黄色潜蝇茧蜂的寄生功能反应研究
2018-03-12曹凤勤崔志富程立生
杨 琪, 曹凤勤, 崔志富, 程立生
(1.海南大学热带农林学院,海南海口 570228; 2.海南省农垦农业科学研究所,海南海口 570200; 3.琼台师范学院,海南海口 571127)
美洲斑潜蝇(LiriamyzasativaeBlanchard)主要危害葫芦科、茄科、十字花科、菊科等植物叶片及果实,我国最早于1993年在海南省三亚市发现美洲斑潜蝇,现已蔓延至全国21个省(市)[1-2]。20 世纪50 年代前,由于寄生性天敌种类丰富、自然控制作用强,美洲斑潜蝇曾被认为是一类不会导致重大经济损失的小型害虫。过去防治美洲斑潜蝇主要采用化学防治,但由于农药的频用和滥用,其自然天敌受到抑制,加之其寄主范围广、世代历期短、繁殖力高、危害隐蔽、易产生抗药性等原因,潜叶蝇类害虫已成功入侵并扩散至世界多个国家并持续危害作物,同时也给我国的蔬菜及花卉等产业造成了巨大的经济损失,成为我国农业生产上最突出的问题之一[1]。由此开始,我国学者开始了对美洲斑潜蝇寄生性天敌的调查和研究,大量学者对美洲斑潜蝇寄生蜂的生物学及生态学特性进行了研究讨论[1-5]。寄生蜂在生物防治中所起到的作用得到充分的认可[6-8]。
经过对海南岛天敌资源害虫的调查,发现黄色潜蝇茧蜂[OpiusflavusWeng et Chen (Hymenoptera:Braconidae)]是海南省的优势种之一,对斑潜蝇等潜叶蝇有较强的控制作用,然而国外尚未见对其寄生作用的相关研究,国内对其报道也甚少。对其成虫功能反应进行研究,能了解该蜂的生活习性,并从中找到该蜂的最佳繁殖条件,为更好地利用该天敌防治美洲斑潜蝇提供理论依据。
功能反应是指每个捕食者的捕食率随猎物密度变化的一种反应,即捕食者对猎物的捕食效应[9]。对天敌功能反应进行研究,对评价其防治效果具有重要意义。后来研究发现功能反应也能用来描述寄生蜂的寄生能力、种群动态与寄主密度之间的关系[9]。寄生蜂的寄主选择行为一般包括寄主栖境定位、寄主定位、寄主接受和寄主适应性,在此过程中,寄生蜂识别寄主与非寄主,并判断不同寄主栖境与不同寄主间的收益性[10]。
本试验在实验室条件下,以美洲斑潜蝇幼虫为寄主,研究了不同温度下寄主密度及寄生蜂自身密度对黄色潜蝇茧蜂寄生行为的影响,旨在为合理有效地利用黄色潜蝇茧蜂提供一定的指导意义。
1 材料与方法
1.1 供试虫源及饲养
美洲斑潜蝇采自海南大学海甸校区实验基地及校园周边菜地,在海南大学入侵害虫实验室进行室内鉴定并利用菜豆进行长期连代饲养。
黄色潜蝇茧蜂采自海南大学海甸校区实验基地及校园周边、琼海等地。在海南大学入侵害虫实验室的温室(26 ℃,湿度50%~70%,光照度3 000 lx)内,用2龄末至3龄初的美洲斑潜蝇幼虫长期继代饲养。
1.2 主要试剂
葡萄糖(广东广州化学试剂厂)、琼脂粉(北京索莱宝科技有限公司)。
1.3 主要仪器
V-LH100HG Olympus显微镜、PRX-450D智能人工气候箱(浙江宁波赛福实验仪器厂)。
1.4 试验方法
试验在人工气候箱内进行。各处理取带有美洲斑潜蝇3龄幼虫的菜豆叶片,在直径为12.5 cm的培养皿内倒入一定量琼脂,后贴入上述叶片,对其进行固定、保湿,引入寄生蜂成蜂,将蘸有20%葡萄糖溶液的棉球置于培养皿内壁,为寄生蜂补充营养,最后用保鲜膜覆盖培养皿,并对保鲜膜进行扎孔处理。
1.4.1 不同温度下黄色潜蝇茧蜂对美洲斑潜蝇的寄生功能反应 美洲斑潜蝇的虫口密度分别设置为30、40、50、60、70头/皿,同时接入羽化48 h且已交配的黄色潜蝇茧蜂雌蜂1头,分别置于22、26、30、34 ℃(光照时间 ∶黑暗时间=14 ∶10,湿度70%~80%)的恒温气候箱中,寄生24 h后移除寄生蜂,随即在显微镜下解剖供试的美洲斑潜蝇幼虫,观察寄生情况。每个处理重复20次。
统计方法:HollingⅡ方程模型为Na(1+aThN)=aTN。式中,Na为被寄生的寄主量(头/皿);N为寄主密度(头/皿);T为总的寄生时间(d);a为瞬间攻击率;Th为寄生蜂处理每头寄主所耗时间(d)。
1.4.2 不同温度下黄色潜蝇茧蜂对美洲斑潜蝇的搜寻效应 试验方法同“1.4.1”。统计方法:S=a/(1+aThN)。式中,S为搜寻效应。
1.4.3 不同温度下黄色潜蝇茧蜂的自身密度干扰效应 分别将2、4、6、8、10头黄色潜蝇茧蜂雌成蜂引入含60头美洲斑潜蝇3龄幼虫的培养皿中,寄生24 h后移除寄生蜂,解剖供试的美洲斑潜蝇幼虫,观察并记录寄生情况。每处理重复20次。
统计方法:Hassell-Varley干扰模型为E=QP-m,其中E为寄生作用率,Q为搜寻常数,P为寄生蜂密度,m为相互干扰系数。
所有数据分析及绘图使用SAS 9.1及Excel软件进行。
2 结果与分析
2.1 不同温度下黄色潜蝇茧蜂对美洲斑潜蝇的寄生功能反应
从表1可以看出,同一温度下,黄色潜蝇茧蜂对美洲斑潜蝇的寄生量与寄主密度成正比,当寄主密度为30头/皿时,日寄生量最少,当寄主密度上升至70头/皿时寄生量达到最大,特别是22 ℃时,寄主密度为70头/皿时的寄生量约是 30头/皿 时的2倍。在相同密度条件下,26 ℃是该寄生蜂寄生量最大的温度,而34 ℃的寄生量最低。由表1还可以看出,温度相同时,寄生率与美洲斑潜蝇密度成反比;而在相同寄主密度条件下,黄色潜蝇茧蜂对美洲斑潜蝇的寄生率先随温度升高而增加,在26 ℃时寄生率达到峰值之后又下降,且34 ℃时的寄生率是4个温度中最低的。
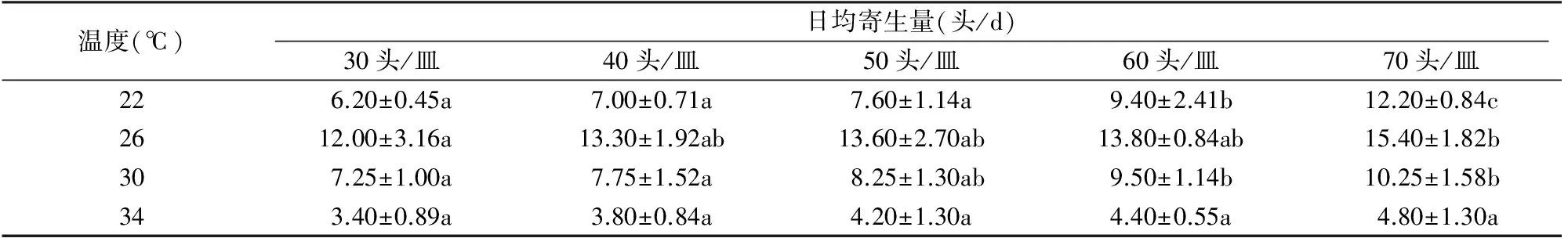
表1 不同温度下黄色潜蝇茧蜂的日均寄生量
注:表中数据为平均值±标准误,同行字母不同表示处理间差异显著(P<0.05,LSD法)。
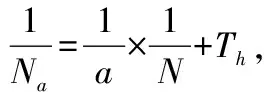
通过对功能方程的模拟,反映出寄生蜂在不同温度下的瞬时攻击率及对寄主的处理时间存在差异。瞬时攻击率由大到小依次为26 ℃>30 ℃>22 ℃>34 ℃。寄生蜂在26 ℃时表现出的瞬时攻击率最强,可达0.80,而34 ℃的瞬时攻击率最低,仅为0.23;对寄主的处理时间随温度升高而延长,34 ℃时处理时间最长,达到0.15 d。参考陈俊谕的评判方法[11],以寄生能效判断寄生能力,由表2可看出,26 ℃时黄色潜蝇茧蜂寄生能力最强,是34 ℃时寄生能效的10倍多。由此可推测26 ℃是较适宜寄生蜂活动的温度,而高温对寄生蜂的寄生能力有较大影响。
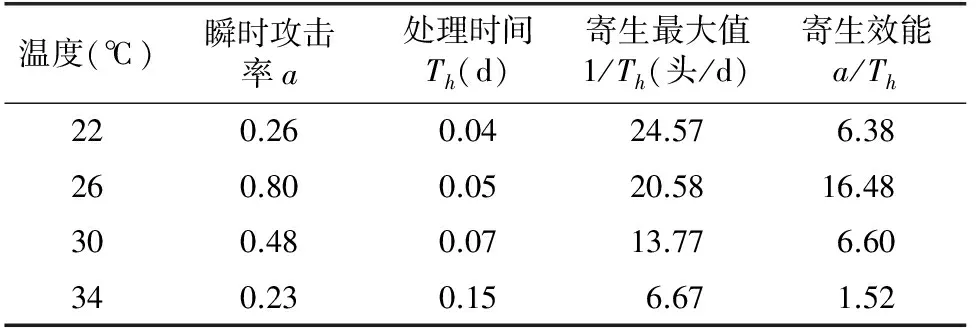
表2 黄色潜蝇茧蜂对美洲斑潜蝇HollingⅡ型功能反应参数估计
2.2 不同温度下黄色潜蝇茧蜂对美洲斑潜蝇的搜寻效应
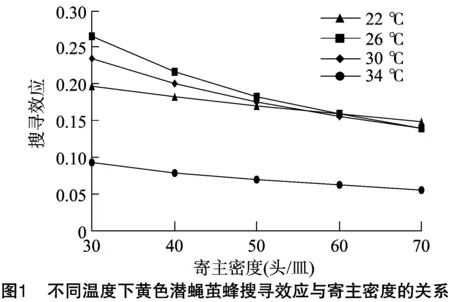
以(N)为横坐标,(S)为纵坐标得到图1。搜寻效应与寄主密度之间的关系分别为S22 ℃=-0.011 9x+0.207 4,r2=0.993 3;S26 ℃=-0.030 8x+0.285 2,r2=0.965 5;S30 ℃=-0.023 5x+0.251 5,r2=0.976 8;S34 ℃=-0.008 9x+ 0.099,r2=0.978 9。同一温度下,搜寻效应与寄主密度成反比,表明越大的寄主密度会对寄生蜂产生更大的干扰作用。同一寄主密度下,温度同样影响着寄生蜂搜寻效应,寄主密度为30~60头/皿时,26 ℃搜寻效应最高;而寄主密度为70头/皿 时,22 ℃的搜寻效应最高,但无论何种寄主密度,寄生蜂在34 ℃条件下的搜寻效应最低。
2.3 不同温度下黄色潜蝇茧蜂的自身密度干扰效应
相同温度条件下,寄生蜂对寄主的寄生作用率随自身密度的增加而呈现负增长趋势;相同寄生蜂密度条件下,温度升高寄生作用率也增大,26 ℃时出现峰值,30 ℃时寄生作用率又降低,34 ℃时寄生作用率降至最低谷。通过Hassell-Varley模型对黄色潜蝇茧蜂的自身密度干扰效应模拟后发现,黄色潜蝇茧蜂自身密度的干扰能较好地拟合该模型。26 ℃ 时的搜寻常数最大,干扰系数最小;34 ℃的搜寻常数最小,干扰系数最大(表3、表4)。说明高温时寄生蜂的搜寻能力较弱,而适当的温度可提高寄生蜂的搜索能力。
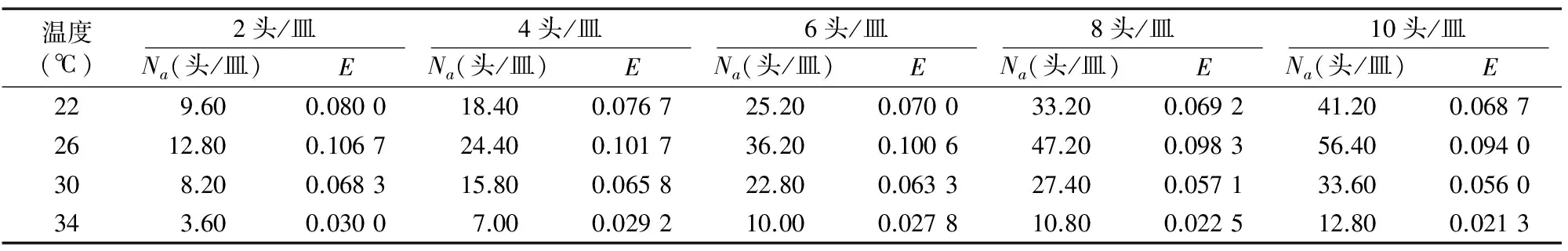
表3 不同温度下黄色潜蝇茧蜂的自身密度干扰效应
3 讨论
Holling提出的功能反应模型是研究天敌对其作用对象作用大小的方法[12],尤其在有害生物的生物防治中有较多的应用,它是天敌对害虫是否有作用的评估标准[13]。功能反应Ⅱ是寄生蜂的主要模式,当前我国学者对寄生蜂功能反应的研究基本上都是进行的实验室实验,呈现出的结果均符合模型Ⅱ[4-5,14-17]。本试验结果也表明,4个温度下,HollingⅡ功能反应模型均适合于模拟黄色潜蝇茧蜂对不同寄主密度的寄生作用。
HollingⅡ模型能够预测瞬时攻击率及作用时间这2个反应参数。本研究结果表明,黄色潜蝇茧蜂的寄生量会跟随寄主密度增加,但寄主密度上升到某个阶段后,被寄生的斑潜蝇数量增幅减缓。原因可能有以下几点:(1)由于寄生蜂对寄主幼虫活动的抗干扰能力较弱,或遗传因素导致的寄生蜂生殖力的影响;(2)天敌在寄生寄主后,通常会用化学物质对寄主或者寄主所在的生境进行标记,这会对后继的寄生蜂产生影响;(3)同种黄色潜蝇茧蜂的个体,其各自的遗产背景、经历及生理状况存在差异。不同学者分别对斑痣悬茧蜂、蝇蛹金小蜂、浅黄蚜恩小蜂、丽蚜小蜂、螟蛉盘绒茧蜂等的功能反应研究也得出以上结论[4-5,14-17]。此外,在温度的影响下,寄生蜂的寄生作用也发生了较大变化,尤其是高温会加速寄生蜂体内物质代谢,从而对寄生蜂的活动能力、体内成熟卵子的形成及寄生行为都有较大影响。本研究发现,26 ℃对黄色潜蝇茧蜂的干扰效应最小。陈俊谕等也发现温度对哥德恩蚜小蜂的干扰效应较大[11];章玉苹等研究后认为,蝇蛹佣小蜂的最佳存活温度为26 ℃[4]。
在野外环境下,无法控制恒定的温湿度,这些都是影响寄生蜂生长发育、觅食及繁殖的重要因素,甚至会使寄生蜂对寄主的功能反应类型发生改变[18]。因此在进行寄生蜂的田间释放时,一定要注意选择合适的放蜂季节及天气。
搜寻效应长与寄主密度及寄生蜂自身密度有密切联系[19]。随着寄生蜂密度增加,个体间出现相互干扰与竞争[16];尤其在活动空间及寄主密度一定时,寄生蜂密度与干扰效应成正比,与寄生作用率成反比。这种现象在许多寄生蜂的室内寄生试验中都存在[4,11,17]。因此也说明,在进行室内人工繁蜂以及室外寄生蜂释放时,都应充分考虑寄生蜂个体之间的相互干扰对其寄生作用力的影响,特别是室外放蜂时,并非寄生蜂释放得越多越好;一定要做好释放田的调查工作,了解目标害虫的虫口密度、发生情况,以确定最适放蜂量,提高寄生蜂的利用率,节省人力、财力。
本试验在实验室条件下进行,光照、温度、湿度等因素都较单一,与野外大容积、高复杂程度的环境空间有较大差异,但基本模拟了繁殖黄色潜蝇茧蜂的条件,给人工大量繁蜂奠定了理论基础。试验时使用的美洲斑潜蝇寄主龄期为2龄末3龄初,但田间条件下美洲斑潜蝇幼虫龄期多样[20]。因此在田间释放时还需考虑田间的温湿度、光照强度、光周期、寄主龄期以及寄生蜂日龄等综合效应对黄色潜蝇茧蜂寄生效能的影响。
[1]许再福,曾 玲. 美洲斑潜蝇寄生蜂研究概况[J]. 昆虫天敌,1998,20(3):129-135.
[2]蔡笃程. 海南寄生性天敌对美洲斑潜蝇自然种群控制作用的生态基础研究[D]. 儋州:华南热带农业大学,2003:3-4.
[3]吴佳教,张维球. 一种值得重视的蔬菜害虫——瓜斑潜蝇[J]. 昆虫知识,1997,34(4):216-218.
[4]章玉苹,李敦松,张宝鑫,等. 蝇蛹俑小蜂对橘小实蝇蛹的功能反应及温湿度对蜂成虫寿命的影响[J]. 中国生物防治,2010,26(4):385-390.
[5]尚 禹. 寄生蜂功能反应研究进展[J]. 江苏农业科学,2014,42(5):130-132.
[6]陈学新. 21世纪我国害虫生物防治研究的进展、问题与展望[J]. 昆虫知识,2010,47(4):615-625.
[7]Johnson M W,Oatman E R,Wyman J A. Natural control ofLiriomyzasativae(Diptera:Agromyzidae) in pole tomatoes in southern California[J]. Entomophaga,1980,25(2):193-198.
[8]Lorini I,Foerster L A. Seasonal fluctuation and parasitism ofLiriomyzasativaeBlanchard,1938 (Diptera:Agromyzidae) on cucumber (CucumissativusL.)[J]. Anais da Sociedade Entomologica do Brasil (Brazil),1985,14(2):243-249.
[9]Houck M A,Strauss R E. The comparative study of functional responses:experimental design and statistical interpretation[J]. The Canadian Entomologist,1985,117(5):617-629.
[10]戈 峰. 昆虫生态学原理与方法[M]. 北京:高等教育出版社,2008:198.
[11]陈俊谕,陈泰运,符悦冠,等. 哥德恩蚜小蜂对螺旋粉虱的功能反应研究[J]. 中国生物防治学报,2013,29(2):175-180.
[12]Holling C S. Some charaeteristics of simple types of predation and parasitism[J]. Canadian entomologist,1959,91(7):385-398.
[13]Jones D B,Giles K L,Berberet R C,et al. Functional responses of an introduced parasitiod and an indigenous parasitiod on greenbug at four temperature[J]. Environmental Entomology,2003,32(3):425-432.
[14]赵海燕,曾 玲,梁广文,等. 蝇蛹金小蜂对橘小实蝇蛹的寄生功能反应[J]. 环境昆虫学报,2014,36(1):122-126.
[15]王 甦,赵 静,张 帆,等. 利用功能反应模型评价两种寄生蜂混合释放对烟粉虱的防控潜能[J]. 环境昆虫学报,2014,36(2):188-193.
[16]陈青召. 丽蚜小蜂寄生蓖麻粉虱的生物学及其功能反应[D]. 杨凌:西北农林科技大学,2015:28-30.
[17]江化琴. 螟蛉盘绒茧蜂生物学特性及其对稻纵卷叶螟控制能力的研究[D]. 重庆:西南大学,2015:32-36.
[18]Mohaghegh J,De Clercq P,Tirry L. Functional response of the predatorsPodisusmaculiventris(Say) andPodisusnigrispinus(Dallas) (Het.,Pentatomidae) to the best armyworm,Spodopteraexigua(Hubner) (Lep.,Noctuidae):effect of temperature[J]. Appl Entomol,2001,125(3):131-134.
[19]巫厚长,程遐年,邹运鼎. 不同饥饿程度的龟纹瓢虫对烟蚜的捕食作用[J]. 应用生态学报,2000,11(5):749-752.
[20]林进添,凌远方. 几种有机磷类杀虫药剂对美洲斑潜蝇活性的研究[J]. 中国蔬菜,1998(2):6-10.
