到调控绵羊季节性发情[31-32]。本研究荧光定量结果表明,EYA3与TSHβ在小尾寒羊和苏尼特羊各组织中的表达情况基本一致,即在两品种绵羊中EYA3均在多个组织中广泛表达,TSHβ主要在垂体中高表达,表明TSHβ主要在绵羊垂体中发挥作用。不同光照条件下,EYA3基因在苏尼特羊松果体、输卵管、小脑中高表达,在小尾寒羊中,不同繁殖时期EYA3在垂体、松果体和小脑中高表达,且黄体期表达量高于卵泡期;其中,黄体期EYA3在松果体、大脑和输卵管中表达量显著高于卵泡期。结合已知的季节性发情调控网络涉及的上游部位(松果体、垂体和下丘脑),上述结果暗示,EYA3可能也在松果体部位发挥作用并对绵羊季节性发情进行调控;另外,在绝大多数组织中EYA3和垂体中TSHβ表现为黄体期表达量高于卵泡期,暗示它们可能参与绵羊繁殖时期转换。
在苏尼特羊中,LP下TSHβ在垂体中表达量极显著高于SP;小尾寒羊中,黄体期TSHβ在垂体中表达量极显著高于卵泡期。季节性发情绵羊休情期PT区TSHβ高表达,导致下丘脑基底部GnRH神经元末梢被室管膜细胞包围,与PT细胞接触减少,末梢释放GnRH的量也相应降低[30,33]。据此,推测在小尾寒羊黄体期时,EYA3表达量升高可促进垂体中TSHβ高表达,进而通过TSH β-DIO 2-TH实现小尾寒羊下丘脑基底部GnRH神经元的作用,最终导致GnRH释放量降低;而卵泡期下丘脑基底部GnRH神经元末梢不被室管膜细胞包围,与PT细胞接触增多,导致GnRH释放量增加,促进FSH和LH的释放,从而影响卵泡发育和排卵,这一推测需进一步研究证实。
3.2 SP转至LP条件下,EYA 3与TSH β表达变化趋势
本试验通过对SP和LP不同时间点绵羊垂体中EYA3及TSHβ表达水平的检测,发现两基因表达趋势基本一致,均在SP转至LP后表达量升高。文献报道EYA3与TSHβ在绵羊昼夜节律调控中发挥重要作用[34-35]。K. H. Masumoto等[12]以雄性CBA/N小鼠为对象,通过PT区的全基因组表达谱分析发现了TSHβ等长光照诱导基因,并对TSHβ上游基因进行鉴定,发现EYA3对TSHβ有重要调控作用;在LP下,EYA3可与Sine Oculis-related Homeobox 1 (Six 1)、促甲状腺胚胎因子(Thyrotroph embryonic factor, Tef)及肝白血病因子(Hepatic leukemia factor, Hlf)共同组成复合体与TSHβ启动子结合促进TSHβ的表达[11,14,36]。N. Nakao等[3]在对鹌鹑的光周期信号转换通路研究中发现,LP 1关灯14 h后,TSHβ迅速表达,在接下来4 h后,DIO 2被快速诱导表达,并通过TSH β-DIO 2-TH通路最终影响垂体性腺轴激素的分泌[5-6]。鸟类中,在LP 1关灯后,上述两个基因高表达,但在LP 12以后表达量不再发生改变[15,37]。本研究中,苏尼特羊垂体的检测结果表明,EYA3在LP 3表达量最高,TSHβ在LP 21表达量最高,提示EYA3和TSHβ分别主要在LP 3和LP 21之前发挥关键作用,启动机体内长日照机制,使绵羊成功进入长日照繁殖模式。
4 结 论
本研究发现,LP下,EYA3在苏尼特羊松果体中表达量显著高于SP,在小尾寒羊黄体期表达量均高于卵泡期,在两品种绵羊中TSHβ均仅在垂体中高表达。结果表明,EYA3可能同时在松果体部位发挥作用并对绵羊季节性发情进行调控,另外,这两个基因可能参与繁殖时期的转换。在SP转至LP过程中,EYA3和TSHβ分别主要在LP 3和LP 21之前发挥关键作用,启动长日照繁殖模式。
[1] DARDENTE H. Circannual biology: the double life of the seasonal thyrotroph[J].CurrBiol, 2015, 25(20): R988-R991.
[2] WEEMS P W, GOODMAN R L, LEHMAN M N. Neural mechanisms controlling seasonal reproduction: principles derived from the sheep model and its comparison with hamsters[J].FrontNeuroendocrinol, 2015, 37: 43-51.
[3] NAKAO N, ONO H, YAMAMURA T, et al. Thyrotrophin in the pars tuberalis triggers photoperiodic response[J].Nature, 2008, 452(7185): 317-322.
[4] MAJUMDAR G, YADAV G, RANI S, et al. A photoperiodic molecular response in migratory redheaded bunting exposed to a single long day[J].GenCompEndocrinol, 2014, 204: 104-113.
[5] NAKANE Y, YOSHIMURA T. Universality and diversity in the signal transduction pathway that regulates seasonal reproduction in vertebrates[J].FrontNeurosci, 2014, 8: 115.
[6] CASSONE V M, YOSHIMURA T. Circannual cycles and photoperiodism[M]//SCANES C G. Sturkie’s Avian Physiology. 6th ed. London: Academic Press, 2015: 829-845.
[7] SURBHI, KUMAR V. Avian photoreceptors and their role in the regulation of daily and seasonal physiology[J].GenCompEndocrinol, 2015, 220: 13-22.
[8] KOZMIK Z, HOLLAND N D, KRESLOVA J, et al. Pax-Six-EYA-Dach network during amphioxus development: conservationinvitrobut context specificityinvivo[J].DevBiol, 2007, 306(1): 143-159.
[9] SALZER C L, ELIAS Y, KUMAR J P. The retinal determination gene eyes absent is regulated by the EGF receptor pathway throughout development inDrosophila[J].Genetics, 2010, 184(1): 185-197.
[10] CUNHA A F D, BRUGNEROTTO A F, DUARTE A S, et al. Global gene expression reveals a set of new genes involved in the modification of cells during erythroid differentiation[J].CellProlif, 2010, 43(3): 297-309.
[11] DARDENTE H, WYSE C A, BIRNIE M J, et al. A molecular switch for photoperiod responsiveness in mammals[J].CurrBiol, 2010, 20(24): 2193-2198.
[12] MASUMOTO K H, UKAI-TADENUMA M, KASUKAWA T, et al. Acute induction ofEYA3 by late-night light stimulation triggersTSHβ expression in photoperiodism[J].CurrBiol, 2010, 20(24): 2199-2206.
[13] HENNINGSEN J B, GAUER F, SIMONNEAUX V. RFRP neurons-the doorway to understanding seasonal reproduction in mammals[J].FrontEndocrinol(Lausanne), 2016, 7: 36.
[14] DUPRÉ S M, MIEDZINSKA K, DUVAL C V, et al. Identification ofEYA3 andTAC1 as long-day signals in the sheep pituitary[J].CurrBiol, 2010, 20(9): 829-835.
[15] MISHRA I, BHARDWAJ S K, MALIK S, et al. Concurrent hypothalamic gene expression under acute and chronic long days: implications for initiation and maintenance of photoperiodic response in migratory songbirds[J].MolCellEndocrinol, 2016, 439: 81-94.
[16] LIAO T H, PIERCE J G. The primary structure of bovine thyrotropin. II. The amino acid sequences of the reduced,S-carboxymethyl α and β chains[J].JBiolChem, 1971, 246(4): 850-865.
[17] BOCKMANN J, BÖCKERS T M, VENNEMANN B, et al. Short photoperiod-dependent down-regulation of thyrotropin-alpha and-beta in hamster pars tuberalis-specific cells is prevented by pinealectomy[J].Endocrinology, 1996, 137(5): 1804-1813.
[18] 赵婉秋, 陈 黎, 沈军达, 等. 动物季节性繁殖机制研究进展[J]. 浙江农业科学, 2017, 58(1): 150-154. ZHAO W Q, CHEN L, SHEN J D, et al. Research progress in seasonal reproduction mechanism of animal[J].JournalofZhejiangAgriculturalSciences, 2017, 58(1): 150-154. (in Chinese)
[19] 黄冬维, 储明星. 动物季节性繁殖分子调控机理研究进展[J]. 遗传, 2011, 33(7): 695-706. HUANG D W, CHU M X. Research progress in molecular mechanism of animal seasonal reproduction[J].Hereditas(Beijing), 2011, 33(7): 695-706. (in Chinese)
[20] SHINOMIYA A, SHIMMURA T, NISHIWAKI-OHKAWA T, et al. Regulation of seasonal reproduction by hypothalamic activation of thyroid hormone[J].FrontEndocrinol, 2014, 5: 12.
[21] 杨 颖, 陈 黎, 卢立志. 松果体调控动物季节性繁殖概述[J]. 农业生物技术学报, 2017, 25(7): 1086-1101. YANG Y, CHEN L, LU L Z. Review of regulation on the pineal gland of animal seasonal reproduction[J].JournalofAgriculturalBiotechnology, 2017, 25(7): 1086-1101. (in Chinese)
[22] WOOD S, LOUDON A. Clocks for all seasons: unwinding the roles and mechanisms of circadian and interval timers in the hypothalamus and pituitary[J].JEndocrinol, 2014, 222(2): R39-R59.
[23] YASUO S, YOSHIMURA T, EBIHARA S, et al. Photoperiodic control ofTSH-βexpression in the mammalian pars tuberalis has different impacts on the induction and suppression of the hypothalamo-hypopysial gonadal axis[J].JNeuroendocrinol, 2010, 22(1): 43-50.
[24] AIZAWA S, HOSHINO S, SAKATA I, et al. Diurnal change of thyroid-stimulating hormone mRNA expression in the rat pars tuberalis[J].JNeuroendocrinol, 2007, 19(11): 839-846.
[25] GOULART-SILVA F, DE SOUZA P B, NUNES M T. T3 rapidly modulates TSHβ mRNA stability and translational rate in the pituitary of hypothyroid rats[J].MolCellEndocrinol, 2011, 332(1-2): 277-282.
[26] 刘 浩, 臧晓怡, 刘春蓉, 等. 促甲状腺激素β基因剪接变体在BALB C小鼠不同组织中的表达[J]. 天津医药, 2011, 39(3): 239-242. LIU H, ZANG X Y, LIU C R, et al. A novel thyroid stimulating hormone β-subunit splice variant in bone marrow and other tissues in BALB C mice[J].TianjinMedicalJournal, 2011, 39(3): 239-242. (in Chinese)
[27] 黄冬维, 曹贵玲, 储明星, 等. 山羊促甲状腺素β亚基基因(TSHB)cDNA克隆与组织表达研究[J]. 安徽农业大学学报, 2012, 39(6): 847-853. HUANG D W, CAO G L, CHU M X, et al. cDNA cloning and tissue expression of thyroid stimulating hormone beta chain gene (TSHB) in goats[J].JournalofAnhuiAgriculturalUniversity, 2012, 39(6): 847-853. (in Chinese)
[28] TSUJINO K, NARUMI R, MASUMOTO K H, et al. Establishment ofTSHβ real-time monitoring system in mammalian photoperiodism[J].GenesCells, 2013, 18(7): 575-588.
[29] SCHMITTGEN T D, LIVAK K J. Analyzing real-time PCR data by the comparativeCTmethod[J].NatProtoc, 2008, 3(6): 1101-1108.
[30] WOOD S H, CHRISTIAN H C, MIEDZINSKA K, et al. Binary switching of calendar cells in the pituitary defines the phase of the circannual cycle in mammals[J].CurrBiol, 2015, 25(20): 2651-2662.
[31] YOSHIMURA T. Thyroid hormone and seasonal regulation of reproduction[J].FrontNeuroendocrinol, 2013, 34(3): 157-166.
[32] DARDENTE H, HAZLERIGG D G, EBLING F J P. Thyroid hormone and seasonal rhythmicity[J].FrontEndocrinol, 2014, 5: 19.
[33] KORF H W. Signaling pathways to and from the hypophysial pars tuberalis, an important center for the control of seasonal rhythms[J].GenCompEndocrinol, 2017, doi: 10.1016/j.ygcen.2017.05.011.
[34] IKEGAMI K, YOSHIMURA T. Comparative analysis reveals the underlying mechanism of vertebrate seasonal reproduction[J].GenCompEndocrinol, 2016, 227: 64-68.
[35] NISHIWAKI-OHKAWA T, YOSHIMURA T. Molecular basis for regulating seasonal reproduction in vertebrates[J].JEndocrinol, 2016, 229(3): R117-R127.
[36] DARDENTE H, LOMET D, ROBERT V, et al. Seasonal breeding in mammals: from basic science to applications and back[J].Theriogenology, 2016, 86(1): 324-332.
[37] MISHRA I, SINGH D, KUMAR V. Seasonal alterations in the daily rhythms in hypothalamic expression of genes involved in the photoperiodic transduction and neurosteroid-dependent processes in migratory blackheaded buntings[J].JNeuroendocrinol, 2017, 29(5): 12469.
