不同地理种群中缅树鼩MC1R基因的差异性研究
2018-03-08付家豪何一帆朱万龙高文荣王政昆
付家豪, 何一帆, 朱万龙, 高文荣, 王政昆
(云南省高校西南山地生态系统动植物生态适应进化及保护重点实验室云南师范大学 生命科学学院,昆明 650500)
哺乳动物的被毛颜色作为其主要的表型之一,主要用来掩饰自己避免天敌的捕捉,而这种性状的产生则是通过自身遗传多样性与环境因子共同作用导致的。有研究指出同一种或者同一种下的各亚种间的毛色都会产生一定的差异,例如方美英等对猪的毛色进行研究发现猪的毛色类型分为白色、黑色和棕色等[1]。这种表性的差异可以归结于“自然选择”的结果,即物种长期生存在某一环境下往往会产生对周围环境相对应的改变,这种改变最终会被固定下来反应在自身遗传多样性上。
动物的毛色主要是由黑色素(melanin)决定的,一般存在于皮肤的表层和眼睛的葡萄膜中。黑色素可以分为两种,一种叫真黑色素(eumelanin),另一种叫伪黑色素(pheomelanin),前者主要表现为黑色和褐色,而后者则主要表现为红色和黄色,二者的分布及比例共同影响了动物的毛色[2]。黑色素皮质激素受体1基因(Melanocortin receptor 1,Mc1r)作为控制动物毛色的主效基因[3],已在哺乳动物中得到验证[4-5]。它是一种最小的G蛋白偶联受体,隶属于G蛋白偶联家族,含有7个跨膜结构域,只有一个外显子长度约为900多bp,编码了310个左右的氨基酸,主要在黑色素细胞中表达造成色素沉积[6]。主要作用机制为:α-促黑色素皮质激素和促肾上素皮质激素作为Mc1r的配体与之结合,促使G蛋白二磷酸鸟苷活化为三磷酸鸟苷,激活腺苷酸活化酶通路生成环腺苷酸进一步活化络氨酸激酶促进黑色素细胞体内生成多巴,多巴聚集到一定程度释放黑色素[7]。如果活化的络氨酸酶过多,则多巴和多巴醌合成黑色素,倘若活化的络氨酸酶处于少量状态,则会出现伪黑色素[8]。
中缅树鼩(Tupaiabelangeri)隶属攀鼩目(Scandentia)树鼩科(Tupaiidae),为东洋界特有的一种小型哺乳动物[9]。其主要分布在南亚、东南亚以及我国的西南地区和海南岛,同样也是分布于我国的唯一一种现生树鼩。本研究旨在通过不同地理种群mc1r彼此间的序列差异及分子进化来探讨不同地理种群间的聚类关系,为以后研究中缅树鼩毛色的遗传机制提供一些基础性材料。
1 材料和方法
1.1 动物来源
实验所采用中缅树鼩(Tupaiabelangeri)捕捉于我国云南省、四川省、广西壮族自治区、贵州省以及海南省(表1)。捕捉后的动物经原地断颈处死,然后以干冰保存连夜送至云南师范大学生命科学学院动物生理生态实验室,用于后续DNA提取。
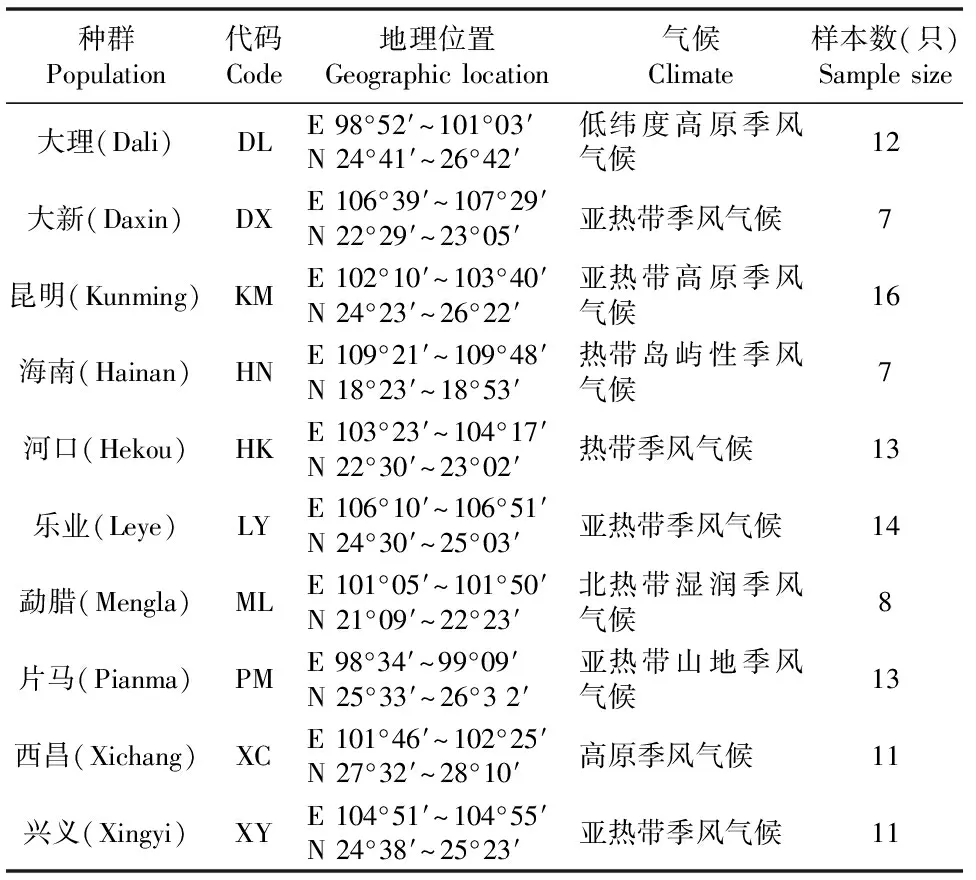
表1 中缅树鼩种群采样地点及样本数
1.2 DNA提取
取实验样品腿部肌肉组织,采用美国OMEGA公司肌肉组织DNA提取试剂盒(Tissue DNA Kit D3396-01)提取DNA,经1%琼脂糖凝胶电泳检验浓度后置于-20℃冰箱中保存。
1.3 MC1R基因扩增与测序
经NCBI查询中缅树鼩(Tupaiabelangri)黑色素皮质激素1(MC1R)基因(GenBank登录号:NW_006159850.1)CDS区碱基序列,设计引物覆盖整个CDS区(表2)。PCR扩增条件:94℃条件下预变性5 min;94℃变性30 s,60℃退火35 s,72℃延伸1 min,共35个循环;最后72℃延伸8 min。PCR产物经1%琼脂糖凝胶电泳检测后,送至上海生工生物工程股份有限公司测序。
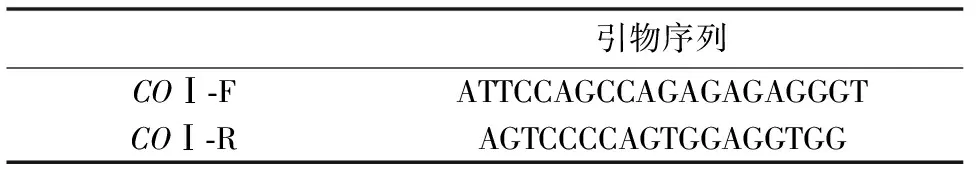
表2 Mc1r引物基本情况
1.4 数据分析与处理
采用Seqmen软件对测序结果进行拼接,获得954 bp基因序列,经BLAST比对确认所测序列为目的基因;采用Mega软件对112条序列进行比对并计算碱基含量及构建系统进化树;采用Dnasp软件计算单倍型及其在各地理种群中的分布并结合NetWork软件绘制单倍型网络进化图[10];通过I-TASSER在线服务[11-12](http://zhanglab.ccmb.med.umich.edu)和TMHMM Server v.2.0 在线软件进行蛋白质二级结构与跨膜结构域预测[13]。
2 结果与分析
2.1 Mc1r基因序列碱基组成
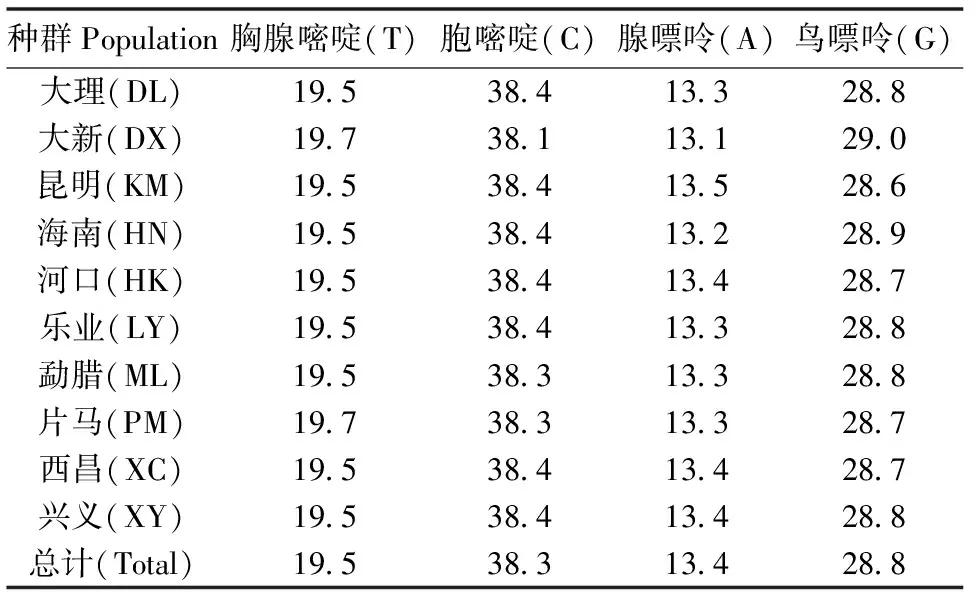
通过聚合酶链式反应(PCR)技术共扩增得到112个样本954 bp碱基序列(图1),其中各碱基平均组成为(表1):A(13.4%)、T(19.5%)、C(38.3%)和G(28.8%)。A+T(32.9%)比例明显低于C+G(67.1%),表现出程度较高的碱基含量偏倚性,G、C含量的过高,说明这段序列中富含氢键。另外一共在954 bp序列中检测到930个保守位点(Conserved sites),占总位点数的97.4%。另有24个变异位点(Variable sites),占总位点数的2.6%。
2.2 生物信息学分析
954 bp基因序列一共编码了317个20种常用氨基酸(表2),其中亮氨酸(Leu)含量最高,占18.2%,含量最少的为色氨酸(Trp),为0.9%。蛋白质二级结构预测表明,α-螺旋占了较大比例,其次是无规则卷曲(图2),并且含有7个跨膜结构域(图3)。
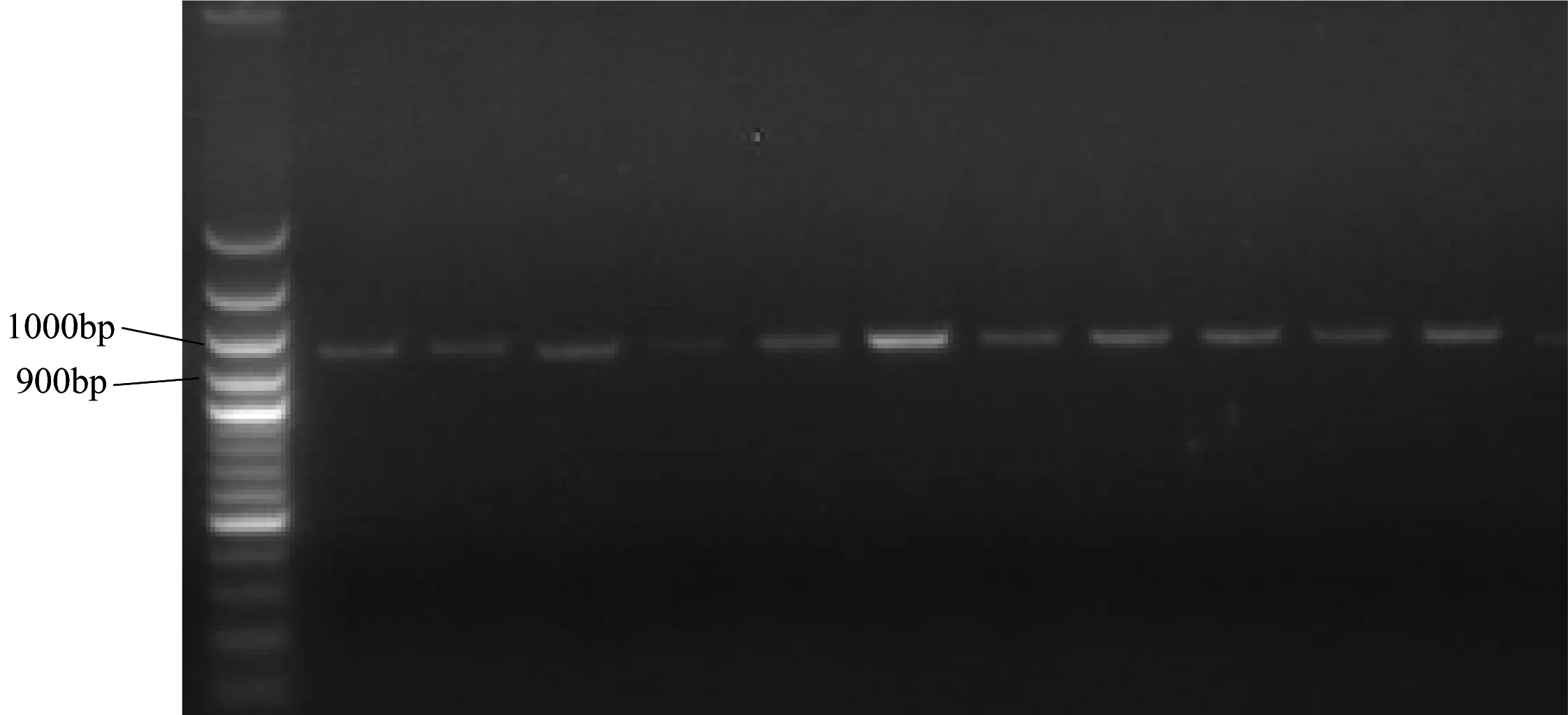
图1 PCR电泳结果(西昌种群)
种群Population胸腺嘧啶(T)胞嘧啶(C)腺嘌呤(A)鸟嘌呤(G)大理(DL)195384133288大新(DX)197381131290昆明(KM)195384135286海南(HN)195384132289河口(HK)195384134287乐业(LY)195384133288勐腊(ML)195383133288片马(PM)197383133287西昌(XC)195384134287兴义(XY)195384134288总计(Total)195383134288
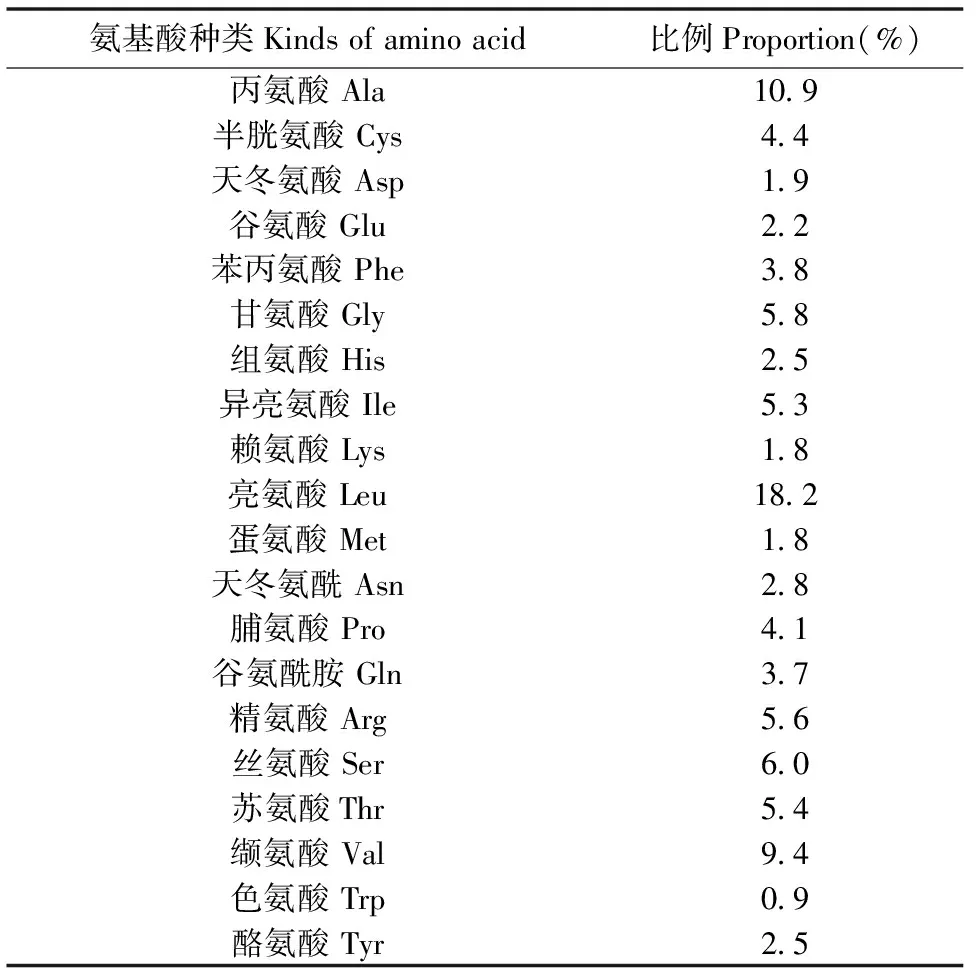
表4 Mc1r基因序列氨基酸组成
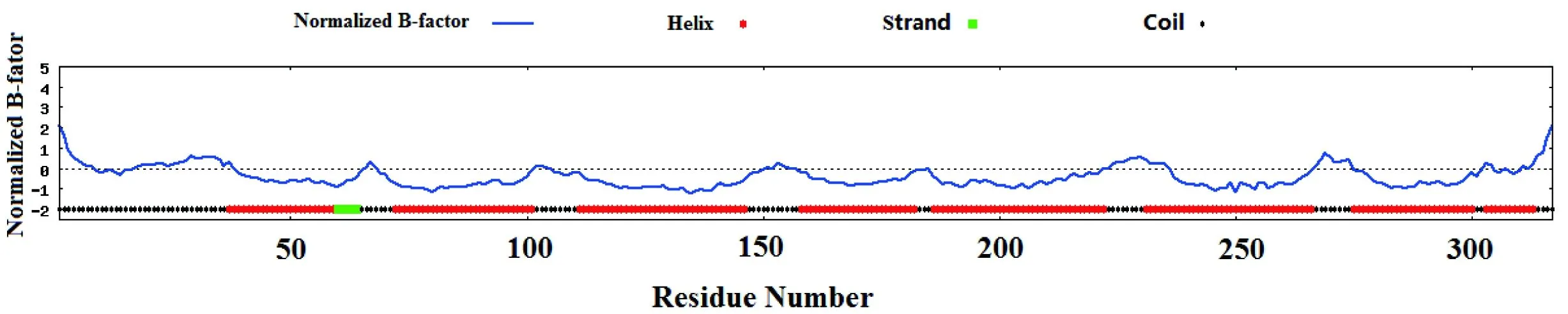
图2Mc1r翻译的蛋白质二级结构预测结果
Fig 2 The secondary structure ofmc1rtranslated protein
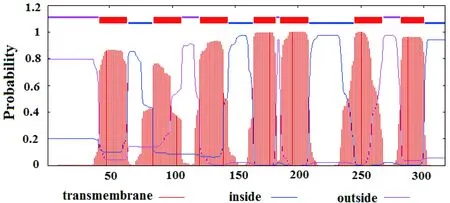
图3 Mc1r跨膜结构域预测
2.3 单倍型分析及种群间遗传距离
在本研究中112条mc1r序列共定义了13个单倍型,各单倍型在各地理种群中的分布如表3所示,其中有4个共享单倍型(H1、H2、H7、H9)占总单倍型数的30.7%。单倍型多样性指数介于0.125(昆明种群)~ 0.764(兴义种群)间,核苷酸多样性指数介于0.00013(昆明种群)~ 0.00122(兴义种群)间。10个地理种群MC1R基因种群间遗传距离结果显示(表4),大新种群、海南种群和片马种群与其他种群两两比较遗传距离值较大,介于0.008 ~ 0.012间,遗传距离最大的为海南种群与昆明种群。
2.4 MC1R基因序列饱和度检验及系统发育树构建
运用DAMBE软件对mc1r进行序列饱和度检验(Test of substitution saturation)(表5),如果ISS
3 讨论
在不同的生存环境中,物种为了适应环境从而形成相差异的遗传多样性,而遗传多样性往往也是研究环境和生物之间关系的重要内容[16]。中缅树鼩生存环境的不同,其被毛颜色也往往也不相同,表现为表型差异性。根据达尔文“自然选择”学说可把表型差异性归结为物种对周围生存环境长时间的适应的结果,并固定下来,最终反映在相应基因的遗传多样性上。于永梅等对野生型中缅树鼩进行表型毛色分析后发现,不同中缅树鼩被毛颜色表型上存在差异,但关于这些差异的遗传基础依旧不明朗[17]。在本研究成功扩增得到不同地理种群中缅树鼩Mc1r基因CDS区954 bp序列,共计编码317个氨基酸,其中各地理种群各氨基酸含量相差不显著。氨基酸二级结构作为整个蛋白质空间结构的基础,形成于蛋白质折叠的早期[18]。mc1r编码蛋白质二级结构以α螺旋为主,其次是无规则卷曲,其往往影响蛋白质三级结构以及亚细胞定位,并在细胞的信号转导和嘌呤和嘧啶的运输过程中发挥作用。10个地理种群中缅树鼩112个样本共定义到了13个单倍型,对这13个单倍型采用Kimura 2-Parameter距离模型构建ME系统进化树,结果显示13个单倍型大致分为4支:海南种群为CladeⅠ一支(Hap5、Hap6),大新种群为一支CladeⅡ(Hap3、Hap4),片马种群为一支CladeⅢ(Hap11),其他种群聚为一支CladeⅣ。
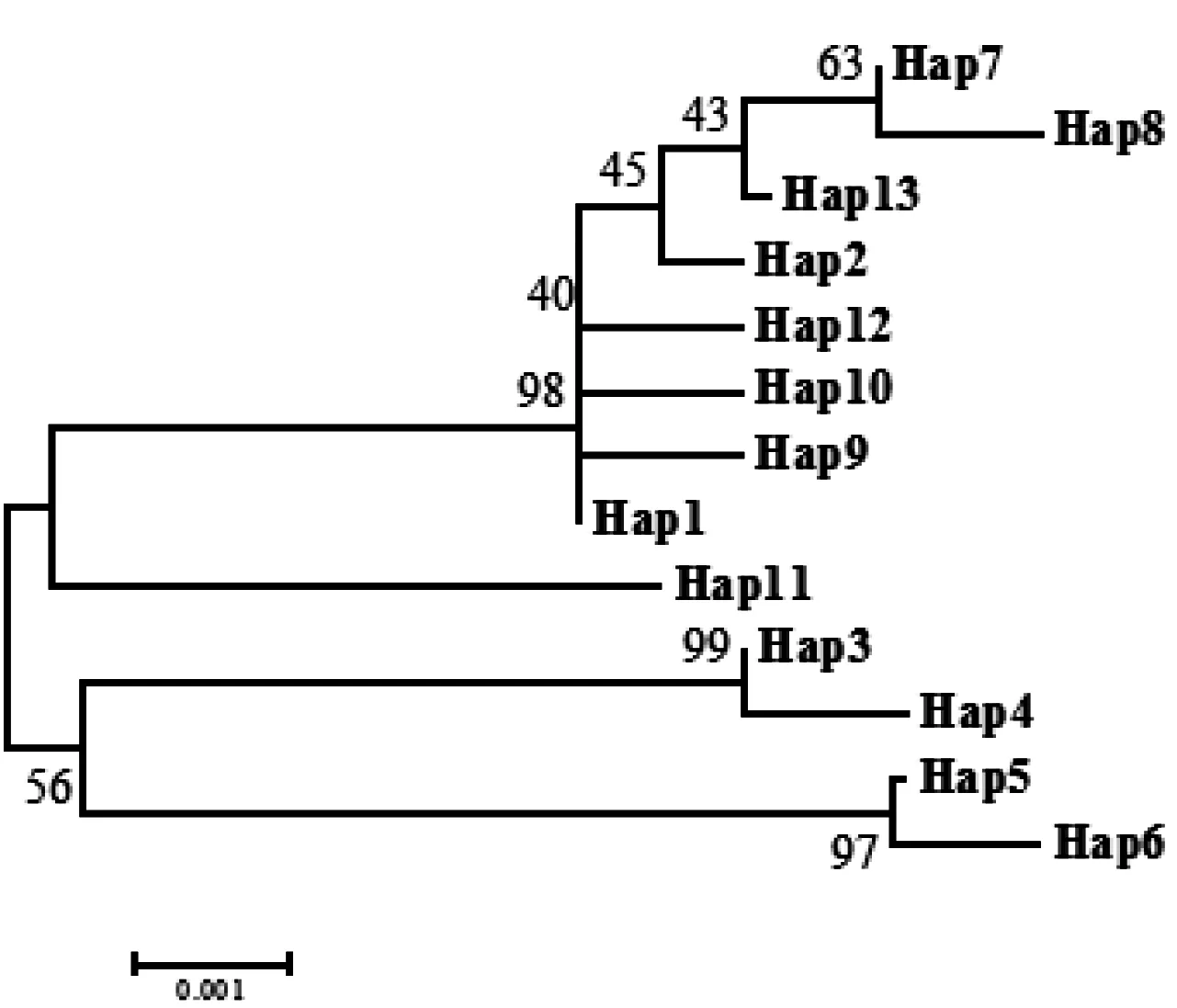
图4 基于Kimura 2-Parameter 距离构建的不同单倍体间的ME系统发育树(1000次重复)
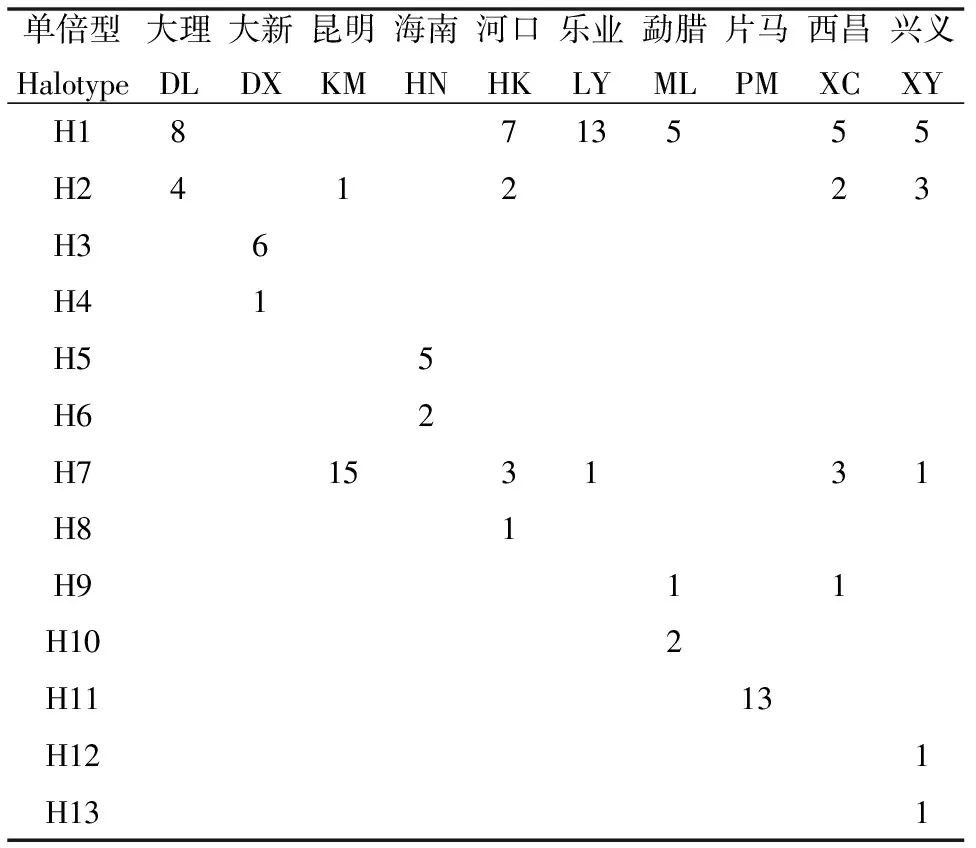
单倍型Halotype大理DL大新DX昆明KM海南HN河口HK乐业LY勐腊ML片马PM西昌XC兴义XYH18713555H241223H36H41H55H62H7153131H81H911H102H1113H121H131

表6 中缅树鼩种群间的遗传距离
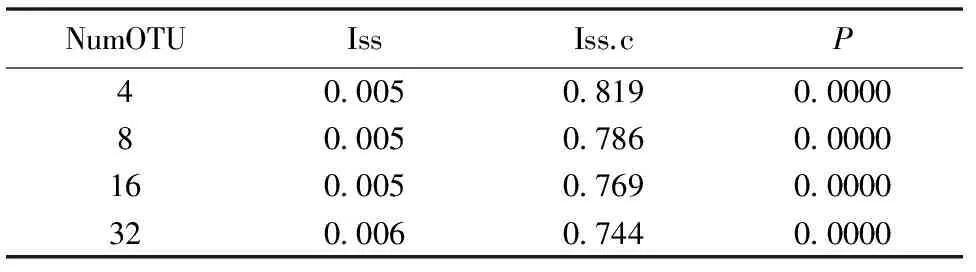
表7 Mc1r基因序列饱和度检验
基于以上实验结果得出结论,各地理种群单倍型虽然有所混杂,但在整体地理分布上趋于各自特有的地理格局,采样地地形整体方面表现为海拔由西到东逐渐降低,片马种群位于高黎贡山上即横断山脉南延部分,海拔在2500 m以上,年平均温度在15℃左右,并且右侧有怒江的阻隔作用,难以与其他种群进行基因交流。而海南种群位于海南岛上,处于典型的热带岛屿性季风气候,并且年平均温度稳定在25℃左右。其作为我国地理位置相对独立的一个省份,海南岛与广州的雷州半岛以及广西壮族自治区之间分别有琼州海峡和北部湾的阻隔作用,因此也难以与其他种群进行交流。而大新种群所处地理位置海拔较低,周围多为孤立的山峰形成屏障隔离难以与其他种群进行基因交流。其他种群多生存于云贵高原及其周边地区,由人为及其他因素原因影响导致种群间相对交流频繁,难以各自分开。气温作为影响生物物种进化的重要因素,其中的温度和降水量两个生态因子往往能影响陆生物种的进化[19],被毛颜色作为动物抵御天敌捕食的最好伪装,由多个因素来调控,基因调控只是一方面。有研究表明,合成真黑色素与伪黑色素的酶系对温度比较敏感[20],因此由于各地理环境的不同而表现出差异。
综上所述,10个地理种群中缅树鼩聚为四大类群,mc1r序列差异可能是由环境差异引起的。本研究主要从不同地理种群间中缅树鼩mc1r序列差异上来分析,为下一步工作从基因表达量方面探讨毛色的差异是如何形成的奠定了基础。
[1]方美英,吴常信. 猪的毛色遗传[J]. 中国畜牧杂志, 2002, 38(2):51-52.
[2]CONE R D, LU D, KOPPULA S, et al. The melanocortin receptors: agonists, antagonists, and the hormonal control of pigmentation [J]. Recent Progress in Hormone Research, 1996, 51 (7): 287-317.
[4]HUNT G, ROT HUIZEN J, VAN B A. Indemnification of 4 premature stop codon in the elanocyte mrnulating hormone receptor gene(MClR)in labrador and Golden rot rievers with yellow coat color [J]. Atom Genet, 2000, 31 (3): 194-199.
[5]LIBOUTET M, PORTELA M, DELESTAING G, et al. MC1R and PTCH gene polymorphism in French patients with basal cell carcinomas [J]. Digest of World Core Medical Journals, 2006, 126 (7): 1510-1517.
[6]王 乐, 吕雪峰, 郑文新,等. 动物毛色形成的原因和机制 [J]. 草食家畜, 2009(1): 20-21.
[7]SUZUKI I, CONE R D, IM S, et al. Binding of melanotropic hormones to the melanocortin receptor MC1R on human melanocytes stimulates proliferation and melanogenesis[J]. Endocrinology, 1996, 137 (5): 1627-1633.
[8]甘海云, 李建斌, 王洪梅,等. 牛毛色基因的研究进展[J]. 中国草食动物科学, 2007, 27 (1): 50-52.
[9]王政昆, 高文荣, 朱万龙. 横断山区三种小型哺乳动物代谢特征的研究进展[J]. 兽类学报, 2015, 35 (4): 445-452.
[10]LIBRADO P, ROZAS J. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data[J]. Bioinformatics, 2009, 25(11): 1451-1452.
[11]YANG J, YAN R, ROY A, et al. The I-TASSER suite: Protein structure and function prediction[J]. Nature Methods, 2015, 12(1):7-8.
[12]ROY A, KUCUKURAL A, ZHANG Y. I-TASSER: a unified platform for automated protein structure and function prediction [J]. Nature Protocols, 2010, 5(4): 725-738.
[13]SONNHAMMER E L, VON HEIJINE G, KROGH A. A hidden Markov model for predicting transmembrane helices in protein sequences[J].Proc Int Conf Intell Syst Mol Biol, 1998, 6: 175-182.
[14]XIA X, XIE Z, SALEMI M, et al. An index of substitution saturation and its application[J]. Molecular Phylogenetics & Evolution, 2003, 26 (1): 1-7.
[15]XIA X, LEMEY P. The phylogenetic handbook: a practical approach to DNA and protein phylogeny[M]. Cambridge: Cambridge University Press,2009: 611-626.
[16]MERRELL D J. Ecological genetics[M]. London: Longman,1981:3-12.
[17]于永梅, 吴一迁, 钱耕荪,等. 中缅树鼩形态性状的遗传制约性初步研究[J]. 遗传, 2004, 26 (4): 478-480.
[18]连云涓. 蛋白质二级结构预测的多核学习方法[D]. 上海:上海交通大学, 2013.
[19]孙儒泳. 动物生态学原理[M]. 北京:北京师范大学出版社, 2001:18-20.
[20]张俊珍, 董常生, 范瑞文,等. 哺乳动物毛色形成研究进展[J]. 动物医学进展, 2006, 27 (增): 65-68.
