一种基于海马认知机理的仿生机器人认知地图构建方法
2018-03-10于乃功苑云鹤蒋晓军罗子维
于乃功 苑云鹤 李 倜 蒋晓军 罗子维
对空间环境进行有效的认知是动物赖以生存的重要能力.Tolman的三迷宫导航实验揭示动物的导航行为依靠被称为认知地图的内在环境表达,这种表达反映了环境中显著路标之间的相互联系[1].
损毁实验研究表明,海马结构是环境认知和情节性记忆产生的关键脑区[2],损毁了海马体[3]、内嗅皮层[4]或后下托[5]的老鼠均不能完成Morris水迷宫导航任务.对海马结构不同区域的细胞进行放电记录表明海马结构中存在着一系列与空间环境认知相关的细胞.存在于CA区(海马角,海马区的一部分)的位置细胞只在空间特定位置放电,是构成认知地图的基础机制[6−8];后下托中的头朝向细胞只在以我为中心特定的头方向上放电,是对动物头朝向的表征,而位于丘脑核背侧的头朝向细胞的放电率还受到当前头部的瞬时转速的影响[9−10].
先前的研究模型[11]仅采用头朝向细胞和位置细胞构建任意尺度空间关系的认知地图,这种对单个头朝向细胞和位置细胞活动的离散表达,不足以支持从一个位置到另一个位置的导航行为.行为学研究表明,动物能够利用自运动线索保持相对于环境重要位置(例如巢穴)的矢量关系,并将空间中的各个位点的信息进行计算和整合,使这种矢量关系不断得到更新,这一过程被称为路径积分[12].内嗅皮层浅皮层中的网格细胞[13−14]被认为是大脑内部的路径积分器[2].
网格细胞在空间六边形区域周期性放电,其放电野随着动物探索环境而覆盖整个环境.无外源性输入、移除或者替换主要路标点后网格野也能够持续存在[13].自运动线索是网格细胞的主要输入,用于保持或更新网格野.网格细胞通过对自运动信号进行路径积分更新位置细胞的放电野[2].因此,网格细胞为认知地图的形成提供了空间度量[15].
几乎所有解释内嗅皮层网格细胞形成机制的模型都是利用速度调节的输入信号的路径积分表征周期性的网格野,现阶段网格细胞的主要计算模型有振荡干涉模型[16−19]和吸引子模型[20−21].2012年O′Keefe和Burgess在Science上发表文章说明,在傍下托和内嗅皮层的浅皮层发现了在平面上有周期性条纹状放电野的细胞族,不同的细胞族有着不同的放电朝向和波长,这些细胞被称为条纹细胞(Band cell).
作为位置细胞的主要输入信号,内嗅皮层网格细胞对空间的表达是对海马位置细胞空间地图的补充,网格细胞族是对空间环境度量信息的表达,与环境中的路标信息无关,而位置细胞地图则是对空间特定位置点的表达,内嗅海马回路对环境的表达构成了环境的认知地图[15].周期性放电的网格细胞如何生成空间特异性放电的位置细胞仍然是一个开放性的问题.现阶段存在两种计算方式,1)采用在特定频率范围具有相同相位和不同空间尺度的网格细胞子集线性相加产生位置细胞[2,14,20];2)采用具有多个空间尺度和朝向的网格细胞竞争学习实现[22−24].外源性信息,例如视觉和嗅觉信号同样能够使位置细胞放电,老鼠在发现空间显著路标时,能够以此作为参考纠正路径积分误差.
已有多个模型利用海马空间细胞的路径积分实现了智能体在环境中的定位,虽然采用强化学习和外界视觉信号能够实现面向目标导航和强化定位功能的目的,但这些模型并未构建出环境的认知地图[19,25−28],多数利用振荡干涉的计算模型还只停留在仿真阶段,并未实现在真实环境中有效的地图构建.澳大利亚昆士兰理工大学的Milford利用神经行为学研究成果,对老鼠大脑假设的“位姿细胞”进行数学建模,利用视觉驱动的导航系统,建立了一个可以实时定位与地图构建的RatSLAM仿生导航算法[29−32],在大规模的室外导航实验中,取得了较好的效果.但RatSLAM算法是一种纯视觉的导航算法,其核心传感信息来源于视觉里程计,受外界环境的影响较大,存在可靠性低、环境适应性较差的问题[33].
本文利用吸引子模型构建与海马认知机理有关的空间细胞计算模型.利用网格细胞对自运动线索进行路径积分,通过竞争性Hebb学习选择相应网格子集生成位置细胞族活动,位置细胞之间的竞争学习产生单峰型放电野,以二维环状吸引子模型表达空间环境的实际位置,位置细胞的路径积分由条纹细胞驱动,实现在仿真和真实环境中的精确路径积分.与此同时,利用RGB-D传感器(3D摄像机可采集颜色深度图)采集视觉线索进行环境的闭环检测,最终生成的认知地图是拓扑度量地图,包含环境特征点坐标、视觉线索以及特定位点的拓扑关系.
本文在以下两个方面取得了进步:
1)利用统一的计算机制模拟了海马结构中四种空间细胞的计算模型,在已知的研究中,本文是第一个采用条纹细胞驱动实现真实环境路径积分功能的模型.网格细胞模型的路径积分能够在持续260m的空间范围中,误差不超过15cm,体现了模型的精确度.
2)利用实际物理平台实现了环境认知地图构建,相比于传统的SLAM导航方法,对硬件和传感器要求低,使用仿生的方式使整个模型有良好的扩展性和适应性,适用于不同室内环境的导航[34−37].
1 基于海马空间细胞的认知地图构建方法
本节对构建认知地图模型的整体结构(如图1所示)和所用方法(如图2所示)进行详细说明. 1)对四种空间细胞,即头朝向细胞、条纹细胞、网格细胞和位置细胞的生理学特征、功能以及计算模型进行说明.2)详细阐述基于空间细胞认知机理的位置细胞地图的构建方法和基于颜色深度图像进行闭环检测的方法.3)展示如何基于视觉线索、自运动信号以及空间细胞认知机理构建精确的环境认知地图.
1.1 头朝向细胞
1.1.1 头朝向细胞的生理学研究基础
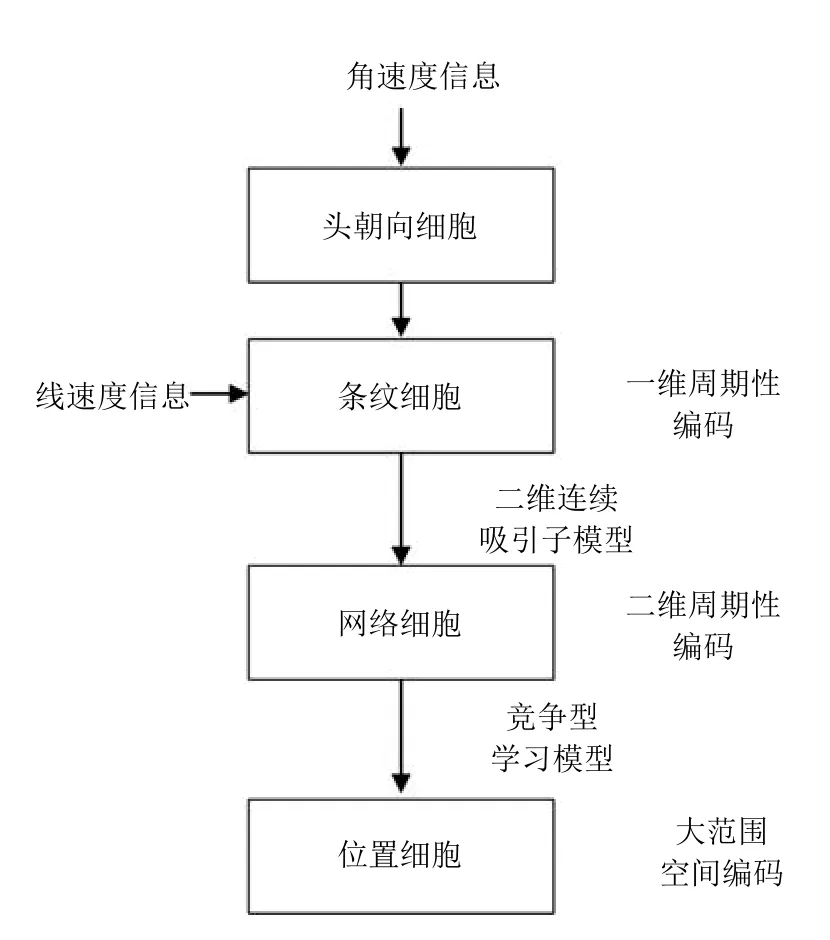
图2 空间细胞路径积分信息传递图Fig.2 Space cell path integral information transfer route
动物如何对它的运动方向进行编码?这个问题随着Ranck等于1984年在老鼠前下托发现了头朝向细胞而得到解决[38−39].头朝向细胞对于指导动物的运动有重要作用,当老鼠头部面向一个特定方向时,头朝向细胞发生最大放电,当头偏离这个方向时,细胞放电会逐渐减弱,其放电情况如图3所示,该头朝向细胞的优先方向是150°,那么以水平角为起点,其放电强度随着老鼠头朝向角度的增大而增大,当老鼠的头朝向到达该头朝向细胞的最优方向(Preferred direction)150°时,电信号强度达到峰值,之后其放电强度随着头朝向角度的增大逐渐偏离最优方向而逐渐减小,这个过程可由一个高斯函数近似表达.位于后下托的头朝向细胞只受当前瞬时头朝向的影响,而位于丘脑核背侧的头朝向细胞还受到当前头部的瞬时转速的影响.
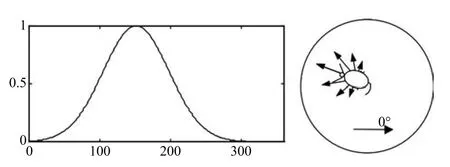
图3 头朝向细胞的放电情况Fig.3 The fi ring of head direction cells
后下托中的头朝向细胞的最优方向是固定的,而且有且只有一个最优方向,不受环境、老鼠身体方向等因素的影响,只与当前的头朝向有关,而丘脑核背侧的头朝向细胞随着头部的转动,其最优方向也有所改变,与老鼠未来的头朝向有关,其放电活动可能是根据头转动方向(顺时针+逆时针)以及当时的转动速度计算出未来的头朝向,这对于指导老鼠的运动有重要作用[40].不论老鼠是静止的或者运动的,头朝向细胞始终是激活的.与位置细胞类似,头朝向细胞的联合响应完成了老鼠对水平方向的编码,当所有头朝向细胞的信息整合到一起时就产生了一个连续的老鼠头朝向信号.先前的实验结果表明,所有的头朝向细胞的整合方向倾向于锁定于一个特定的主要朝向[41−42].
1.1.2 头朝向细胞计算模型
基于以上的生理学研究事实,本文使用头朝向细胞产生一个角速度调节信号,即放电率与当前头朝向和老鼠运动的速度成比例.第i个头朝向细胞的优先方向可以表示为一个相对于主要朝向θ0的角偏移量θi,那么给定头朝向细胞的调整内核:

通过编码器获取机器人的瞬时角速度,那么头朝向信号为
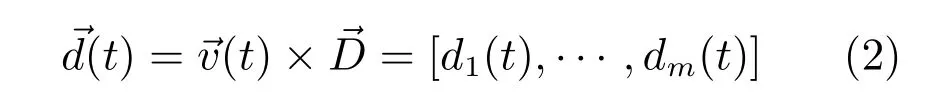
其中,m为头朝向细胞的个数,di(t)是头朝向细胞族中优先朝向为θi的第i个细胞在t时刻的头朝向信号.
1.2 条纹细胞计算模型
1.2.1 条纹向细胞的生理学研究基础
从发现网格细胞以来,其形成机理一直是研究热点.先前的研究模型中,速度信息被直接用于驱动网格细胞吸引子在神经板上的运动.2012年在傍下托和内嗅皮层浅皮层的细胞放电采样显示,存在着周期性条纹状放电野细胞,被称为条纹细胞,条纹细胞作为网格细胞上游皮层的细胞,被认为是形成网格细胞的基础神经元[22].傅里叶分析表明,多个条纹细胞族相互作用能形成周期性的网格细胞族放电野,被认为是形成网格细胞的基础机制[22].
Grossberg等以条纹细胞放电率作为输入信息,通过SOM 网络生成网格细胞和位置细胞的放电野[25−26,43],这种计算机制本质上属于振荡干涉模型,而振荡干涉模型在最近的研究中被认为缺乏生理学依据,而且在大范围空间环境表达中存在着累积误差大和鲁棒性不足的缺点[15].而采用连续吸引子模型生成网格细胞放电野的模型,并未考虑条纹细胞对网格细胞的影响,本文利用条纹细胞驱动网格细胞进行精确路径积分.
1.2.2 条纹细胞计算模型
在本文提出的模型中,条纹细胞编码了特定方向上的位移,是对动物在特定方向上相对位移的表征,其放电活动可由四个指标确定,如图4所示,分别是条纹间距L、条纹野宽度r、条纹相位(Δx,Δy)和优先朝向θ.沿一个方向上的线速度可以通过运动速度信号与头朝向方向的信号联合编码.对于有限数量的神经网络,对速度的积分不能无限期地持续,必须进行周期性的重置,并重新开始编码,条纹细胞必须被编码成一个周期性的放电细胞,负责对线速度进行路径积分操作.
本文使用一维环状吸引子模型对条纹细胞进行建模研究,如图5(a)所示,一个环状吸引子模型中的条纹细胞有着相同的优先朝向与周期,不同的位置选择性,如图5(b)所示.动物的运动速度在优先方向上的投影驱动吸引子运动,当这个投影为零时,吸引子位置保持不变,因此在这个时间间隔内,相同的细胞一直处于激活状态.环状吸引子模型有着长度的限制,当活动包返回到圆环的相同位置时,意味着动物在优先朝向上移动了特定的距离,将会使相同的条纹细胞再次激活.具有不同空间相位的条纹细胞沿着优先方向在不同的位置被周期性地最大程度地激活,如图5(b)所示,其放电野朝着相同的方向形成平行的条纹状覆盖动物经过的位置.条纹细胞环状吸引子的活动包在优先朝向完成一次移动的放电率与条纹间距成反比.

图4 条纹细胞的基础特征Fig.4 Basic characteristics of stripe cells
定义优先朝向在以我为中心方向θ,相位为α,放电周期为l的条纹细胞的放电率为xθα,那么该条纹细胞族在nl+α的相位处有最大的放电率.在t时刻动物沿着ϕ(t)方向上的移动速度为v(t),那么沿着θ方向的速度为
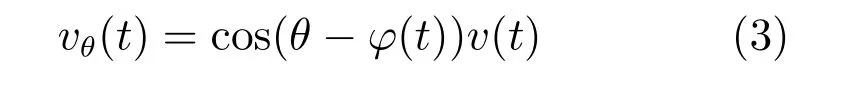
图5 条纹细胞的一维环状吸引子模型Fig.5 The model of one dimensional ring attractor of stripe cell
沿上述动物运动方向的位移可以由速度的积分得到
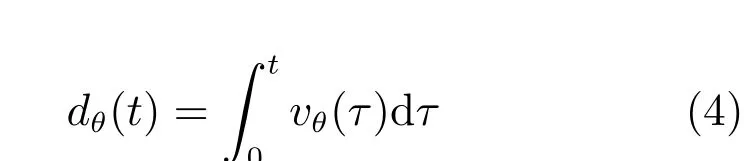
条纹细胞周期性放电重置的距离为

将条纹细胞的放电率建模成高斯模型,其放电率可以表示为
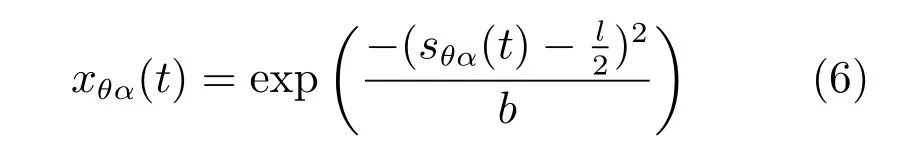
条纹细胞作为网格细胞吸引子的前向输入,是对输入信号的耦合,确定了细胞吸引子的移动方向.
1.3 网格细胞
1.3.1 网格细胞计算模型
网格细胞在动物探索环境的过程中周期性放电形成六边形的放电野覆盖整个环境,其放电活动具有高度的稳定性,在累计100米持续超过10分钟的运动后,仍然能持续放电;网格细胞的放电与外源性信息无关,只与自身的运动状态有关[44].网格细胞之间的连接是抑制性的[45],放电活动受来自于头朝向细胞和内嗅皮层中条纹细胞的信号调节.网格细胞的放电活动被认为能够形成空间环境的度量地图.网格细胞被认为是动物进行环境认知的路径积分器,是动物内在的位置预测的基础机制.
1.3.2 网格细胞精确路径积分计算模型
Burak等提出了一种网格细胞路径积分模型,能够在周期和非周期网络中实现精确路径积分[21],本文在此模型的基础上,加入了条纹细胞与网格细胞的前向连接,驱动网格细胞吸引子对环境的编码,这种结构更符合生理学研究事实.
网格细胞吸引子的形成受周围网格细胞递归连接权值和来自于条纹细胞的前向连接权值调节,使用二维连续吸引子模型对网格细胞进行建模.如图6(a)所示的神经板,包含有Nx×Ny个神经元,吸引子可由相应的神经元表示,随着探索路径的增大,表示空间相对位置的神经元不可能无限制的增加,这不可避免的涉及到吸引子网络的边界问题,为了解决边界问题,将该平面左右边界的神经元与上下边界的神经元相互连接,形成二维环状吸引子模型,如图6(b)所示.
由于内嗅皮层第二层的网格细胞之间的相互连接是抑制性的,在本文的模型中,每个网格细胞神经元从周围环状的局部神经元接收抑制性输入,整个网络接收来自上游皮层的前向兴奋性投射.在二维神经板中基于率编码的网格细胞神经元动力学等式为
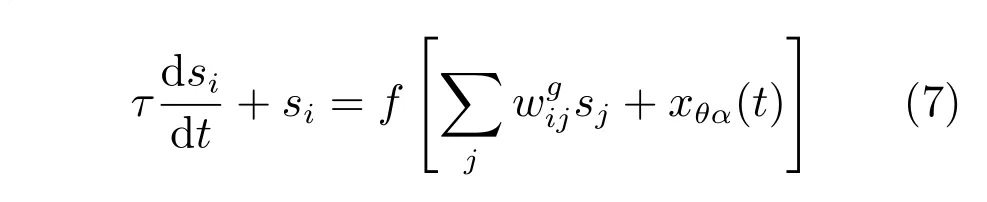
其中,τ是神经元相应的时间常量,神经元传递函数f[·]是一个简单的非线性整流函数,当x>0时,f(x)=x,当x≤0时,f(x)=0.当前位置神经元i的状态为是在该神经板中神经元j到神经元i的连接权值,是投射到神经元i的抑制性递归输入,xθα(t)是来自于条纹细胞的前向性兴奋性输入.如果神经板的长度为N,那么整个神经网络有N2个神经元,每个神经元i都有着一个优先方向,这个优先方向信息由条纹细胞的投射确定.
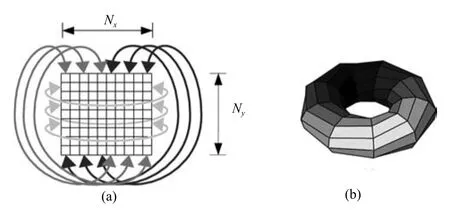
图6 二维环状吸引子模型的示意图Fig.6 The model of two dimensional ring attractor
网格细胞接收来自于条纹细胞的前向投射,前向投射中的优先朝向信息,被用于确定输出权值的改变方向,并确定其接收到老鼠的速度输入信息.网格细胞的递归连接权值为
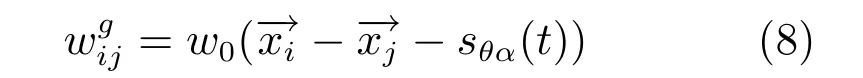
1.4 网格细胞到位置细胞计算模型
1.4.1 位置细胞的生理学研究基础
位置细胞能够对空间相对位置进行特征性的编码是脑内认知地图形成的基本单元,其放电活动提供了一个连续和动态的空间位置表达.大量位置细胞的联合响应(Combined activity)产生了一个对环境空间的离散表达,这说明位置细胞地图本质上属于路标地图[15].群放电活动在以我为中心的参考系下编码了动物的当前位置[6−7],建立了脑区和外界物理世界稳定的一一映射关系[2].位置细胞的放电特性如图7所示,实线段表示老鼠的运动轨迹,圆点表示位置细胞的激发点.

图7 位置细胞的放电情况Fig.7 The fi ring of place cells
网格细胞作为位置细胞的主要输入信息来源,位置细胞的放电活动是路径积分系统的输出.为了产生位置细胞族的单峰型放电野,需要学习网格细胞族到位置细胞的突触权值分布,确定在单个位置具有重叠活动包的网格细胞族的激活比例[46].这个过程可以通过具有确定的空间放电频率的网格细胞子集线性相加或者使用Hebb学习的方式和线性相加的方式.
课题组在先前的研究中构建了网格细胞到位置细胞的竞争性网络,采用竞争性网络进行自组织编码,成功模拟了齿状回中具有多个峰值的位置细胞位置野的特性.然后进一步经过Hebb学习产生了具有单个峰值的位置细胞位置野,模拟了CA3区(海马角的一部分)内的位置细胞特性[47].
1.4.2 网格细胞到位置细胞的竞争型网络模型
本文使用竞争性的Hebb学习寻找网格细胞族活动的子集,计算位置细胞族活动.
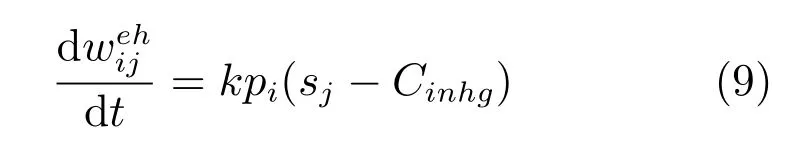
其中,k为学习率,pi为位置细胞的放电率,sj为网格细胞的放电率,Cinhg为网格细胞的抑制水平,由网格细胞平均放电率确定.式(9)的右边确定权重变化的方向:如果当前网格细胞激活率大于网格细胞的抑制性水平,突触连接增强;否则,突触连接发生抑制.
在一个神经板上的网格细胞群活动不足以完成路径积分,因为这会导致局部空间表达的歧义性.位置细胞对给定位置的空间选择性来源于对该位置联合编码的具有多个空间相位和空间间距的网格细胞活动的选择性结合.因此,需要多个空间间距和朝向的网格细胞,同样需要有多个空间间距和朝向的条纹细胞.每个神经板代表了一个网格细胞族活动,本文对a和λnet进行均匀采样产生多个不同尺度的网格细胞族.式(9)可以从多层神经板的空间重合检测网格细胞群体活动.
位置细胞的族活动来源于网格细胞的投射信息
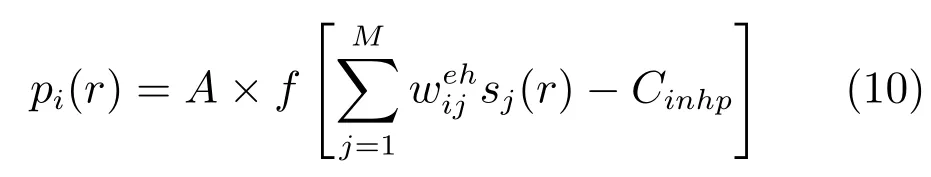
其中,A和Cinh分别为位置细胞网络的增益和抑制常量,M是网格细胞神经板的层数,是网格细胞族j到位置细胞族i的突触连接权值,r是当前动物的位置.Cinhp用来控制位置细胞放电野的个数,由B×max(pi(r))确定.通过上述的竞争性算法,确保了只有少量的网格细胞族的子集被选择形成位置细胞.
1.5 位置细胞的环境表达计算模型
位置细胞之间同样存在着相互作用,在局部的连接中,位置细胞通过抑制突触分支对周围的神经细胞产生抑制作用,网格细胞激活率大的子集生成的位置细胞的兴奋程度相对较强,通过侧抑制现象,对周围神经细胞的抑制作用也越强,最后的结果是兴奋作用最强的神经细胞在竞争中获胜,形成单一型的放电野.
为了实现位置细胞放电率在实际物理空间中的量化,采用二维的吸引子模型构建了对于实际外界环境相对位置的度量模型.二维连续吸引子模型由局部兴奋性、抑制连接和全局抑制性连接在一个神经板上形成一个随机的活动包(Activity bump),这个吸引子由空间细胞路径积分系统驱动,由来自于当前位置的图像信息进行重置.活动包如图8中灰色神经元所示,如果整个位置细胞网络有边界,那么当机器人离开位置细胞网络覆盖的区域时,整个网络将不能表示当前机器人的位置信息,所以,类似于网格细胞的环状吸引子模型,网络边界的位置细胞与另一边界的位置细胞相连接,形成环状.
一个二维的高斯分布被用于创建位置细胞的兴奋性权值连接矩阵εm,n,其中下标m和n代表在坐标系X和Y中所属单元横纵坐标之间的距离.权值分布可表示为

其中,kp为位置分布的宽度常量.
图8 位置细胞的连续吸引子模型表达Fig.8 The expression of the continuous attractor model of place cells
由于局部兴奋性连接导致的位置细胞活动的变化量为

其中,nX和nY是在(X,Y)空间中位置细胞二维矩阵的大小,代表吸引子模型在神经板上活动的范围.因为网络中位置细胞的无边界性,如图8所示,处于神经板边界的位置细胞会与相对边界的位置细胞产生兴奋性连接,而进行位置细胞迭代和视觉模板匹配的前提是查找位置细胞吸引子在神经板中的相对位置,这个相对位置坐标由权值矩阵的下标表示,可由下式计算得到

每个位置细胞同样接收着整个网络的全局性抑制信号.与网格细胞所形成的墨西哥帽吸引子模型不同,位置细胞的抑制信号发生在局部性兴奋连接之后,而不是同时起作用.兴奋性和抑制性连接矩阵的对称性保证了合适的神经网络动力学,确保空间中的吸引子不会无限制地兴奋.位置细胞由抑制性连接权值引起的活动变化量为
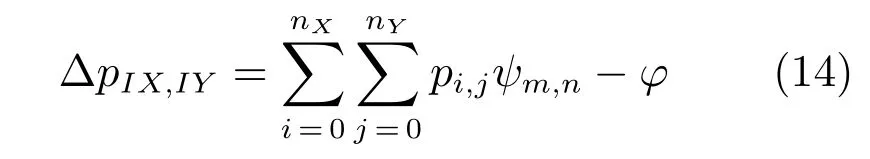
其中,ψm,n是抑制性连接权值,ϕ控制全局性的抑制水平.所有位置细胞的活动都是非零的并且进行归一化.
为确保所有时刻的所有位置细胞的活动都是不小于零的,将位置细胞的激活率与0进行比较
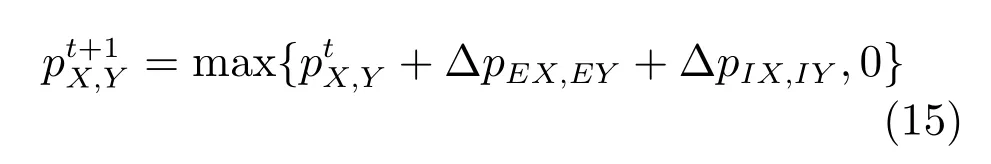
随后将位置细胞的活动率进行归一化

在本文模型中,位置细胞吸引子的移动来自于空间细胞对自运动线索的路径积分,条纹细胞编码了特定方向上的位移,驱动网格细胞吸引子的移动,网格吸引子对特定方向上的二维空间进行编码,使得不同的网格细胞族激活.不同的网格细胞族活动的子集确定了位置吸引子的移动.这一过程的示意图如图9所示.
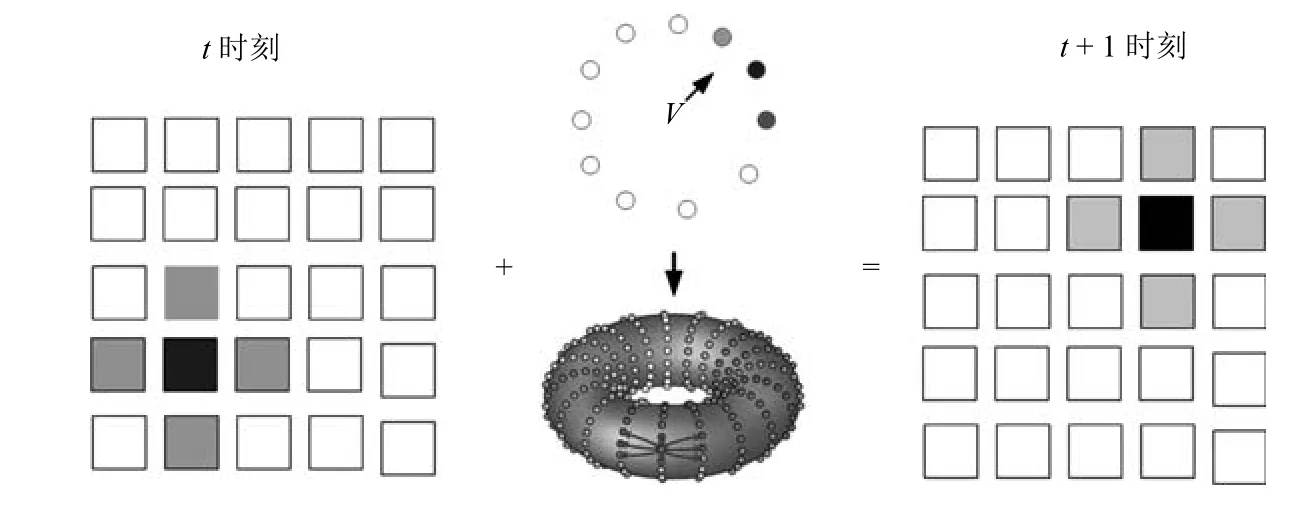
图9 位置细胞路径积分示意图Fig.9 Place cell path integral

其中,δX0,δY0是X-Y坐标系中向下取整的偏置量,这一偏置量由速度和方向信息确定.
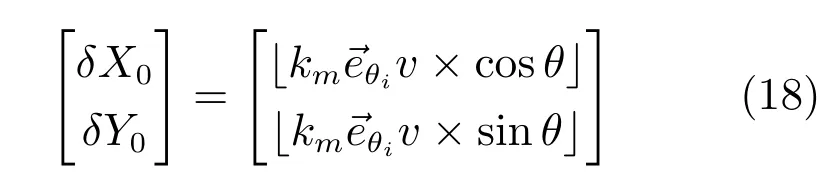
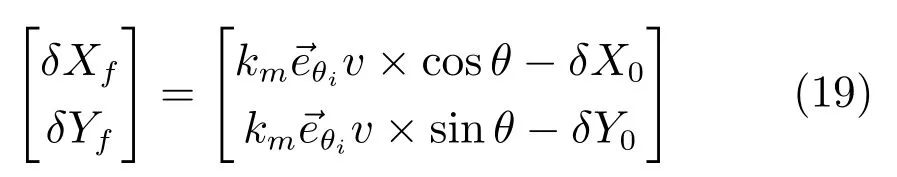
残差量是残差偏置量的分段函数:
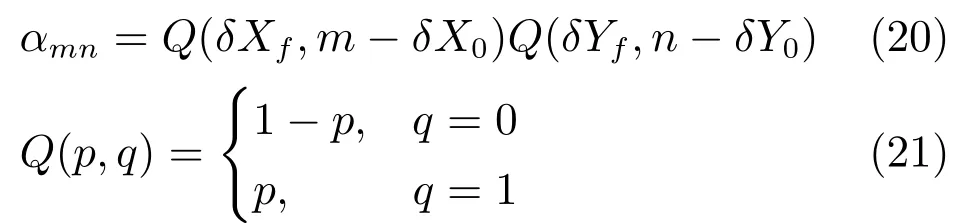
1.6 视觉图像处理与闭环检测
使用自运动线索进行路径积分在大范围的空间表达中,不可避免地会存在误差累积.研究表明,老鼠在遇到显著路标点时会对路径积分误差进行纠正[47],本文引入颜色深度图像构建环境视图模板进行闭环检测,纠正路径积分误差.
本文采用视图模板匹配算法实现闭环检测,利用颜色和深度图像中的扫描线强度分布(Scanline intensity pro fi le)[32]进行视图模板匹配.扫描线强度分布是一维向量,它是对灰度图片所有像素列强度求和并归一化的结果.图10(a)和图10(b)是一幅图像的颜色图和深度图.
两个图像扫描线强度分布间的平均绝对强度差称为强度偏移量,用g(c)表示.
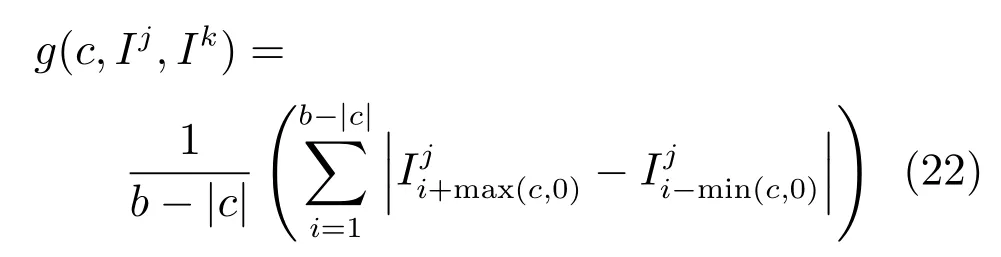
其中,Ij和Ik是被比较图像的扫描线强度分布,c是分布偏移(Pro fi le shift)量,b是图像的宽度.
由于图像匹配受光照影响较大,为了减弱光照条件的影响,提高在不同环境下的图像匹配精度,本文采用颜色图和深度图两种图像同时匹配来确定绝对位置的方法.由于实际空间环境在不同时间段光照强度有所不同,对颜色图和深度图的偏移量之差赋予不同的权值,可以得到图像的匹配度度量G.

其中,µR和µD分别为颜色图和深度图的权值,且µR+µD=1.
在连续的图像中Ij和Ik像素的最小偏移量cm是对两幅图像的匹配度度量G取最小值.

图10 图像与扫描线强度分布图Fig.10 Image and scanning line intensity distribution

其中,偏置ρ确保了两幅图像有一个重叠量.设定图像的比较阈值为ct,当cm<ct时,当前视图为新的视图,保存到视觉模板集{Vi}中,当cm≥ct时,认为回到了一个重复的场景.
1.7 认知地图的构建与修正
本文模型构建的认知地图属于半拓扑地图,同样建立位置细胞放电活动位置之间的拓扑关系,由具有拓扑关系的认知点e组成,认知点之间的拓扑联系由tij表示.每个认知点包含当前位点位置细胞放电活动pi、视觉模板Vi以及位置间的拓扑关系.单个认知点定义为
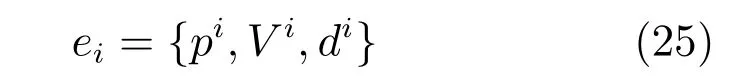
1)认知地图的构建
设定认知阈值为Sth,当前位置与存在的认知点中的位置比较,能够得到一个位置度量S.

当前认知点的位置度量超过认知阈值或者当发现新的视觉模板时,新的认知点被创建.转换量tij存储由路径积分计算的位置改变量,即

其中,tij形成了先前经验与新的经验点的连接关系,新的认知点j可表示为
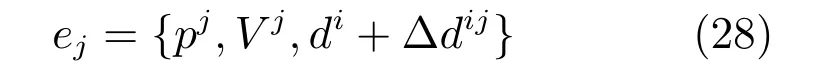
2)闭环处的认知地图更新
当视图模板检测到实际的闭环点时,认为机器人回到了相同的位置,然而位置变量的累加量在闭环处所形成的新的经验与这个相同位置是不相匹配的,为了达到两者的匹配,需要在闭环处对所有的经验进行更新.
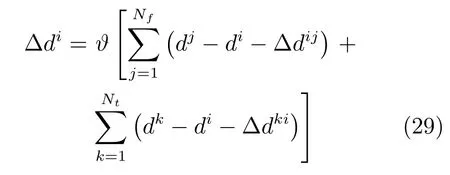
其中,ϑ是一个纠正率常量,Nf是从认知点ei到其他认知点的转移个数,Nt是从其他认知点到当前认知点ei的转移个数.在实际的实验中取ϑ=0.5,更大取值将会导致整个地图的不稳定.整个地图的更新过程是连续的,但在闭环处最为明显.为了保证构建地图的准确性,只有在连续两帧图像检测到闭环时,才对整体的认知地图进行调整.
2 实验验证
利用仿真和物理实验对构建的模型进行实验验证.为验证构建的细胞模型的有效性,采用实际机器人采集的数据、真实的生理学数据和仿真实验数据进行实验验证,生理学数据来源于Hafting等在Nature上发表的“Microstructure of a spatial map in the entorhinal cortex”中公布的老鼠在直径为1m的圆形环境中运行的真实轨迹信息,截取了部分轨迹用于仿真实验.仿真实验数据是让智能体(Agent)在自定义的环境中,以0.5m/s的速度,朝着任意方向在直径为4m圆形的环境中运行的速度和方向信息.真实的实验数据是利用先锋3-DX机器人与Kinect组合采集的速度、方向以及环境的颜色深度信息.
2.1 机器人平台
实验采用的平台主体是先锋3-DX机器人,移动装置是两个前轮和一个后轮,后轮是一个小型的万向轮,方便机器人的稳定支撑和变相.前轮配备有光电编码器,能采集和记录机器人的移动距离.
Kinect置于平台面板上方,使用逆变器供电,在机器人移动的时候,进行RGB-D图像的采集. Kinect直接与电脑连接,以2帧/s的速率采集RGB和DEPTH图像各一张.
整个平台通过USB口与机器人进行通讯,数据采集时,使用电脑控制机器人在环境中移动,采集速度、方向以及该位置点的图像信息.采样周期是500ms.移动机器人平台最大移动速度设定为0.5m/s.
先锋机器人主要使用其自带的Aria系统框架,上位机主要调用其通信SDK来实现对先锋机器人的控制与数据采集.Kinect主要使用开源的OpenNI2开源3D视觉框架,进行颜色与深度图像采集.
2.2 参数设定
头朝向细胞的个数设定为360个,λnet是对从12∼52之间间隔为1的均匀分布采样.w0的调节向量a=1.1,学习率k为0.0005,抑制性系数B= 0.5.位置细胞与视图模板比较的相关参数设定如表1所示.
2.3 空间细胞计算模型的实验验证
条纹间距决定网格节点的间距,条纹朝向决定网格细胞输出权值的改变方向和速度信号的调节方向[48−49].调节条纹间距可以得到不同朝向的条纹细胞放电野,在直径为4m的圆形平面中,智能体速度大小设置为0.5m/s,移动的时间为100s.条纹间距为10cm,35cm和50cm,优先朝向分别为0°,60°和90°的条纹细胞放电率图如图11所示.从第1行到第3行分别是10cm,35cm和50cm条纹间距放电率图,从第1列到第3列分别对应了条纹的优先朝向.结果表明,条纹细胞的放电野能够形成对空间特定方向位移的表达,本文提出的计算模型能够在不同的空间尺度上对条纹细胞的放电特征进行描述,说明了计算模型的有效性.

图11 不同朝向的条纹细胞的放电率图Fig.11 Firing rate of stripe cells with diあerent orientation

表1 参数设置
条纹间距决定网格间距,采用不同的条纹间距.将条纹细胞的条纹朝向设置为网格细胞族的放电活动能够覆盖整个平面,记录单个网格细胞的在一个周期内的放电情况以及单个神经元在直径4m的圆环型平面上移动的轨迹和放电情况,可以看到,以0.2m/s的速度移动1200s后,不同间距的单个网格细胞都可以生成六边形的放电野覆盖整个空间,如图12所示,在运动轨迹上有许多放电脉冲的位置.随着变大,网格野间距变大,这与生理学研究中,网格野沿着背腹侧轴网格野间距递增的生理学事实一致.单个网格细胞能够以不同的尺度周期性的表达环境,在长时间的路径积分中,其放电野保持六边形覆盖整个空间,这说明了网格细胞路径积分过程中的误差累积小于半个周期的相位误差,不足以破坏网格细胞的放电特性,说明了计算模型的有效性与准确性.

图12 网格细胞的精确路径积分Fig.12 Exact path integration of grid cells

图13 老鼠移动轨迹上网格细胞和位置细胞放电率图Fig.13 The fi ring rate of place cells and grid cells based on mouse trajectory
利用Hafting采集的真实老鼠数据进行位置细胞计算模型有效性检测,截取2s的轨迹进行仿真实验如图13所示,第1行分别是原始轨迹图、位置细胞放电率图和网格细胞到位置细胞权值连接图;第2行是不同朝向和尺度的网格细胞放电率图.可以看到在平面的相应位置,位置细胞产生了单一型的位置野,网格细胞与位置细胞之间的权值连接,使用放电率图表示,特定范围内的网格细胞通过竞争学习,选择了相应的网格子集确定了位置细胞.
2.4 认知地图构建方法物理实验验证
利用机器人平台对实验室10m×3m的范围持续探索170秒,实际的运动轨迹如图14中画线部分所示.认知地图构建算法如图15所示.
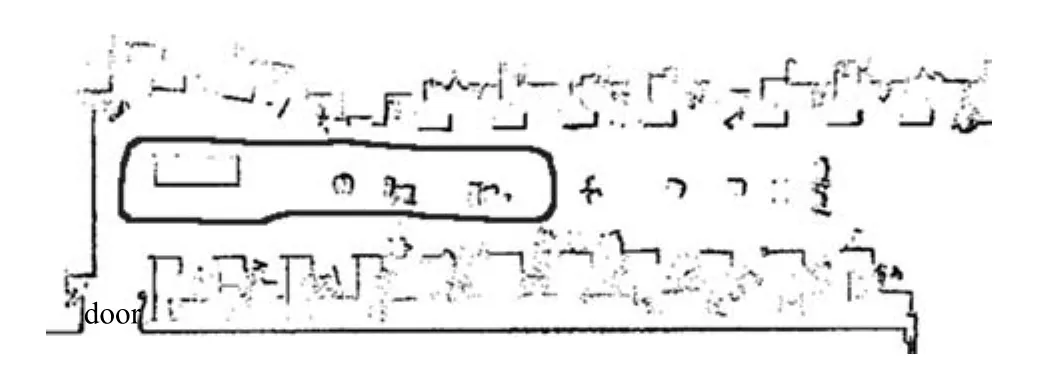
图14 实际物理环境扫描图Fig.14 Actual physical environment scanning
根据表1的相关参数设定,构建地图的过程如图16所示.第1行是机器人采集的里程计地图,第2行是根据图15所示的算法构建认知地图的过程.可以看到,当没有检测到闭环点时,里程计地图和认知地图并无差别,在89s时检测到了环境的闭环点,认知地图在第89.5s进行调整,随着时间的推移,里程计地图误差越来越大,而认知地图因为进行了闭环检测,越来越符合实际的活动轨迹.
闭环检测与空间细胞放电重置的过程如图17所示,图17(a)为7.5s时的认知地图和位置细胞放电率,在89s时,检测到环境的闭环点(图17(b)),在89.5s时模型进行认知地图的调整(图17(c)),并进行放电率重置,值得注意的是图17的位置细胞族放电活动与认知地图点(图中第1列画圈部分所示)并不完全一致,这是因为位置细胞族活动表达的是机器人在环境中的相对位置.
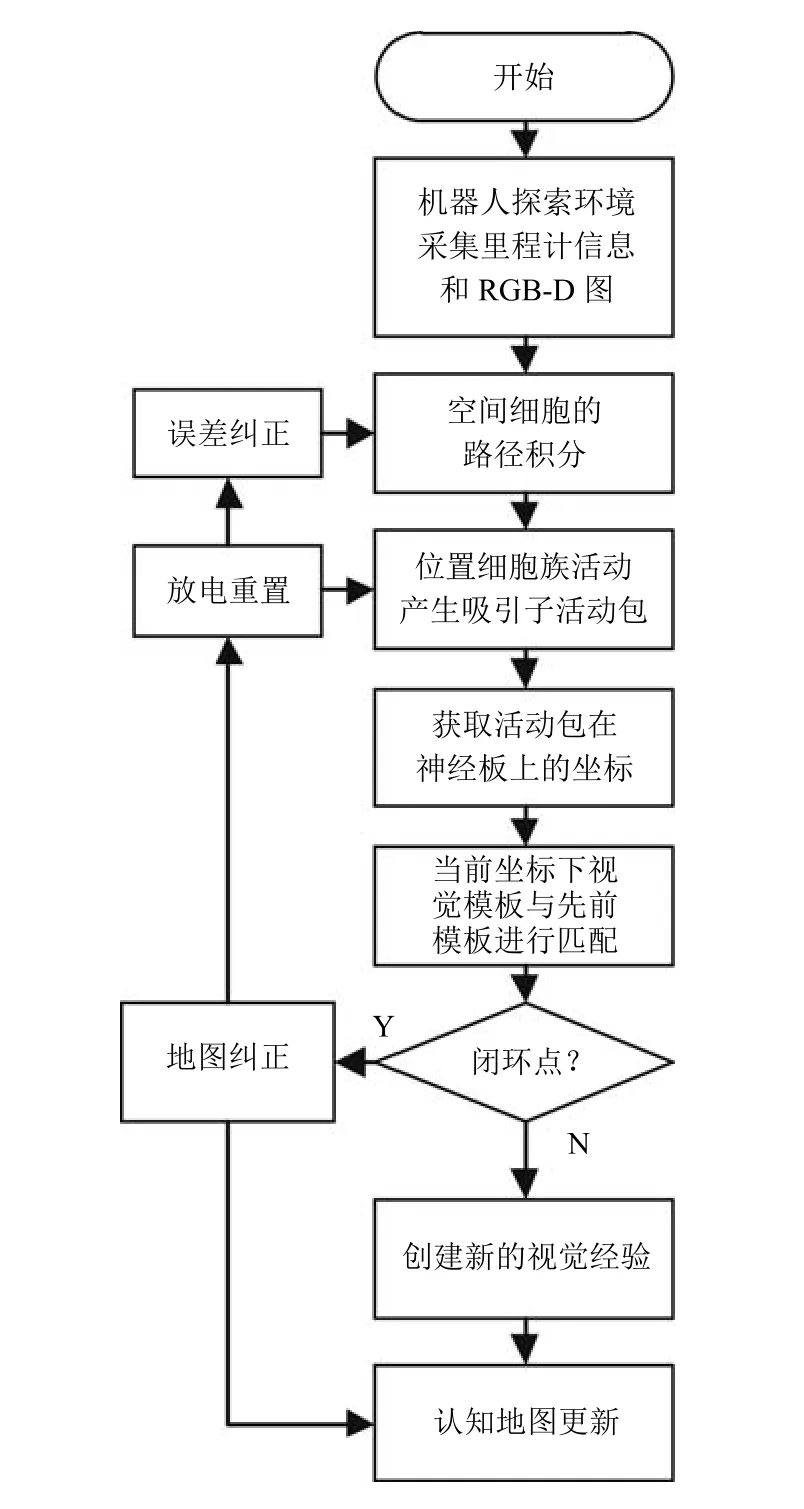
图15 算法流程图Fig.15 Algorithm fl ow chart

图16 地图的构建过程Fig.16 The map building process
3 实验结果分析与讨论

图17 闭环检测与空间细胞放电重置过程Fig.17 Closed loop detection and space cell discharge reset process
本文使用条纹细胞前向驱动网格细胞吸引子进行周期性的位置表达,使网格细胞权值调节在优先方向上不仅仅局限于四个方向,更加具有生理学依据,通过网格细胞的子集产生位置细胞族活动,并通过位置细胞之间的相互作用形成了单一型的放电野,对环境进行表达.
3.1 位置细胞的递归连接对模型的影响
先前的位置细胞计算方法中,仅通过网格细胞子集产生的位置细胞放电野总是存在多峰值放电现象[24](如图18(a)、图18(b)和图18(c)右图).在本文的模型中,通过位置细胞之间的相互竞争作用产生单峰型的放电野(如图18(a)、图18(b)和图18(c)左图),更加符合生理学研究事实.
3.2与RATSLAM和传统SLAM实验对比
3.2.1与RATSLAM进行实验对比

图18 位置细胞相互竞争前后的放电率图Fig.18 The fi ring rate before and after the place cell competition
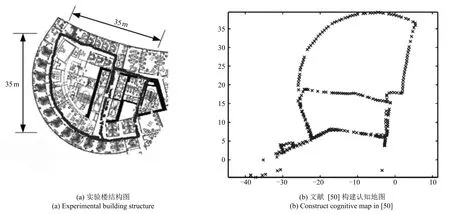
图19 文献[50]实验结果Fig.19 Experimental results in[50]
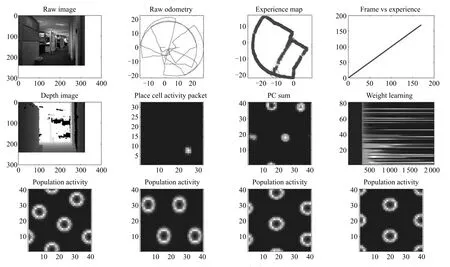
图20 本文算法运行实验结果Fig.20 The results of running the algorithm
与RATSLAM对比实验,采用文献[50]中的数据集进行实验验证,与文献[50]构图效果进行对比.文献[50]是第一次采用RGB-D图像作为外部信息用于RATSLAM认知地图构建,此前昆士兰大学的Milford等多采用单彩色摄像头和全景摄像头进行数据采集.通过与文献[50]的认知地图构建结果(图19所示)进行对比,验证本文算法.该数据集包含了机器人在半径为35m办公楼探索1050s的里程计和RGB-D图信息,实验结果如图20所示.
从图19和20对比可以看出,与RATSLAM相比,本文算法同样可以精确构建出地图,虽然数据含有严重累计误差.空间细胞的路径积分在大规模的环境表达中,不可避免地存在累积误差,本文通过外界的视觉线索进行闭环检测,纠正累积误差,生成环境精确的认知地图.与RatSLAM算法相比,采用条纹细胞驱动网格细胞对速度和方向信号进行路径积分,采用颜色深度图进行闭环检测,更加符合生理学研究事实[51].
3.2.2 与传统SLAM进行实验对比
对比实验平台,采用开源硬件Turtlebot2的ROS系统中自带的Navigation SLAM模块进行传统SLAM导航与本文算法作对比.实验环境16.8m×8.46m的矩形走廊,对比机器人平台Turtlebot2 (编码器、陀螺仪),Kinect传感器.设定最大速度为0.5m/s,围绕走廊运行2圈,构建2D认知地图.本文算法运行平台参数设定如上文所述,由于光照变弱,µR设为0.55,µD设为0.45.实验结果如图21所示.

图21 第1次对比实验结果Fig.21 The fi rst comparative experimental results
如图21所示,两种方法构建地图并不能完全正确构建出矩形走廊地图.但本文算法效果明显优于ROS系统自带Navigation SLAM算法.根据环境分析,可能是因为走廊环境相似度太高,不利于算法进行闭环检测,导致两种算法都无法正确构建环境地图.本文采用的闭环点检测的的方法是基于图像的强度分布线进行检测,比较简单,后期会考虑g2o闭环检测方法,进一步研究提高其性能.
在第二次对比实验中,在走廊中设置了一些特定的路标(如图22(a)放置箱子、椅子等障碍物),提高构建地图的准确性,如图22(b)和图22(c)所示.在加入一些特定的路标之后,两种方法都能消除累计误差,精确地构建出走廊的认知地图.与传统的SLAM方法相比,在没有明显路标的环境中,明显比ROS自带的传统SLAM方法构建的地图更加精确.
在计算成本方面,虽然有些是离线数据集实验,但是从程序完成的时间上来看,基本可以达到实时构建的程度.考虑到目前计算环境主要是Matlab环境,代码的运行效率较低.计算速度和成本还有很大的改进空间.
3.3 精确路径积分的量化分析
3.3.1 仿真理论分析
条纹细胞驱动网格细胞对速度和方向信号进行路径积分,产生网格细胞六边形放电野覆盖机器人经过的空间环境,对环境进行特征性表达.仿真环境下机器人的位姿信息是随机产生的速度信息和当前头朝向信息.该仿真位姿信息经过一些微分算法处理,去除不真实的情况(例如速度、头朝向突变),确保随机轨迹符合生物运动规律.为了评估模型对环境表达的精确性,以网格野节点的歧义性作为衡量指标,当网格节点偏差超过网格间距的一半时,网格节点之间的环境表达产生歧义,此时,网格细胞无法对环境进行精确的表达.改变λnet的大小,控制网格间距的大小,使智能体在直径4m的圆形环境中连续运动1200s,其移动速度为0.2m/s.
图23是网格间距为4cm时单个神经元的响应图,当设置条纹间距为4cm时,单个网格细胞的响应已经崩溃.其结果与图12对比,可以看到当网格间距大于9cm时,网格细胞都能够生成六边形放电野覆盖整个环境,在运动距离达到240m时,仿真结果表明,累计误差最多不超过9cm,体现了路径积分的精确性.
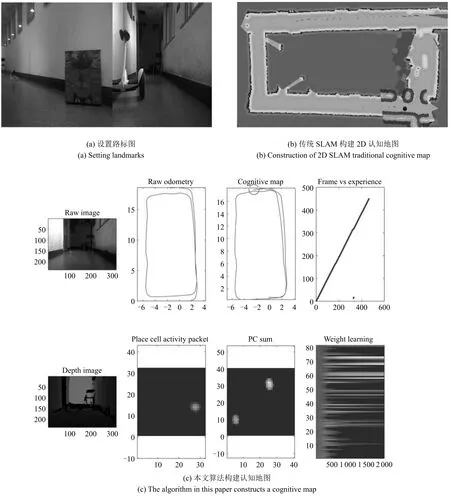
图22 第2次对比实验结果Fig.22 The second comparative experimental results
3.3.2 真实环境路径积分结果分析
为了证明条纹细胞驱动网格细胞对速度和方向信号进行路径积分在实际应用中的准确性.本文设计实验在长16.8m的走廊中,采集实验机器人编码器以及陀螺仪数据获取真实位姿信息,进行纯路径积分实验与利用该细胞的模型的路径积分的比较实验.为了增加实验难度,机器人速度不设为匀速,手动随机控制,机器人的运行速度最大为0.5m/s.实验如图24所示.
由实验结果可得,普通纯路径积分方法结果为x=16.57,y=0.5642,基于细胞模型路径积分结果为x=16.88,y=0.5835.实际走廊长度为16.8m,很明显在相同数据的条件下,基于细胞模型的路径积分精度要比普通纯路径积分高.这主要是因为在条纹细胞驱动网格细胞对速度和方向信号进行路径积分的过程中,对于不合法或者突变的数据有一定的滤波整合效果,在一定程度上弥补传感器以及路面不平带来的影响,从而提高路径积分精度.但此方法也不能去除累计误差和积分漂移对路径积分带来的影响.

图23 网格间距为4cm时单个神经元的响应图Fig.23 The response of a single neuron when the grid space is 4cm
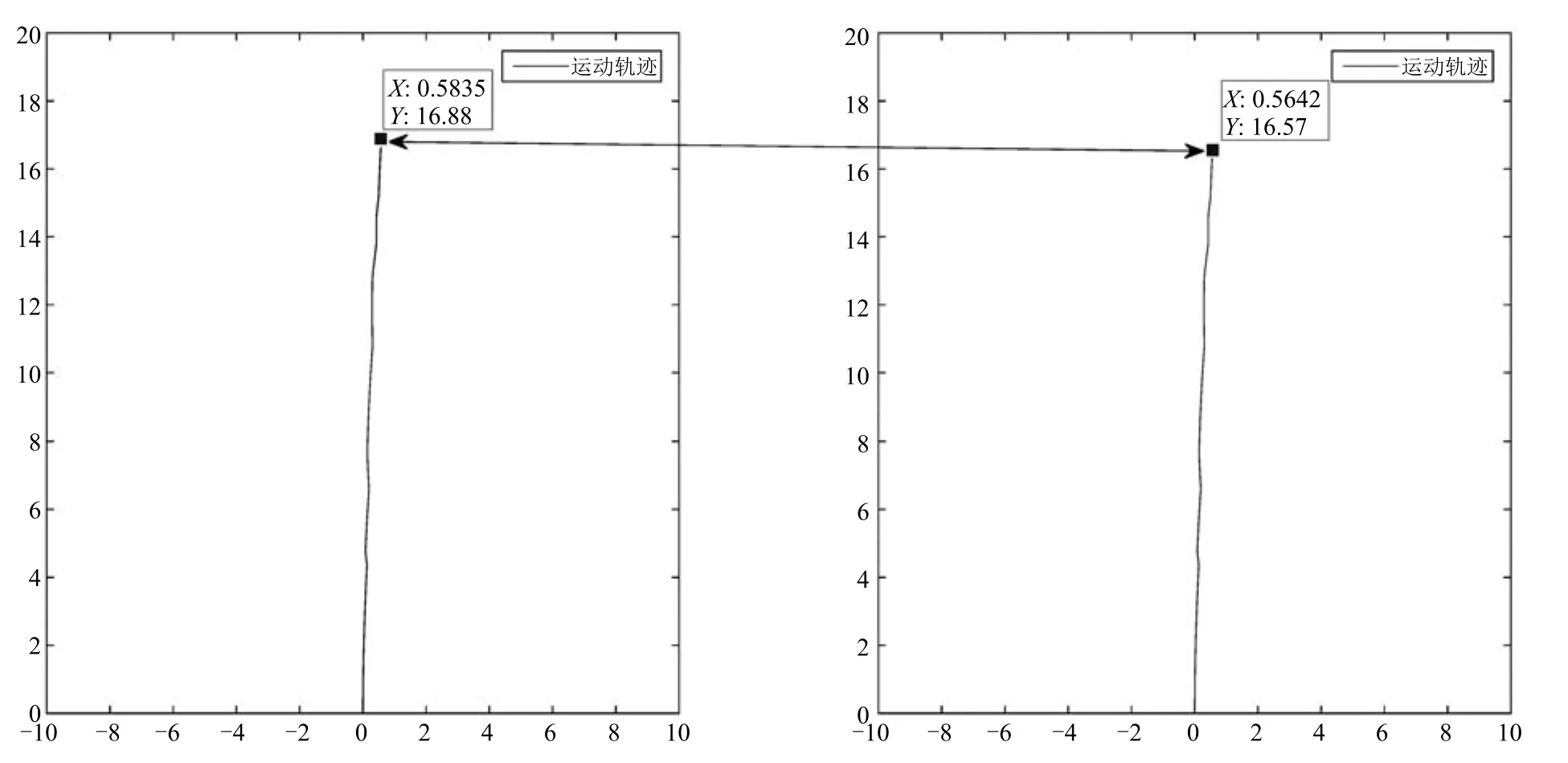
图24 真实环境路径积分精度结果Fig.24 Real environment path integral precision result
4 结论
本文依照海马空间细胞的环境认知机理,采用吸引子模型对4种空间细胞进行了建模研究,通过空间细胞的路径积分对环境的特征位点进行了表达,利用颜色深度图进行了环境的闭环检测,提出的方法能够构建实际物理空间的精确认知地图.通过与传统SLAM和RATSLAM对比,本文算法在性能结果方面都不输于前者.而且本文提出的认知地图构建方法完全依据海马空间细胞的生理学研究机理,为进一步研究机器人仿生导航方法奠定了基础.
1 Tolman E C.Purposive Behavior in Animals and Men.New York,USA:University of California Press,1932.
2 McNaughton B L,Battaglia F P,Jensen O,Moser E I,Moser M B.Path integration and the neural basis of the“cognitive map”.Nature Reviews Neuroscience,2006,7(8):663−678
3 Steele R J,Morris R G.Delay-dependent impairment of a matching-to-place task with chronic and intrahippocampal infusion of the NMDA-antagonist D-AP5.Hippocampus, 1999,9(2):118−136
4 Steあenach H A,Witter M,Moser M B,Moser E I.Spatial memory in the rat requires the dorsolateral band of the entorhinal cortex.Neuron,2005,45(2):301−313
5 Taube J S,Kesslak J P,Cotman C W.Lesions of the rat postsubiculum impair performance on spatial tasks.Behavioral and Neural Biology,1992,57(2):131−143
6 O′Keefe J,Dostrovsky J.The hippocampus as a spatial map.Preliminary evidence from unit activity in the freelymoving rat.Brain Research,1971,34(1):171−175
7 O′Keefe J.Place units in the hippocampus of the freely moving rat.Experimental Neurology,1976,51(1):78−109
8 Tsao A,Moser M B,Moser E I.Traces of experience in the lateral entorhinal cortex.Current Biology,2013,23(5):399−405
9 Taube J S,Muller R U,Ranck Jr J B.Head-direction cells recorded from the postsubiculum in freely moving rats.II. Eあects of environmental manipulations.The Journal of Neuroscience,1990,10(2):436−447
10 Sharp P E.Multiple spatial/behavioral correlates for cells in the rat postsubiculum:multiple regression analysis and comparison to other hippocampal areas.Cerebral Cortex, 1996,6(2):238−259
11 O′Keefe J,Burgess N.Geometric determinants of the place fi elds of hippocampal neurons.Nature,1996,381(6581): 425−428
12 Mittelstaedt M L,Mittelstaedt H.Homing by path integration in a mammal.Naturwissenschaften,1980,67(11):566−567(in German)
13 Hafting T,Fyhn M,Molden S,Moser M B,Moser E I.Microstructure of a spatial map in the entorhinal cortex.Nature,2005,436(7052):801−806
14 Solstad T,Moser E I,Einevoll G T.From grid cells to place cells:a mathematical model.Hippocampus,2006,16(12): 1026−1031
15 Moser E I,Roudi Y,Witter M P,Kentros C,Bonhoeあer T, Moser M B.Grid cells and cortical representation.Nature Reviews Neuroscience,2014,15(7):466−481
16 Burgess N,Barry C,O′Keefe J.An oscillatory interference model of grid cell fi ring.Hippocampus,2007,17(9):801−812
17 Hasselmo M E,Giocomo L M,Zilli E A.Grid cell fi ring may arise from interference of theta frequency membrane potential oscillations in single neurons.Hippocampus,2007, 17(12):1252−1271 18 Si B,Romani S,Tsodyks M.Continuous attractor network model for conjunctive position-by-velocity tuning of grid cells.PLoS Computational Biology,2014,10(4):e1003558
19 Erdem U M,Hasselmo M.A goal-directed spatial navigationmodel using forward trajectory planning based on grid cells.European Journal of Neuroscience,2012,35(6):916−931
20 Fuhs M C,Touretzky D S.A spin glass model of path integration in rat medial entorhinal cortex.The Journal of Neuroscience,2006,26(16):4266−4276
21 Burak Y,Fiete I R.Accurate path integration in continuous attractor network models of grid cells.PLoS Computational Biology,2009,5(2):e1000291
22 Krupic J,Burgess N,O′Keefe J.Neural representations of location composed of spatially periodic bands.Science,2012, 337(6096):853−857
23 Raudies F,Hasselmo M E.Diあerences in visual-spatial input may underlie diあerent compression properties of fi ring fi elds for grid cell modules in medial entorhinal cortex.PLoS Computational Biology,2015,11(11):e1004596
24 Yuan M L,Tian B,Shim V A,Tang H,Li H.An entorhinalhippocampal model for simultaneous cognitive map building.In:Proceedings of the 29th AAAI Conference on Artifi cial Intelligence.Austin,TX:AAAI,2015.
25 Grossberg S,Pilly P K.How entorhinal grid cells may learn multiple spatial scales from a dorsoventral gradient of cell response rates in a self-organizing map.PLoS Computational Biology,2012,8(10):e1002648
26 Pilly P K,Grossberg S.Spiking neurons in a hierarchical self-organizing map model can learn to develop spatial and temporal properties of entorhinal grid cells and hippocampal place cells.PLoS One,2013,8(4):e60599
27 Cuperlier N,Quoy M,Gaussier P.Neurobiologically inspired mobile robot navigation and planning.Frontiers in Neurorobotics,2007,1:Article No.3
28 Jauあret A,Cuperlier N,Gaussier P.Multimodal integration of visual place cells and grid cells for robots navigation.In: Proceedings of the 2nd Symposium on Biology of Decision Making.Paris,France:BDM,2012.136−145
29 Milford M J,Wyeth G F,Prasser D.RatSLAM:a hippocampal model for simultaneous localization and mapping.In: Proceedings of the 2004 IEEE International Conference on Robotics and Automation.New Orleans,LA,USA:IEEE, 2004.403−408
30 Milford M,Wyeth G,Prasser D.RatSLAM on the edge:revealing a coherent representation from an overloaded rat brain.In:Proceedings of the 2006 IEEE/RSJ International Conference on Intelligent Robots and Systems.Beijing,China:IEEE,2006.4060−4065
31 Milford M J,Wyeth G F.Mapping a suburb with a single camera using a biologically inspired SLAM system.IEEE Transactions on Robotics,2008,24(5):1038−1053
32 Milford M J,Wyeth G F.Single camera vision-only SLAM on a suburban road network.In:Proceedings of the 2008 IEEE International Conference on Robotics and Automation.Pasadena,CA,USA:IEEE,2008.3684−3689
33 Erdem U M,Hasselmo M E.A biologically inspired hierarchical goal directed navigation model.Journal of Physiology, Paris,2014,108(1):28−37
34 WangNan,MaShu-Gen,LiBin,WangMing-Hui, Zhao Ming-Yang.A model transformation of map representation for hierarchical SLAM that can be used for after-earthquake buildings.Acta Automatica Sinica,2015, 41(10):1723−1733
(王楠,马书根,李斌,王明辉,赵明扬.震后建筑内部层次化SLAM的地图模型转换方法.自动化学报,2015,41(10):1723−1733)
35 Xiao Yun-Tao,Ou Lin-Lin,Yu Li.Optimal patrolling path planning via linear temporal logic.Acta Automatica Sinica, 2014,40(10):2126−2133
(肖云涛,欧林林,俞立.基于线性时序逻辑的最优巡回路径规划.自动化学报,2014,40(10):2126−2133)
36 Xu Bo,Bai Jin-Lei,Hao Yan-Ling,Gao Wei,Liu Ya-Long. The research status and progress of cooperative navigation for multiple AUVs.Acta Automatica Sinica,2015,41(3): 445−461
(徐博,白金磊,郝燕玲,高伟,刘亚龙.多AUV协同导航问题的研究现状与进展.自动化学报,2015,41(3):445−461)
37 Chen Cheng,He Yu-Qing,Bu Chun-Guang,Han Jian-Da.Feasible trajectory generation for autonomous vehicles based on quartic b´ezier curve.Acta Automatica Sinica, 2015,41(3):486−496
(陈成,何玉庆,卜春光,韩建达.基于四阶贝塞尔曲线的无人车可行轨迹规划.自动化学报,2015,41(3):486−496)
38 Ranck J B.Head direction cells in the deep cell layer of dorsolateral presubiculum in freely moving rats.Electrical Activity of the Archicortex,Budapest:Akad´emiai Kiad´o, 1984.
39 Raudies F,Chapman W,Brandon P,Hasselmo M E.Head direction is coded more strongly than movement direction in a population of entorhinal neurons.Brain Research,2015, 1621:355−367
40 MizumoriS J,WilliamsJ D.Directionally selective mnemonic properties of neurons in the lateral dorsal nucleus of the thalamus of rats.The Journal of Neuroscience, 1993,13(9):4015−4028
41 Taube J S,Muller R U,Ranck Jr J B.Head-direction cells recorded from the postsubiculum in freely moving rats.I. Description and quantitative analysis.The Journal of Neuroscience,1990,10(2):420−435
42 Knierim J J,Kudrimoti H S,McNaughton B L.Place cells, head direction cells,and the learning of landmark stability.The Journal of Neuroscience,1995,15(3 Pt 1):1648−1659
43 KropあE,Carmichael J E,Moser M B,Moser E I.Speed cells in the medial entorhinal cortex.Nature,2015,523(7561): 419−424
44 Juavinett A L,Callaway E M.Pattern and component motion responses in mouse visual cortical areas.Current Biology,2015,25(13):1759−1764
45 Couey J J,Witoelar A,Zhang S J,Zheng K,Ye J,Dunn B,Czajkowski R,Moser M B,Moser E,Roudi Y,Witter M P.Recurrent inhibitory circuitry as a mechanism for grid formation.Nature Neuroscience,2013,16:318−324
46 Si B L,Treves A.The role of competitive learning in the generation of dg fi elds from ec inputs.Cognitive Neurodynamics,2009,3(2):177−187
47 Yu Nai-Gong,Wang Lin,Li Ti,Chen Huan-Zhao.Competitive neural network model from grid cells to place cells.Control and Decision,2015,30(8):1372−1378
(于乃功,王琳,李倜,陈焕朝.网格细胞到位置细胞的竞争型神经网络模型.控制与决策,2015,30(8):1372−1378)
48 Bonnevie T,Dunn B,Fyhn M,Hafting T,Derdikman D, Kubie J L,Roudi Y,Moser E I,Moser M B.Grid cells require excitatory drive from the hippocampus.Nature Neuroscience,2013,16(3):309−317
49 Winter S S,Mehlman M L,Clark B J,Taube J S.Passive transport disrupts grid signals in the parahippocampal cortex.Current Biology,2015,25(19):2493−2502
50 Tian B,Shim V A,Yuan M L,Srinivasan C,Tang H J,Li H Z.RGB-D based cognitive map building and navigation. In:Proceedings of the 2013 IEEE/RSJ International Conference on Intelligent Robots and Systems(IROS).Tokyo, Japan:IEEE,2013.1562−1567
51 Pan Chao,Liu Jian-Guo,Li Jun-Lin.An optical fl ow-based composite navigation method inspired by insect vision.Acta Automatica Sinica,2015,41(6):1102−1112
(潘超,刘建国,李峻林.昆虫视觉启发的光流复合导航方法.自动化学报,2015,41(6):1102−1112)
