柏木低效林不同改造模式优势草本植物多样性及其生态位
2018-03-08陈丝露李贤伟肖宝茹
陈丝露, 赵 敏, 李贤伟, 范 川,*, 肖宝茹
1 四川农业大学林学院,长江上游林业生态工程四川省重点实验室, 成都 611130 2 德阳市林业局, 德阳 618000
生态位自提出后[1],众多学者对于其理论[2-3]、测度及应用[4-7]进行了大量的研究,生态位宽度可以度量物种对环境资源的利用情况与其在环境中所处地位,生态位重叠能反映生态位分化状况,生态位分化指两个物种对某种或多种环境资源利用的差异,这种差异可以使两个物种形成平衡而共存,基于生态位分化的物种共存理论是群落生物多样性维持的重要机制之一[8],在群落构建中具有重要的作用[9]。生态位现已被广泛应用于评价种间关系、群落结构、生物多样性及演替等方面[10],如对不同类型的森林群落[11-13]、灌木或草本群落[14-16]的优势种群的生态位进行研究,可以揭示群落中物种间的关系和群落演替状况;研究濒危植物的生态位[17-18]能指导植物保护措施;对恢复过程中植物群落[19-20]生态位变化进行研究,可预测未来群落植物发展趋势等。这些研究大多涉及到自然状态下的植物群落或人工林植物群落的生态位,但有关人工林改造的群落生态位变化研究仍不多见[21-22]。柏木(Cupressusfunebris)林是四川盆地丘陵区主要的森林类型,也是主要的低产低效林分[23-24],属于我国亚热带常绿针叶林之一。由于初植密度过高,树种单一,柏木林下植被结构简单,生物多样性低,而草本层相对于灌木层和乔木层具有更高的敏感度,在改造初期能更快反映改造带来的影响[25-26],并且在促进人工林养分循环和维护森林立地生产力等方面有重要的作用[27]。本文通过分析不同改造模式的柏木林的优势草本植物多样性及生态位,揭示人工林经改造后草本群落的地位、作用、多样性等变化,以最直观的草本植物组成变化推测林分各因子的变化,预测短期内草本植物群落的发展趋势,确定适宜的改造模式,以期为柏木低效林分改造提供理论依据和指导。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于四川盆地中部,成都平原东北边缘的德阳市旌阳区(104°15′—104°32′ E,31°05′—31°20′ N)。该区为低山丘陵地貌,海拔457—764 m,属中亚热带湿润季风气候区,年平均气温16.1℃,最高气温36.5℃,最低气温-6.7℃,无霜期271 d,日照时间1251.4 h,年均降雨量893.4 mm,集中在7—8月,该地区林地土壤类型为紫色土且侵蚀严重,土壤贫瘠,呈中性至微碱性。柏木低效林改造前生境质量较差,树种单一,林下植被稀少,生物多样性低。2011年(改造前)柏木林平均胸径为7 cm,平均高度8 m,郁闭度0.8,林下灌木主要为铁仔(MyrsineAfricana)、黄荆(Vitexnegundo),草本主要为薹草(Carextristachya)、竹叶草(Oplismenuscompositus)、铁线蕨(Adiantumcapillus-veneris),林下物种稀少,多样性低,生境质量差。2011年实施改造,形成了以下模式:高密度桤木(QMⅠ)、香椿(XCⅠ)、银木(YMⅠ)模式(株行距1.5 m×2 m)、低密度桤木(QMⅡ)、香椿(XCⅡ)、银木(YMⅡ)模式(株行距1.5 m×4 m)、香椿+银木(XY)、银木+桤木(YQ)、香椿+桤木(XQ)、桤木+香椿+银木(QXY)模式(株行距均为1.5 m×2 m)和采伐迹地(CF),其中银木+桤木、香椿+银木、香椿+桤木模式混交比例1∶1,桤木+香椿+银木模式则为1∶1∶1,各模式采用行状混交,柏木纯林(CB)为对照。试验地基本情况见表1。

表1 试验地植被概况Table 1 Vegetation characteristics of experimental plots
QMⅠ:高密度桤木,high densityAlnuscremastogyne;XCⅠ:高密度香椿,high densityToonasinensis;YMⅠ:高密度银木,high densityCinnamomumseptentrionale;XY: 香椿+银木,Toonasinensis+Cinnamomumseptentrionale;YQ:银木+桤木,Cinnamomumseptentrionale+Alnuscremastogyne;XQ:香椿+桤木,Toonasinensis+Alnuscremastogyne;QXY:桤木+ 香椿+银木,Alnuscremastogyne+Toonasinensis+Cinnamomumseptentrionale;QMⅡ:桤木,low densityAlnuscremastogyne;XCⅡ:低密度香椿,low densityToonasinensis;YMⅡ:低密度银木,low densityCinnamomumseptentrionale;CF:采伐迹地,cutting-blank;CB:柏木,Cupressusfunebris
1.2 研究方法
1.2.1 调查方法
于2015年8月在每个模式设置4个面积为10 m×10 m的样方,样方内的四角和中心随机设置5个1 m×1 m的固定小样方来调查草本植物,草本样方共240 个,并记录样方内草本植物的种类、数量、均高、盖度等。计算各草本重要值,在重要值的基础上结合各物种在样方中的盖度及其在样方中出现的频率确定16 个主要物种进行生态位分析。
1.2.2 计测方法
(1)生物多样性
本文主要采用以下α多样性相关指数[28-29]:
1)多样性指数采用Shannon-wiener(H)和Simpson(H′)指数计算:
2)均匀度指数Pielou指数:
3)物种丰富度指数R:
R=(S-1)/ln(N)
式中,S为样方中出现的物种数;N为物种总数量;pi=ni/N;ni为第i个种的个体数目。
(2)重要值
重要值=(相对密度+相对频度+相对盖度)/ 3
(3)生态位宽度
生态位宽度采用Shannon-Wiener生态位宽度指数[30-32]:
式中,Bi为i物种的生态位宽度;Pij=nij/Ni,式中nij为物种i在资源j上的重要值,Ni表示物种i在所有资源上的重要值之和。Pij代表物种i在资源j上的重要值占该种在所有资源上的重要值比例,r为资源位总位数,j指不同改造模式。
(4)生态位重叠
采用Levins[11,33]生态位重叠公式:

1.3 统计分析
应用Excel 2010、SPSS 20.0和GraphPad Prism 5对实验数据进行整理和分析,柏木低效林不同改造模式间的多样性的差异性采用单因素方差分析(one-way ANOVA)。
2 结果与分析
2.1 柏木低效林不同改造模式间草本植物多样性

图1 柏木低效林不同密度纯林模式间草本植物多样性Fig.1 The plant diversity of herbs between different density reconstructing patterns of low efficiency stands of Cuprssus funebris不同小写字母表示草本植物多样性在不同密度模式间差异显著(P<0.05)
由图1可知,不同密度的纯林模式间草本植物多样性表现为,高密度桤木模式草本植物Shannon-Wiener指数(1.59)、丰富度指数(1.29)均显著小于低密度桤木模式(1.69,1.55),均匀度指数(0.89)大于低密度桤木模式(0.81)但不显著,Simpson指数相差不大。高密度香椿模式草本植物的Shannon-Wiener指数(1.30)、丰富度指数(1.15)均显著大于低密度香椿模式(1.19,0.66),而均匀度指数(0.72)显著小于低密度香椿模式(0.86),Simpson指数为两者相同。高密度银木模式草本植物的丰富度指数(1.58)显著大于低密度银木模式(0.85),而均匀度指数(0.75)显著小于低密度银木模式(0.95),Shannon-Wiener指数(1.56)与Simpson指数(0.80)为高密度大于低密度(1.48,0.76)但不显著。由此可以看出,银木和香椿模式的草本植物多样性表现为高密度优于低密度,桤木模式的草本植物多样性表现为低密度优于高密度。在纯林模式中,香椿模式对于草本植物多样性改造的效果较其他模式差,银木模式次之,桤木模式最好。
从图2可以看出,高密度不同树种模式的草本植物Shannon-Wiener多样性指数为:香椿+桤木模式(1.83)和香椿+银木模式(1.82)显著高于其他模式;银木+桤木模式(1.59)、高密度桤木模式(1.59)、高密度银木模式(1.56)与采伐迹地(1.54)次之,但与柏木纯林(1.48)差异不显著;桤木+香椿+银木模式(1.44)低于柏木纯林但差异不显著;高密度香椿模式(1.30)显著低于柏木纯林(1.48)。关于Simpson指数:只有香椿+桤木模式(0.83)和香椿+银木模式(0.83)显著高于柏木纯林;高密度银木模式(0.80)、银木+桤木模式(0.77)、高密度桤木模式(0.77)、采伐迹地(0.73)及桤木+香椿+银木模式(0.72)高于柏木纯林(0.69)但差异不显著;高密度香椿模式(0.65)低于柏木纯林但差异不显著。在均匀度指数方面:香椿+桤木模式(0.93)、香椿+银木模式(0.94)、高密度桤木模式(0.89)、银木+桤木模式(0.88)均大于柏木纯林(0.82),而桤木+香椿+银木模式(0.81)、采伐迹地(0.79)、高密度银木模式(0.75)及高密度香椿模式(0.72)均小于柏木纯林,但所有模式均与柏木纯林差异不显著;从丰富度指数可以看出:高密度银木模式(1.58)显著高于其他模式;香椿+桤木模式(1.33)、香椿+银木模式(1.30)、采伐迹地(1.30)、高密度桤木模式(1.29)高于柏木纯林(1.27)但不显著;高密度香椿模式(1.15)低于柏木纯林但不显著;桤木+香椿+银木模式(1.12)和银木+桤木模式(1.08)显著低于柏木纯林。由此可看出,本研究中最适宜的改造模式为香椿+银木模式和香椿+桤木模式和高密度桤木模式,桤木+香椿+银木模式与高密度香椿模式改造效果较差。

图2 柏木低效林不同树种模式间草本植物多样性Fig.2 The plant diversity of herbs between different tree species reconstructing patterns of low efficiency stands of Cupressus funebris不同小写字母表示不同模式间草本植物多样性差异显著(P<0.05)
2.2 柏木低效林不同改造模式间草本植物生态位宽度
重要值反应物种的优势度,如表2所示,总重要值较大的物种有薹草(18.244)、竹叶草(16.124)、地果(Ficustikoua)(14.210)、五节芒(Miscanthusfloridulus)(13.056),它们在各样地中的重要值也相对较大,说明这些物种在群落中占优势,为草本群落中的优势种。而蛇莓(Duchesneaindica)(10.068)、铁线蕨(4.034)、马兰(Kalimerisindica)(3.904)、中日金星蕨(Parathelypterisnipponica)(3.578)、三裂蛇葡萄(Ampelopsisdelavayana)(3.123)、蜈蚣草(Pterisvittata)(2.609)、序叶苎麻(Boehmeria clidemioides)(1.816)、渐尖毛蕨(Cyclosorusacuminatus)(1.811)、茼蒿(Chrysanthemumcoronarium)(1.412)、井栏边草(Pterismultifida)(1.396)、鼠麴草(1.308)、牡蒿(Artemisiajaponica)(1.305)重要值较小,依次位于群落的下层。在各模式中,薹草在高密度桤木模式(30.603)、香椿+银木(24.241)、香椿+桤木模式(28.773)、桤木+香椿+银木模式(32.746)中重要值最大;竹叶草在高密度银木模式(23.685)、高密度香椿模式(31.620)、银木+桤木模式(21.866)、采伐迹地(30.782)中最大;五节芒在低密度桤木、香椿、银木模式中最大(29.606、28.939、31.546);铁线蕨在柏木纯林(25.040)中最大,以上物种在不同的模式中分别占据了最大的重要值,相对与其他物种更有生长优势。
生态位宽度是度量种群对环境资源利用的尺度,可以表达种群在群落中的地位和作用。由表3可看出总生态位宽度从大到小分别为薹草(2.276)、竹叶草(2.239)、地果(2.255)、蛇莓(2.152)、五节芒(1.745)、马兰(1.508)、中日金星蕨(1.375)、三裂蛇葡萄(1.361)、铁线蕨(1.093)、序叶苎麻(1.091)、蜈蚣草(1.030)、渐尖毛蕨(0.693)、茼蒿(0.679)、井栏边草(0.656)、鼠麴草(0.647)、牡蒿(0.614)。与重要值的测算结果大致相同,说明重要值大的物种一般具有较宽的生态位,重要值小的物种一般则具有较小的生态位。薹草、竹叶草、地果在10个资源位中有分布,是草本群落的优势种,数量多,生态幅较大,对资源利用能力强,故分布较其他物种广。而渐尖毛蕨、茼蒿、井栏边草等物种只在少数资源位有分布,对资源利用能力不强,生态适应范围较窄,分布不均,生态位宽度较小。
表2柏木低效林不同改造模式草本优势种群重要值
Table2TheimportancevalueofherbaceousdominantpopulationsofthedifferentreconstructingpatternsoflowefficiencystandsofCupressusfunebris
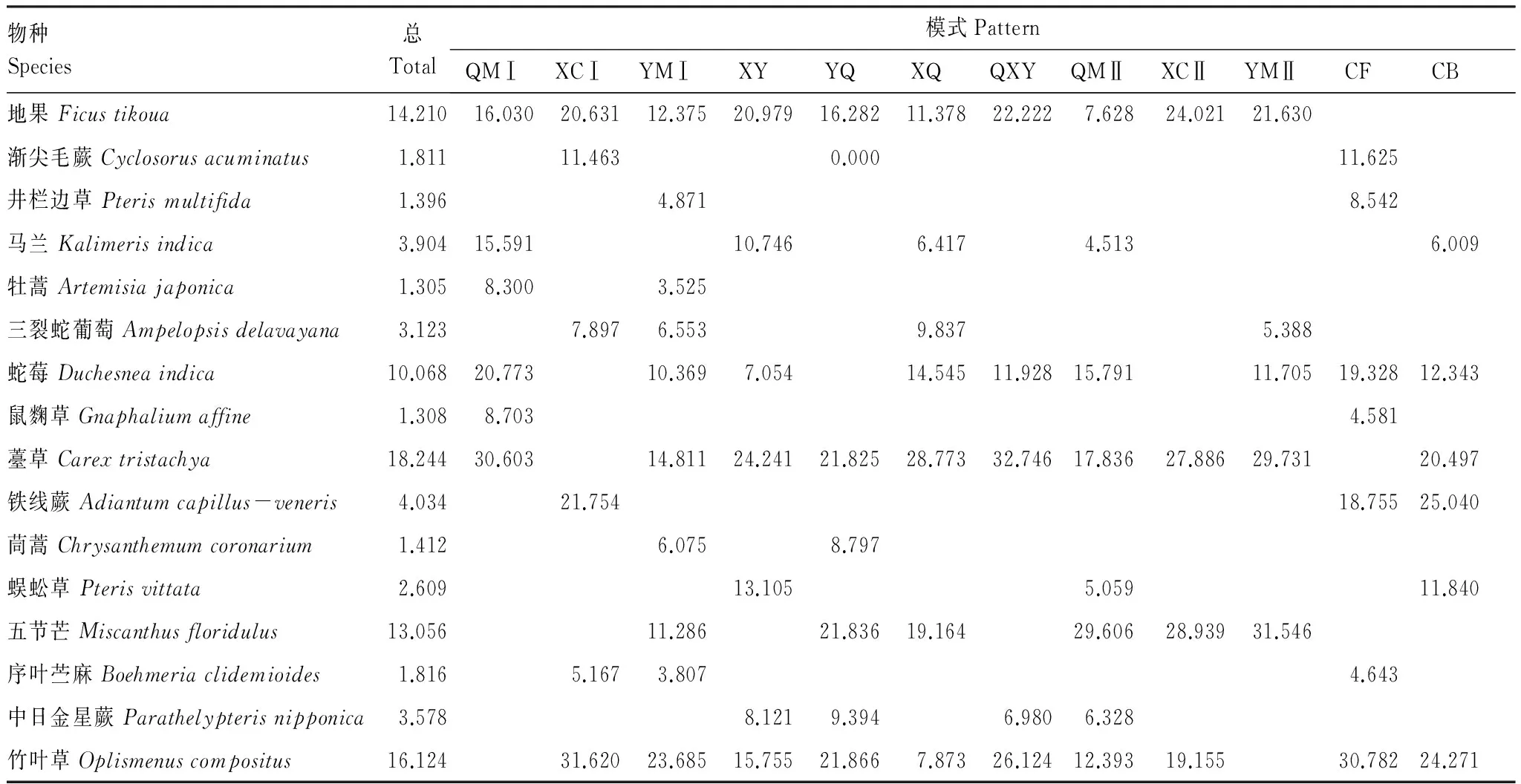
物种Species总Total模式PatternQMⅠXCⅠYMⅠXYYQXQQXYQMⅡXCⅡYMⅡCFCB地果Ficustikoua14.21016.03020.63112.37520.97916.28211.37822.2227.62824.02121.630渐尖毛蕨Cyclosorusacuminatus1.81111.4630.00011.625井栏边草Pterismultifida1.3964.8718.542马兰Kalimerisindica3.90415.59110.7466.4174.5136.009牡蒿Artemisiajaponica1.3058.3003.525三裂蛇葡萄Ampelopsisdelavayana3.1237.8976.5539.8375.388蛇莓Duchesneaindica10.06820.77310.3697.05414.54511.92815.79111.70519.32812.343鼠麴草Gnaphaliumaffine1.3088.7034.581薹草Carextristachya18.24430.60314.81124.24121.82528.77332.74617.83627.88629.73120.497铁线蕨Adiantumcapillus-veneris4.03421.75418.75525.040茼蒿Chrysanthemumcoronarium1.4126.0758.797蜈蚣草Pterisvittata2.60913.1055.05911.840五节芒Miscanthusfloridulus13.05611.28621.83619.16429.60628.93931.546序叶苎麻Boehmeriaclidemioides1.8165.1673.8074.643中日金星蕨Parathelypterisnipponica3.5788.1219.3946.9806.328竹叶草Oplismenuscompositus16.12431.62023.68515.75521.8667.87326.12412.39319.15530.78224.271
表3柏木低效林不同改造模式草本优势种群的生态位宽度
Table3ThenichebreadthofherbaceousdominantpopulationsofdifferentreconstructingpatternsoflowefficiencystandsofCupressusfunebris

物种Species总Total模式PatternQMⅠXCⅠYMⅠXYYQXQQXYQMⅡXCⅡYMⅡCFCB地果Ficustikoua2.2551.7362.0561.3132.2412.0421.3562.1151.0162.2511.919渐尖毛蕨Cyclosorusacuminatus0.6931.5601.734井栏边草Pterismultifida0.6561.0891.003马兰Kalimerisindica1.5082.0481.7601.0061.0741.098牡蒿Artemisiajaponica0.6141.5810.693三裂蛇葡萄Ampelopsisdelavayana1.3611.0701.0711.0901.081蛇莓Duchesneaindica2.1522.1681.5981.0421.3701.3341.7131.3052.2211.715鼠麴草Gnaphaliumaffine0.6471.7781.077薹草Carextristachya2.2762.4541.7632.3742.2822.0562.4492.1362.5132.3642.174铁线蕨Adiantumcapillus-veneris1.0932.2882.0472.365茼蒿Chrysanthemumcoronarium0.6791.3502.020蜈蚣草Pterisvittata1.0301.7191.0601.763五节芒Miscanthusfloridulus1.7451.3822.1072.3532.2502.3352.456序叶苎麻Boehmeriaclidemioides1.0911.0440.6811.076中日金星蕨Parathelypterisnipponica1.3751.0041.3781.0651.097竹叶草Oplismenuscompositus2.2392.3682.1901.8812.4231.0912.1381.0622.2482.3692.070
2.3 柏木低效林不同改造模式间草本植物生态位重叠
由表4可看出,草本物种之间的生态位重叠值均较小,生态位重叠最大值0.123,为鼠麴草和蛇莓,其次为渐尖毛蕨和竹叶草(0.120),说明它们对环境的适应性和对资源的利用能力具有较大的相似性。生态位宽度较大的物种间既有较大的生态位重叠值,如薹草与地果的Lih(0.078)和Lhi(0.081)、薹草与蛇莓的Lih(0.065)和Lhi(0.074);也有较小的生态位重叠值,如地果与蛇莓的Lih(0.055)和Lhi(0.049)、薹草与竹叶草的Lih(0.054)和Lhi(0.054),说明生态位宽度的大小与生态位重叠没有明显的规律。生态位重叠为0的有56对,占总数的23.33%;0—0.02的有44对,占总数的18.33%;0.02—0.04的有60对,占总数的25%;0.04—0.06的有33对,占总数的13.75%;0.06—0.08的有27对,占总数的11.25%;0.08—0.1的有14对,占总数的5.83%;大于0.1的有6对,占总数的2.5%[34]。表明草本层只存在低度生态位重叠,各物种对资源的利用情况相似度低,竞争并不激烈。

表4 柏木低效林不同改造模式草本优势种生态位重叠Table 4 The niche overlap of herbaceous dominant populations of different reconstructing patterns of low efficiency stands of Cupressus funebris
2.4 柏木低效林不同改造模式间生态位宽度与重要值和资源位的关系
由图3可看出,柏木低效林不同改造模式间草本植物生态位宽度与重要值呈极显著的正相关关系,相关系数为0.916,说明重要值较大的物种一般具有较大的生态位宽度。由图4可知,柏木低效林不同改造模式间生态位宽度与资源位也呈极显著的正相关关系,相关系数为0.977,资源位多的物种往往具有较大的生态位宽度,如草本群落中如五节芒(13.056)比蛇莓(10.068)的重要值大,生态位宽度(1.745)却小于蛇莓(2.152),是由于蛇莓在9个资源位中均有分布而五节芒只在6个资源位分布。总的来说生态位宽度与重要值和资源位的关系都十分密切。

图3 柏木低效林不同改造模式草本植物生态位宽度与重要值相关性Fig.3 Correlation between the niche breadths and importance value of the different reconstructing patterns of low efficiency stands of Cupressus funebris**表示相关性系数达到极显著水平(P<0.01)
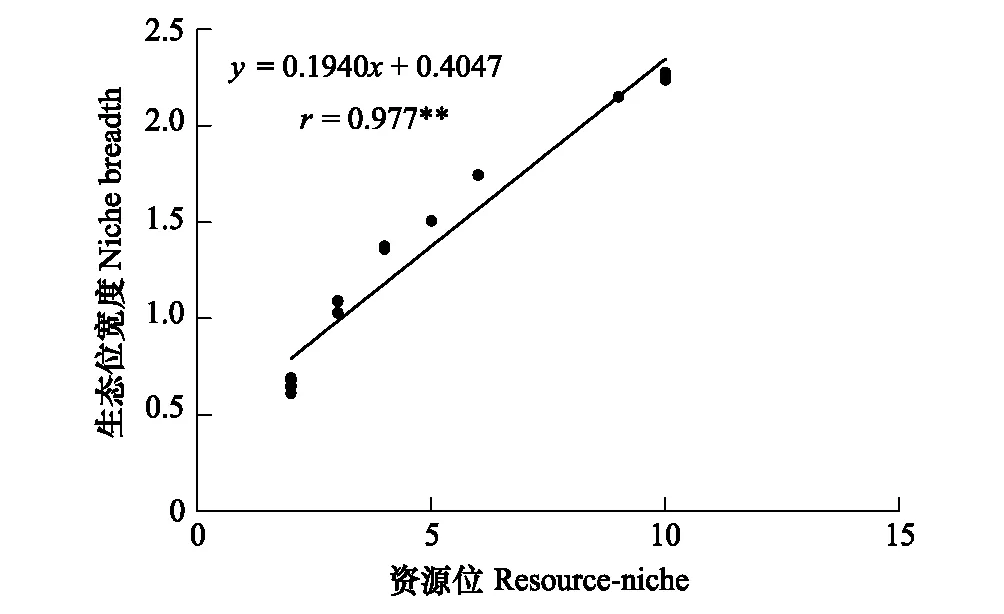
图4 柏木低效林不同改造模式草本植物生态位宽度与资源位相关性Fig.4 Correlation between the niche breadths and resource-niche of the different reconstructing patterns of low efficiency stands of Cupressus funebris**表示相关性系数达到极显著水平(P<0.01)
3 讨论
3.1 改造对草本植物多样性的影响
生物多样性的保育是森林经营的重点[35],本研究采取的模式中香椿+桤木、香椿+银木模式显著的提高了柏木纯林的林下草本植物的生物多样性,桤木+香椿+银木模式、高低密度香椿模式与低密度银木模式改造效果较差。对于银木和香椿模式来说,高种植密度的改造效果比低种植密度好,而对于桤木来说,低种植密度的改造效果比高种植密度好,造成这种现象的原因可能是不同树种对种植密度的适应性不同[36]。混交林改造模式比纯林改造模式效果好[37],但混交度较大的桤木+香椿+银木模式林下草本植物多样性低于两种树种的混交模式,原因可能是虽然混交度越高,一般意味着群落越稳定,但多数研究认为其基于成熟的林分[38],而本研究中的各模式均处于幼龄林阶段,草本物种组成不稳定,具有多变性[39]。
改造会引起林分物理环境和生物因子的改变,这些因素的改变往往和植物多样性有关。引起草本层植物多样性变化的主要原因之一是林下光照资源的改变[40]。改造降低了乔木层郁闭度,增加了林下植被的光照资源,在郁闭度最低的低密度香椿模式和银木模式中,喜阳植物五节芒大量繁殖,占据了林下生长空间,生物多样性低,高密度模式的郁闭度相对较高,种间光资源竞争增强,独占生长资源的喜阳植物五节芒逐渐被淘汰,喜阴植物开始处于优势地位,在郁闭度最高的香椿+桤木模式和香椿+银木模式的草本植物多样性最大。在采伐迹地中,由于郁闭度的大幅度降低且没有其他乔木树种的竞争,盐肤木迅速生长居于优势地位,占有大量的光照资源,林下草本多为耐阴植物,多样性不高。由此可见,林下草本层多样性与林木郁闭度有密切关系,郁闭度过低,喜阳植物会占据大量生长资源,植物多样性较低,随郁闭度增加,植物多样性逐渐增大,当喜阳和喜阴植物达到平衡时植物多样性达到最大[41]。林分的改造不仅降低了乔木层的郁闭度,也降低了乔木层的林冠截流,有利于林分涵养水源[42],许多研究发现改造后的林分凋落物、土壤的持水能力及肥力都有一定程度的提升[23],而土壤水分和养分的改变对植物多样性的变化有较大的影响[43],本研究中除香椿模式和低密度银木模式改造效果低于柏木纯林外,其他模式均不同程度的优于柏木模式,可以推测改造后土壤水分及养分条件的改变导致了不同模式间植物多样性的改变,香椿+桤木模式和香椿+银木模式的土壤条件的改变最利于草本植物多样性的增加,其他模式次之,而香椿模式、低密度银木模式的草本植物多样性反而低于柏木纯林,可能是因为这几种改造模式的郁闭度过低,过度的降低了林冠截流,导致降雨时形成地表径流,直接冲刷并侵蚀了土壤[42],造成土壤养分的流失,从而导致草本植物多样性降低。除了光照和土壤因素外,生物因子也是改变植物多样性的原因之一。改造后,除香椿模式与低密度银木模式外,其他模式的灌草植物种类增加,林下结构较柏木纯林更为复杂,而桤木和香椿为落叶乔木,能产生更多的地表枯落物,加之改造林分透光度增加,雨水更易穿透林层到达地面,温度和水分的改善使凋落物更容易腐殖化,有利于土壤微生物的生长[43]。
香椿+桤木和香椿+银木模式能较快的进行演替,为林下草本植物提供最适宜的光照环境,又不至于导致阳生植物迅速生长,能保持较高的生物多样性,是较为理想的改造模式。关于土壤因子及生物因子,具体情况需做进一步的研究,但草本植物多样性的增加无疑预示着草本群落的生存条件的改善。
3.2 改造对草本植物生态位宽度的影响
物种的生态位宽度越大,表明物种具有更强的资源利用能力,更高的生态适应性和更广的分布幅度[11]。改造前生态位宽度最广的是铁线蕨,说明其在柏木纯林中具有较强的适应性,可利用资源幅度广。改造后由于郁闭度降低,喜阳植物五节芒不断拓展生态位宽度,在低密度纯林模式中五节芒的生态位宽度高于其他物种,资源利用能力较强,抑制了其他植物生长。随改造模式的郁闭度增加,种间光资源竞争增强,独占生长资源的喜阳植物五节芒逐渐被淘汰,喜阴植物逐渐占据优势地位,生态位宽度不断变大,薹草、地果、竹叶草等占据了林下生长空间,成为草本层的优势种。不同物种的生态位宽度的此消彼长反应了群落演替的规律[19],改造打破了柏木纯林原有的林下植被结构,为其他物种提供了有利的生存条件[44],改善了草本群落的结构组成。Brown认为生态位宽度较大的物种能利用更多的资源,能在更大范围内优势生长[45],也有很多学者认为生态位宽度大的物种不一定有较大的生长范围[46-48],在本研究中草本植物重要值与生态位宽度呈极显著的正相关关系(r=0.916),重要值较大的物种具有大的生态位宽度,分布幅度更广;资源位也与生态位宽度呈极显著正相关关系(r=0.977),说明物种所占的资源位数越多,生态位宽度则越大,这与胡正华等[11]、钱逸凡等[49]的研究结果相一致。
不同的改造模式改变了不同植物群落的生态位宽度,从而改变了草本植物的多样性。从种类数量来看,柏木纯林中具有较高生态位宽度的优势草本物种有6种,而多样性指数低于柏木纯林的高低密度香椿、低密度银木与桤木+香椿+银木模式中优势草本物种分别为6种、4种、5种和5种,多样性大于柏木纯林的高密度银木、低密度桤木、香椿+银木和香椿+桤木模式中优势草本物种分别为10种、8种、7种和7种,这些模式中的丰富度指数又以低密度香椿、低密度银木与桤木+香椿+银木模式显著低于柏木纯林,高密度银木、低密度桤木模式显著高于柏木纯林。可以看出,改造后各模式草本植物生态位的变化导致了植物多样性的变化,拥有较大生态位宽度的优势种越多,植物多样性指数尤其是丰富度指数越高。在对科尔沁沙地植物恢复研究中,也能发现随演替时间推移,多样性增加,优势种也随之增加的规律[19],而在退化次生林恢复的研究中,植物多样性越高的次生林类型也拥有更多的生态位宽度较大的优势种[20]。
3.3 改造对草本植物种间关系的影响
种间关系在生物多样性维持中有重要的作用[50],通过生态位重叠可以揭示出不同物种间的种间关系。当两个或更多物种对一个资源或更多资源有共同利用时就会产生生态位重叠,生态位重叠一定程度的反映了物种间资源利用的相似性和物种的竞争关系[13],造成这种物种间生态位分化的来源可能是生活史差异、资源利用方式差异或类型差异等[51]。总的来说草本植物群落间的生态位重叠都比较低(均低于0.2),说明草本群落保持了一定的稳定性,生态位分化较明显,物种间利用资源的相似性较低,竞争不激烈,草本植物间可以较好的相互共存。在本研究中,由于对乔木层进行了改造,导致具有较强适应性的物种如薹草、竹叶草、五节芒等草本植物占据了优势地位,但由于改造时间不长,物种还处于资源利用扩张阶段,生态位还没有达到高度重叠而形成竞争,随着群落的发展和演替,物种的组成有可能还会继续改变。许多研究认为生态位宽度大的物种往往有着较大的生态位重叠[52],本研究中生态位宽度较大的物种间既有较大的生态位重叠,如薹草与蛇莓、薹草与地果;也有较小的生态位重叠,如竹叶草和薹草,地果与蛇莓,生态位重叠与生态位宽度间没有明显的规律[53-54],原因可能是生态位宽度较大的物种与其他物种的生态学特性和功能不一定相同,对资源的利用途径也存在异同,这也是植物对环境适应的结果[55],不能单纯的用生态位宽度来判断生态位重叠。而生态位宽度较小的物种如鼠麴草和蛇莓间却形成的较大的生态位重叠可能是由于两者对环境资源的需求相近或具有较高的聚集度[11]。
4 结论
草本层对环境的变化较为敏感,在改造初期更能反映植物群落的变化情况[56]。柏木低效林不同改造模式中香椿+桤木模式和香椿+银木模式对草本植物多样性的提高效果最好,是较为理想的改造模式,其他模式次之,高低密度香椿模式、低密度桤木模式和桤木+香椿+银木模式的改造效果差于柏木纯林;混交林模式较纯林模式改造效果好;银木和香椿模式高密度的改造效果优于低密度,桤木模式则为低密度优于高密度。草本层对逆境具有更强的耐受性,最能适应持久和强烈的干扰,对生态系统起到基础保障作用[57],草本植物多样性的提升预示着植物群落生境的改善,如光因子、土壤因子、生物因子等,但具体情况还需做进一步研究。川中丘陵区柏木低效林乔木层的改造打破了铁线蕨处于优势地位的草本群落结构,促进了更适应改造环境的草本群落如薹草、地果、竹叶草等植物群落的建立,实现了草本植物对资源分配的调节。植物的演替是长期的过程,由于改造时间较短,各改造模式草本植物还处于演替初期阶段,所有模式的草本层生态位重叠值低,分化较明显,种间竞争不激烈,物种处于资源扩张阶段。草本植物具有较强的不稳定性和多变性,随群落的发展和演替,喜阳植物五节芒等会继续被喜阴植物如薹草、竹叶草等取代,物种组成还会继续变化。
[1] Grinnell J. The niche-relationships of the California Thrasher. The Auk, 1916, 34(4): 427-433.
[2] 牛克昌, 刘怿宁, 沈泽昊, 何芳良, 方精云. 群落构建的中性理论和生态位理论. 生物多样性, 2009, 17(6): 579-593.
[3] Westman W L. Measuring realized niche spaces: climatic response of chaparral and coastal sage scrub. Ecology, 1991, 72(5): 1678-1684.
[4] 朱耿平, 刘国卿, 卜文俊, 高玉葆. 生态位模型的基本原理及其在生物多样性保护中的应用. 生物多样性, 2013, 27(1): 90-98.
[5] Tuomisto H. Edaphic niche differentiation amongPolybotryaferns in western Amazonia: implications for coexistence and speciation. Ecography, 2006, 29(3): 273-284.
[6] Kluge J, Kessler M. Influence of niche characteristics and forest type on fern species richness, abundance and plant size along an elevational gradient in Costa Rica. Plant Ecology, 2011, 212(7): 1109-1121.
[7] Gaston K J, Spicer J I. The relationship between range size and niche breadth: a test using five species ofGammarus(Amphipoda). Global Ecology and Biogeography, 2001, 10(2): 179-188.
[8] Kraft N J, Comita L S, Chase J M, Sanders N J, Swenson N G, Crist T O, Stegen J C, Vellend M, Boyle B, Anderson M J, Cornell H V, Davies K F, Freestone A L, Inouye B D, Harrison S P, Myers J A. Disentangling the drivers of β diversity along latitudinal and elevational gradients. Science, 2011, 333(6050): 1755-1758.
[9] Chesson P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 2000, 31(4): 343-366.
[10] Silvertown J, Dodd M, Gowing D. Phylogeny and the niche structure of meadow plant communities. Journal of Ecology, 2001, 89(3): 428-435.
[11] 胡正华, 钱海源, 于明坚. 古田山国家级自然保护区甜槠林优势种群生态位. 生态学报, 2009, 29(7): 3670-3677.
[12] 汤景明, 艾训儒, 易咏梅, 李玲, 徐红梅, 宋亚丽. 鄂西南木林子常绿落叶阔叶混交林恢复过程中优势树种生态位动态. 生态学报, 2012, 32(20): 6334-6342.
[13] 康永祥, 康博文, 岳军伟, 梁宗锁, 雷瑞德. 陕北黄土高原辽东栎(Quercusliaotungensis)群落类型划分及其生态位特征. 生态学报, 2007, 27(10): 4096-4105.
[14] 李斌, 李素清, 张金屯. 云顶山亚高山草甸优势种群生态位研究. 草业学报, 2010, 19(1): 6-13.
[15] 王惠, 邵国凡, 代力民, 许东, 王飞. 采伐干扰下长白山阔叶红松林主要灌木种群生态位动态特征. 东北林业大学学报, 2007, 35(11): 27-28.
[16] Laaka-Lindberg S, Pohjamo M, Korpelainen H. Niche breadth and niche overlap in three epixylic hepatics in a boreal old-growth forest, southern Finland. Journal of Bryology, 2005, 27(2): 119-127.
[17] 王立龙, 王广林, 黄永杰, 李晶, 刘登义. 黄山濒危植物小花木兰生态位与年龄结构研究. 生态学报, 2006, 26(6): 1862-1871.
[18] 林大影, 邢韶华, 赵勃, 崔国发. 北京山区森林植被中珍稀濒危草本植物生态位. 生态学杂志, 2007, 26(6): 781-786.
[19] 张继义, 赵哈林, 张铜会, 赵学勇. 科尔沁沙地植物群落恢复演替系列种群生态位动态特征. 生态学报, 2003, 23(12): 2741-2746.
[20] 刘宪钊, 陆元昌, 周燕华. 退化次生林恢复过程中群落结构和生态位动态. 生态学杂志, 2010, 29(1): 22-28.
[21] 王秀云, 朱汤军, 陆超, 沈建军, 徐高福. 千岛湖马尾松林抚育改造后主要种群生态位研究. 浙江林业科技, 2014, 34(3): 1-7.
[22] 王惠, 李倩, 韩雪梅, 邵国凡, 代力民. 采伐干扰下长白山阔叶红松林优势树种生态位变化动态. 东北林业大学学报, 2011, 39(10): 18-20, 53-53.
[23] 李平, 李凤汀, 范川, 李贤伟, 张健, 黄明莉. 川中丘陵区柏木低效林改造模式植物多样性对土壤有机碳的影响. 生态学报, 2015, 35(8): 2667-2675.
[24] 范川, 周义贵, 李贤伟, 张健, 廖洪流, 李凤汀, 冯茂松. 柏木低效林改造不同模式土壤抗蚀性对比. 林业科学, 2014, 50(6): 107-114.
[25] Gallet S, Roze F. Resistance of Atlantic Heathlands to trampling in Brittany (France): influence of vegetation type, season and weather conditions. Biological Conservation, 2001, 97(2): 189-198.
[26] Cole D N, Monz C A. Impacts of camping on vegetation: response and recovery following acute and chronic disturbance. Environmental Management, 2003, 32(6): 693-705.
[27] 王伟伟, 杨海龙, 贺康宁, 胡兴波, 王辉, 董梅, 谢增武. 祁连山青海云杉人工林草本层生态位及生态响应研究. 草地学报, 2012, 20(4): 626-630.
[28] 马克平, 刘玉明. 生物群落多样性的测度方法: Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239.
[29] Petraitis P S, Latham E R, Niesenbaum R A. The maintenance of species diversity by disturbance. The Quarterly Review of Biology, 1989, 64(4): 393-418.
[30] 任青山. 天然次生林主要种群生态位结构的研究. 东北林业大学学报, 1998, 26(2): 5-10.
[31] 李契, 朱金兆, 朱清科. 生态位理论及其测度研究进展. 北京林业大学学报, 2003, 2(1): 100-107.
[32] 张金屯. 数量生态学. 北京: 科学出版社, 2004.
[33] 刘金福, 洪伟. 格氏栲群落生态学研究——格氏栲林主要种群生态位的研究. 生态学报, 1999, 19(3): 347-352.
[34] 杨利民, 周广胜, 王国宏. 草地群落物种多样性维持机制的研究Ⅱ物种实现生态位. 植物生态学报, 2001, 25(5): 634-638.
[35] Gao T, Hedblom M, Emilsson T, Nielsen A B. The role of forest stand structure as biodiversity indicator. Forest Ecology and Management, 2014, 330: 82-93.
[36] 贾亚运, 何宗明, 周丽丽, 马祥庆, 吴鹏飞, 邹显花, 刘青青, 刘雨晖. 造林密度对杉木幼林生长及空间利用的影响. 生态学杂志, 2016, 35(5): 1177-1181.
[37] 马洪婧, 李瑞霞, 袁发银, 史珑燕, 关庆伟. 不同演替阶段栎树混交林群落稳定性. 生态学杂志, 2013, 32(3): 558-562.
[38] 岳永杰, 余新晓, 李钢铁, 樊登星, 叶俊道. 北京松山自然保护区蒙古栎林的空间结构特征. 应用生态学报, 2009, 20(8): 1811-1816.
[39] 袁王俊, 卢训令, 张维瑞, 王智慧, 张灵敏, 丁圣彦. 不同植被类型植物物种多样性. 生态学报, 2015, 35(14): 4651-4657.
[40] Macarthur R H, Connell J H. Organisms in communities. Science, 1966, 154(3752): 999-999.
[41] 王世雄, 王孝安, 李国庆, 郭华, 朱志红. 陕西子午岭植物群落演替过程中物种多样性变化与环境解释. 生态学报, 2010, 30(6): 1638-1647.
[42] 黎燕琼, 龚固堂, 郑绍伟, 陈俊华, 慕长龙, 朱志芳, 吴雪仙, 牛牧. 低效柏木纯林不同改造措施对水土保持功能的影响. 生态学报, 2013, 33(3): 934-943.
[43] 刘飞鹏, 曾曙才, 莫罗坚, 丁雪腾, 朱剑云, 刘颂颂, 李年生. 尾叶桉人工林改造对土壤和凋落物持水效能的影响. 生态学杂志, 2013, 32(5): 1111-1117.
[44] Alofs K M, Fowler N L. Loss of native herbaceous species due to woody plant encroachment facilitates the establishment of an invasive grass. Ecology, 2013, 94(3): 751-760.
[45] Brown J H. On the relationship between abundance and distribution of species. The American Naturalist, 1984, 124(2): 255-279.
[46] Burgman M A. The habitat volumes of scarce and ubiquitous plants: a test of the model of environmental control. The American Naturalist, 1989, 133(2): 228-239.
[47] Thompson K, Gaston K J, Band S R. Range size, dispersal and niche breadth in the herbaceous flora of central England. Journal of Ecology, 1999, 87(1): 150-155.
[48] Thompson K, Ceriani R M. No relationship between range size and germination niche width in the UK herbaceous flora. Functional Ecology, 2003, 17(3): 335-339.
[49] 钱逸凡, 伊力塔, 胡军飞, 张超, 余树全, 沈露, 彭东琴. 普陀山主要植物种生态位特征. 生态学杂志, 2012, 31(3): 561-568.
[50] Falcão J C F, Dáttilo W, Izzo T J. Efficiency of different planted forests in recovering biodiversity and ecological interactions in Brazilian Amazon. Forest Ecology and Management, 2015, 339(3): 105-111.
[51] Mangan S A, Schnitzer S A, Herre E A, Mack K M L, Valencia M C, Sanchez E I, Bever J D. Negative plant-soil feedback predicts tree-species relative abundance in a tropical forest. Nature, 2010, 466(7307): 752-755.
[52] 胡相明, 程积民, 万惠娥, 赵艳云. 黄土丘陵区不同立地条件下植物种群生态位研究. 草业学报, 2006, 5(1): 29-35.
[53] 钟宇, 张健, 刘泉波, 杨万勤, 吴福忠, 冯茂松. 巨桉人工林草本层主要种群的生态位分析. 草业学报, 2010, 19(4): 16-21.
[54] 陈玉凯, 杨琦, 莫燕妮, 杨小波, 李东海, 洪小江. 海南岛霸王岭国家重点保护植物的生态位研究. 植物生态学报, 2014, 38(6): 576-584.
[55] Silva I A, Batalha M A. Plant functional types in Brazilian savannas: The niche partitioning between herbaceous and woody species. Perspectives in Plant Ecology, Evolution and Systematics, 2011, 13(3): 201-206.
[56] Littlemore J, Barker S. The ecological response of forest ground flora and soils to experimental trampling in British urban woodlands. Urban Ecosystems, 2001, 5(4): 257-276.
[57] Grime J P. Plant Strategies, Vegetation Processes, and Ecosystem Properties. 2nd ed. New York: Wiley, 2003.
