广东省森林死木碳库特征
2018-03-08赵嘉诚李海奎
赵嘉诚,李海奎
中国林业科学研究院资源信息研究所,北京 100091
死木是森林生态系统的重要组成部分,不仅对推动生态系统能量流动[1- 2],固碳[3- 5],森林演替更新[6- 7],有着重要的影响,而且在维持物种多样性等方面也发挥着不可或缺的作用,例如,为鸟类觅食,筑巢,贮藏食物提供必要场所[8- 9],为哺乳动物提供栖息地等[10- 11]。按照IPCC的定义,死木包括枯立木、枯倒木、死根和直径大于或等于10 cm(或不同国家规定的大小)的树桩[12],是森林生态系统的五大碳库之一,也是IPCC要求报告的数据。在全球气候变化的背景下,研究死木碳库的动态和特征,对于揭示森林生态系统碳循环规律,明确森林生态系统碳汇(源)过程,有着重要的意义。
在以往对森林生态系统死木的研究中,对象主要集中在粗木质残体(Coarse Woody Debris,CWD),以及它和细木质残体、树桩组成的枯倒木(Down and Dead Woody,DDW)碳库[13];研究内容主要是碳库贮量、死木的成因和腐烂分解速率等[14- 23];研究尺度多为区域性或特定森林类型[24- 27]。这些研究主要从生态学的角度出发,粗木质残体碳库数据多为存量。国际履约的需要,引起了各国林业工作者对死木碳库研究的重视,但由于调查成本过高,只有美国、日本和俄罗斯等少数国家开展了国家尺度上的枯倒木调查[13, 28- 32]。据估计,在美国生态系统中,大约14%的碳存储于死有机质(死木和枯枝落叶层,不包括细木质残体)中[33]。枯倒木在森林中普遍存在,地区间的差异主要由地方生态系统和经营实践引起,单位面积枯立木、活立木和枯倒木的生物量之间复杂的关系,主要取决于林分相对密度和平均年降水量或最低温度等因子的变化[28]。而国内在国家尺度上尚没有针对死木碳库的专项调查,在大尺度上的相关研究和估算几乎没有报道。
本文依据第8次(2009—2013)全国森林资源连续清查中广东省的调查数据(2012),采用分树种、分组分的生物量模型,估算2007—2012年间,广东省森林中的死木碳库变化,研究其树种分布、龄组比例和死亡成因等特征;在2016年系统抽取八分之一的固定样地中,通过典型抽样方法,抽取2012年调查时已发生死亡样木较多的样地进行实测调查,记录死木至调查时的腐朽程度(这些腐朽分解发生在2012—2016年间),由此推算广东省全省死木碳库在近10年间的动态,为精准估算广东省森林死木碳库储量提供科学依据。
1 研究区概况
研究区广东省位于我国南部,属亚热带-热带湿润季风气候,地理坐标为20°15′—25°31′N、109°40′—117°20′E,自然资源丰富,水热条件优厚,高温多雨。全年平均气温在19°以上,平均降水量多在1500 mm以上,一般雨季为4—9月,降水量约占全年总量的70%—90%。全省地势大体上表现为北高南低,山地丘陵约占总面积的2/3,粤北山地海拔为1000—1500 m。全省森林总面积为1082.79万hm2,森林覆盖率58.69%,总蓄积5.47亿m3。北部南岭地区的典型植被为亚热带山地常绿阔叶林,中部为亚热带常绿季雨林,南部为热带常绿季雨林,主要以针叶林、中幼林为主[34]。常见的针叶树包括马尾松(Pinusmassoniana)、杉木(Cunninghamialanceolata)、湿地松(Pinuselliottii)、火炬松(Pinustaeda)等。
2 数据来源
本文的数据来源分为两个部分,一是第8次全国森林资源连续清查中广东省固定样地的样木数据和样地数据,调查在2012年进行。按检尺类型,死木可以分为枯立木、枯倒木和采伐木,死亡发生在2007—2012年之间,样木表中记录的死木胸径为前期调查(2007)时,尚未死亡时的胸径,样地数据记录了死木所在样地的林分特征,包括林分类型和龄组等;二是2016年进行的已有死木腐朽等级调查数据。调查在系统抽取八分之一固定样地的基础上,通过典型抽样,在死木较多的样地内进行。表1和表2给出了数据的基本统计量,从中可以看到系统抽取的八分之一固定样地中枯立木、枯倒木和死木总数,基本上为全部样地的八分之一,涉及到的样地数也接近八分之一,覆盖了大多数的树种;典型抽取的实际调查样地,虽然样地数只占八分之一固定样地数的15%,但死木株数却占40%左右,也覆盖了大多数树种。

表1 全部样地死木统计表

表2 八分之一样地死木和腐朽等级调查样地死木统计表
3 研究方法
3.1 腐朽分级
8次清查的死木数据,在2012年调查时,并没有记录其腐朽程度,只是区分了枯立木和枯倒木,知道其枯死发生在之前的0—5年之间。在没有发生大的自然灾害且林木自然生长的情况下,研究假设每年发生枯死的树木数量是相等的,这时每株死木的平均死亡时间为2—3年,按照闫恩荣等[35]提出的5级森林生态系统CWD分类系统(表3),可以基本认为枯立木或枯倒木的腐朽等级为第1级。2016年实际调查的死木是严格按5级CWD分类系统进行腐朽等级划分的。以死木胸径为协变量,用多因素方差分析的方法确定死木种类、树种组对腐朽等级的影响,为精准估算死木保留碳储量提供方法选择。
3.2 碳计量参数
对于表1中17个树种碳计量参数的选择,其中马尾松、杉木、湿地松、栎类、木荷和枫香这六个树种,采用国家林业局发布的行业标准[36- 41],这些行业标准中的生物量方程包括与总量兼容的各个组分的生物量参数和区分地上、地下生物量的含碳系数;对于柏木、其他硬阔和桉树,使用文献[42]中的区分各个组分的生物量方程和含碳系数;以上9个树种的死木株数占全部死木的81.3%;对于铁杉、水杉、樟木、楠木和楝树等8个树种,由于文献中[42]只区分了地上生物量和地下生物量,故采用与此关系密切的其他树种的各个生物量组分比例关系确定这些树种各个组分的生物量,树种关系的密切程度及其归并见文献[43]。
3.3 单木碳储量计算
单木碳储量是树木各个组成部分碳储量之和,而碳储量通常是由生物量乘以树种含碳系数而获得的。所以对于活立木:
Bab=Bst+Bba+Bbr+Ble
(1)
Bbl=Broot
(2)
Btotal=Bab+Bbl
(3)
Ctotal=Btotal×Crate
(4)
式(1)—(4)中,Bab:地上总生物量;Bst:干材生物量;Bba:树皮生物量;Bbr:树枝生物量;Ble:树叶生物量;Bbl:地下生物量;Broot:树根生物量;Btotal:总生物量;Ctotal:总碳储量;Crate:含碳系数。
含碳系数细分的情况下,式(3)—(4)可以变换为:
Ctotal=Bab×Crab+Bbl×Crbl
(5)
式中,Crab:地上生物量含碳系数;Crbl:地下生物量含碳系数。
对于单株枯死木,首先根据死木类型和腐朽等级,给予各个组分用于计算现存生物量的权重(表3),然后按照式(6)计算其碳储量:
(6)

表3给出了枯立木和枯倒木腐朽等级及其现存生物量权重。

表3 枯立木和枯倒木腐朽等级及其现存生物量权重
3.4 全省死木碳储量估算
基于8次清查数据的死木碳储量(2007—2012年间新增死木碳库,以下简称2012年碳库)按照森林资源连续清查估算蓄积量的方法估算。
基于腐朽等级调查数据的全省死木保留碳储量(2007—2012年间新增死木到2016年时的保留碳库,以下简称2016年碳库)的估算,按以下公式计算:
(7)
(8)
式中,TDC:全省死木总保留碳储量;DCi:第i株死木碳储量(2012年);BDCi:第i株死木保留碳储量(2016年);n:八分之一样地的死木总株数;N:全部样地的死木总株数;m:腐朽等级调查的样木株数;S:由样地水平到全省尺度的扩展系数。
式(7)和式(8)的差别在于八分之一样地死木对总体样地死木的代表性。在不同树种间死木腐朽等级存在显著差异的情况下,式(7)和式(8)变为:
(9)
(10)
式中,DCji:第j树种的第i株死木碳储量(2012年);BDCji:第j树种第i株死木保留碳储量(2016年);nj:八分之一样地中第j树种死木株数;Nj:全部样地中第j树种死木株数;mj:第j树种腐朽等级调查的样木株数;k:树种数。
由于在全省固定样地与抽样的八分之一样地中,死木均以杉木、马尾松、湿地松、栎类、其他硬阔、其他软阔等六个树种组的株数为最多,所以在死木腐朽等级调查时,把数量较少的树种合并到相近的树种中,例如把桉树、泡桐等软阔树种和其他软阔树种合并,形成软阔树种组。这样,所最终确定的树种组为杉木、马尾松、湿地松、栎类、其他硬阔和软阔。
3.5 死木成因分析及碳库特征
按照表4对死木形成原因进行初步分类,结合气象资料,例如台风、冰雪、火灾和病虫等自然灾害统计等资料,对死木成因进行综合分析,并对2012年的死木碳库,按枯立木、枯倒木,分死木所在林分的林分类型(不是死木具体的树种)和龄组等,分析死木碳库数量特征。

表4 样地死木特征及形成原因判别
4 结果与分析
4.1 死木腐朽程度
由于清林等人为活动的影响,2012年(期初)调查时的死木,在2016年(期末)时,只有大约43%的保留了下来,分树种保留木的平均胸径和腐朽程度见表5。其中不同树种的腐朽程度相差较大,腐朽等级从小到大的排序顺序为:杉木、其他硬阔、马尾松、栎类、湿地松和软阔。进一步的方差分析(显著性水平为0.05)表明:死木的胸径大小与腐朽程度不存在显著性差异;死木类型间存在显著性差异,枯倒木的腐朽程度(3.7)大于枯立木(2.9)相差近一个等级;树种组之间存在显著性差异,其中杉木腐朽程度最低,与其他所有树种组之间均存在显著性差异,软阔与湿地松腐朽程度最高,它们之间差异不显著,而与其他树种组差异显著,其余树种组之间差异不显著。
4.2 死木库碳储量
表6给出了2012年碳库和4种计算方法估算的2016年碳库。结果显示,2012年广东全省乔木林死木碳库为5811.86 Pg,占同期乔木林活立木生物量碳库的2.94%;其中,枯立木碳库为2230.44 Pg,占死木碳库的38.38%,枯倒木碳库为3581.42 Pg,占61.62%;地上部分碳库为4687.09 Pg,占死木木碳库的80.65%,地下部分碳库为1124.77 Pg,占19.35%。死木库碳密度(单位面积碳储量)为(0.7612 ± 3.3988) Mg/hm2,其中,枯立木碳密度为(0.2806 ± 1.7288) Mg/hm2,枯倒木碳密度为(0.4806 ± 2.2835) Mg/hm2;地上部分碳密度为(0.5898 ± 2.7683) Mg/hm2,地下部分碳密度为(0.1415 ± 0.6365) Mg/hm2。所有的碳密度都有较大的标准差,这主要是因为,尽管死木只出现在不到1/3的样地中(表1),但在出现死木的样地中,有可能全部样木均为死木。通过四种方法计算的2016年碳库,全省总碳储量从最小的785.57 Pg(式(9))到最大的1089.52 Pg(式(8)),两者相差近40%(以最小值为基数),但相对应于2012年的碳库,最大和最小减少率分别为86.48%和81.25%,两者差别不大,地上部分和地下部分碳储量也有同样的趋势。死木保有碳储量锐减的主要原因是大多数死木已经被清林等人为经营活动被移出,同时广东省高温、多雨的特点加速了死木的腐烂。
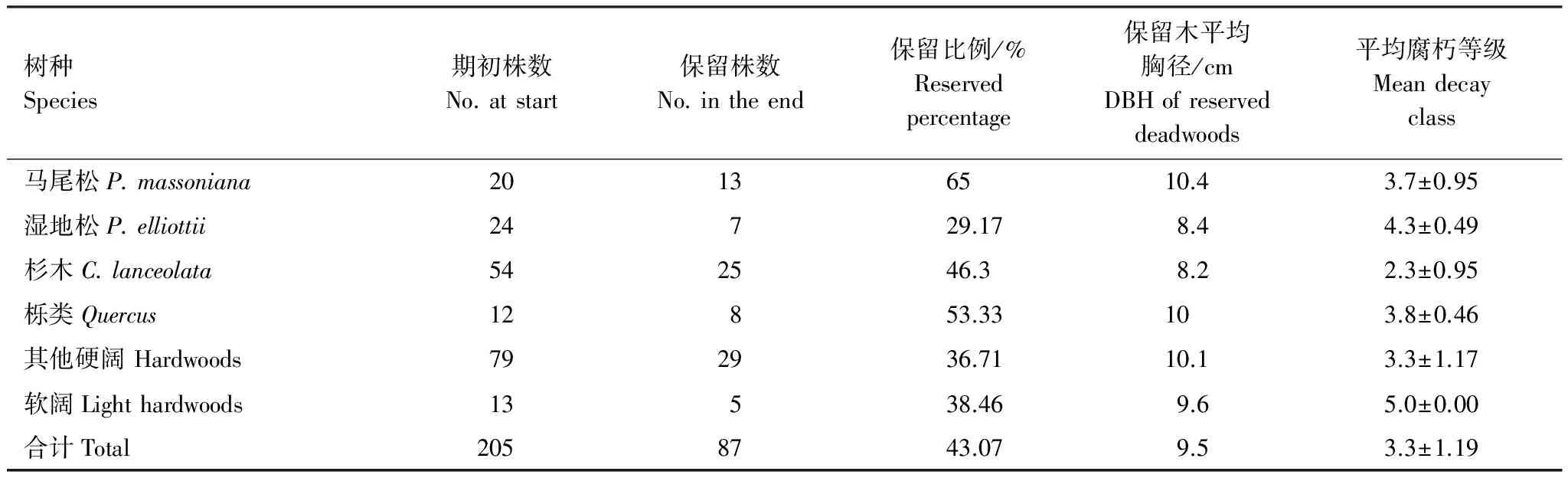
表5 分树种的死木腐朽程度统计表

表6 广东死木库碳储量和碳密度
对于四种估算方法的选用,由于树种间的腐朽程度存在显著性差异,式(9)和式(10)的估算结果理论上应优于式(7)和式(8),同时八分之一样地的抽样没有直接的全部样地的计算结果准确,所以我们认为式(9)给出了最准确的估算结果785.57 Pg。
4.3 死木成因及碳库特征

图1 死木的成因比例图Fig.1 Proportions of deadwoods with different cause of death
图1给出了造成死木死亡的不同原因的比例关系,从中可以发现:自然竞争存在于50%以上死木的样地,但其造成的死木碳储量却不足15%;衰老只存在于10%左右的死木样地中,其碳储量也约为10%;风倒、冰雪、火灾或病虫等自然灾害虽然只发生在不到10%的死木样地中,但造成的死木碳储量却超过40%,特别是风倒或冰雪灾害,只发生在5%左右的死木样地中,却有超过30%的死木碳储量,这与2008年我国南方发生了罕见的大面积冰雪灾害,致使粤北地区很多森林的死木数量剧增有密切关系,另有30%的死木样地和死木碳储量不易判断其成因。
表7列出了2012年死木碳库按林分类型和龄组的数量分配特征。阔叶混交林的死木碳储量占死木总储量的近50%,马尾松也超过20%,其余8个林分类型的比例均不超过10%,其中桉树和杉木的比例均不到2%,这也是经营强度最大的两种森林类型;与同类林分的现存碳储量相比,马尾松占比超过达7.45%,其余林分类型均不超过5%,其中桉树和杉木不到1%,从大到小的排列顺序为:马尾松、其他软阔、湿地松、其他硬阔、阔叶混交林、栎类、针阔混交林、针叶混交林、杉木和桉树,全省平均占比为2.97%。按龄组分,中龄林的死木碳储量占总碳储量比例最大,达44.77%,过熟林比例最小,仅为5.12%,比例从大到小的排列顺序为:中龄林、近熟林、幼龄林、成熟林和过熟林;与同龄组林分的现存碳储量相比,各龄组相差不大,从2.03%到4.56%,其中幼龄林最小,成熟林最大。
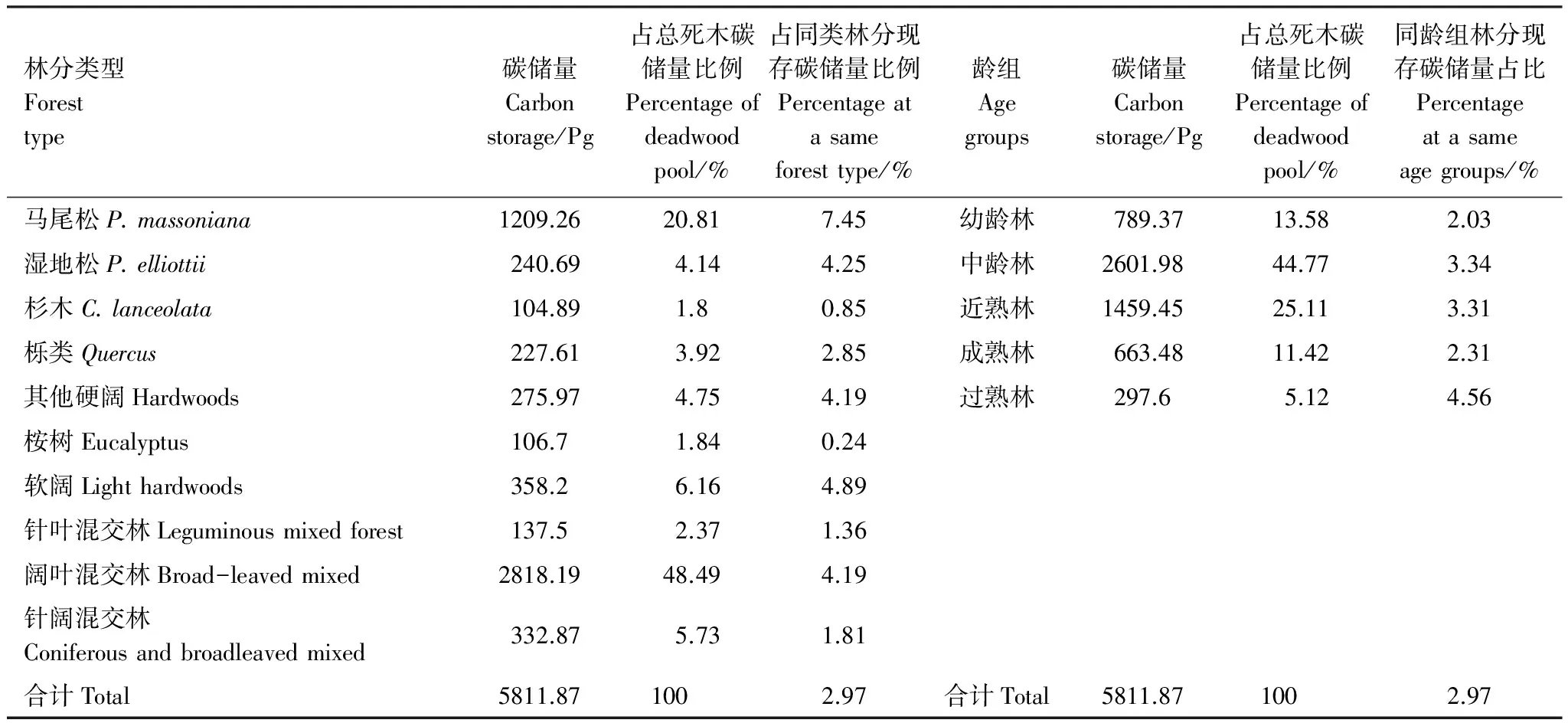
表7 2012年死木碳库按林分类型和龄组的数量分配特征
5 讨论
包括枯立木、枯倒木、根桩和大枯枝等在内的粗木质残体(CWD),是森林生态系统的重要组成部分、营养库和碳库[14]。本文估算的广东省2012年死木碳储量是2007—2012年间增加到死木碳库中的储量,这并不包括2007年以前的那些到2012年时仍然保留在死木库的储量,按照本文调查的腐朽程度推算,死木碳库中最近10年的新增的碳储量不大于6599.43 Pg, 占全省活立木碳储量的比例仅大于3%,远低于美国生态系统中粗木质残体的碳储量占比(约14%)[33];碳密度不超过0.86 Mg/hm2,低于李凌浩等[44]对武夷山甜槠林CWD碳储量(约3.67 Mg/hm2)的估计,远低于杨方方等[45]对鼎湖山季风常绿阔叶林CWD碳储量(约21 Mg/hm2)的估计,但介于张修玉等[46]对广州森林CWD碳储量估计的范围内(约0.11—9.1 Mg/hm2)。气候差异、森林类型、树种组成、林分龄组、现有活立木碳储量和研究区域的大小等,可能是造成死木碳存量不一致的内源性原因。广东省处于亚热带和热带地区,高温多雨,水、气、热条件较好,树木死亡后枯倒木数量多于枯立木(表1,表2),同时,死后的腐朽程度,枯倒木也重于枯立木(表5)等现象,均与寒冷恶劣的气候条件会减缓分解过程的研究结果一致[44,47]。林龄对死木碳储量的影响主要表现为,从幼龄林到近熟林,死木存量逐渐积累,到林分成熟时,由竞争和人为扰动带来的林木死亡进一步增加了死木碳库存量,而当林分衰老时,CWD开始下降[48];邓绍林等[49]在对桂西北杉木立木腐朽程度研究中也指出,林龄对杉木腐朽的影响多表现为腐朽率随林龄的增大而增大,而本文研究范围内,除杉木外,还包含了较多的树种和森林类型,但按龄组划分,死木碳储量占同类森林的比例总体上也基本呈现出从幼龄林(2.03%)到过熟林(4.56%)的上升趋势(表7)。森林经营、人类活动以及不同抽样方法所带来的野外测量和计算过程的差异等外源性原因亦会对CWD估算结果产生巨大影响[50-51]。例如,当前国际上对死木的调查方法主要有样方法和样带法[29,52],但范围较小,而本文研究的省级尺度的死木碳库,采用的是固定样地的资料和典型抽样的方法,并没有包括采伐木的伐桩和树根等碳储量。Krankina等[53]在比较瑞典和俄罗斯森林CWD储量时指出,森林经营的强度要比不同的野外测量方法对CWD估算影响更大,一些瑞典北部的经营强度相对较高的森林具有更低的CWD与活立木的材积比。同样,本文中的桉树和杉木这些集约经营强度大的人工林中,死木比例也较低,均不足1%(表7)。
林木腐朽这一复杂的生态学过程,除了受温度、湿度、氧气、二氧化碳、代谢基质和微生物等多种因素的共同影响[54]之外,与树种各自的生物学特性也有关。张修玉等[46]通过对广州3种森林CWD储量与分解特征的调查分析得到,常绿阔叶林的CWD分解速率大于混交林,而混交林又大于针叶林,这一结论在本研究中也得到了印证,例如:本文不同树种之间死木的腐朽程度表现出显著性差异,杉木的腐朽分解最慢,而软阔腐朽程度最高,所有被调查到的软阔类死木的腐朽等级均为5级(表5)。但从总体来看,腐朽等级以2级和3级为最多,与蔡慧颖等[55]以及Oheimb等[56]得到的CWD多分布在中等腐朽等级的研究结果类似。
自然状态下,死木的形成主要来自于以下几个原因:(1)病虫,风倒,冰雪,森林火等自然灾害[20-21];(2)群落的生长竞争所引发的个体密度逐渐降低的自然稀疏(self thinning)过程[22];(3)树木在达到老龄时自然衰老死亡[17]。本文研究发现:林分本身的竞争和衰老,虽然发生的60%的死木样地中,但增加到死木碳库中储量不足总量的四分之一,而自然灾害虽然只发生在10%的死木样地中,对死木碳库的贡献却超过40%,特别是风倒或冰雪灾害,只发生在5%左右的死木样地中,却有超过30%的死木碳储量,这与2008年我国南方发生了罕见的大面积冰雪灾害有密切关系。死木碳库在林分类型间的分布不均,其中阔叶混交林和马尾松占近7成,这两个森林类型死木碳库占活立木碳库的比例也是最高的,特别是马尾松林;死木碳库在林分龄组间的分布,中龄林和近熟林比例最大,占同龄组林分活立木碳库的比例也较高,过熟林虽然在死木库中所占比例不大,但单位面积储量大,总活立木碳储量小,当严重灾害发生时,造成其死木占活立木的比例最大。
6 结论
2007—2012年间广东省乔木林死木碳库新增碳储量为5811.86 Pg,占同期乔木林活立木生物量碳库的2.94%,新增死木库碳密度为(0.7612 ± 3.3988) Mg/hm2。枯倒木新增碳储量多于枯立木;阔叶混交林和马尾松贡献了近70%的死木碳储量;马尾松、其他软阔等林分类型的死木碳储量占同类森林总活立木碳储量的比例较大;按龄组划分,中龄林占总死木碳储量比例最大,过熟林最小。占同龄组林分现存碳储量的比例,从幼龄林到过熟林基本呈上升趋势。竞争引起的枯死在林分中普遍存在,但由此导致的新增死木碳库的储量不大,而自然灾害对死木碳库储量有较大的影响。到2016年,原有2007—2012年间新增碳储量下降到785.57 Pg,减少约85%,清林等人为经营活动和死木较快地腐朽是存量减少的主要原因。
[1] Graham R L, K Cromack Jr. Mass, nutrient content, and decay rate of dead boles in rain forests of Olympic national park. Canadian Journal of Forest Research, 1982, 12(3): 511- 521.
[2] Finér L, Mannerkoski H, Piirainen S, Starr M. Carbon and nitrogen pools in an old-growth, Norway spruce mixed forest in eastern Finland and changes associated with clear-cutting. Forest Ecology and Management, 2003, 174(1/3): 51- 63.
[3] Janisch J E, Harmon M E. Successional changes in live and dead wood carbon stores: implications for net ecosystem productivity. Tree Physiology, 2002, 22(2/3): 77- 89.
[4] Gough C M, Vogel C S, Kazanski C, Nagel L, Flower C E, Curtis P S. Coarse woody debris and the carbon balance of a north temperate forest. Forest Ecology and Management, 2007, 244(1/3): 60- 67.
[5] Woodall C W, Liknes G C. Climatic regions as an indicator of forest coarse and fine woody debris carbon stocks in the United States. Carbon Balance and Management, 2008, 3(1): 5.
[6] Hagan J M, Grove S L. Coarse woody debris: humans and nature competing for trees. Journal of Forestry, 1999, 97(1): 6- 11.
[7] Weaver J K, Kenefic L S, Seymour R S, Brissette J C. Decaying wood and tree regeneration in the Acadian Forest of Maine, USA. Forest Ecology and Management, 2009, 257(7): 1623- 1628.
[8] Conner R N. Foraging habitats of woodpeckers in southwestern Virginia. Journal of Field Ornithology, 1980, 51(2): 119- 127.
[9] Gibbs J P, Hunter Jr M L, Melvin S M. Snag availability and communities of cavity nesting birds in tropical versus temperate forests. Biotropica, 1993, 25(2): 236- 241.
[10] Freedman B, Zelazny V, Beaudette D, Fleming T, Johnson G, Flemming S, Forbes G, Gerrow J S, Johnson G, Woodley S. Biodiversity implications of changes in the quantity of dead organic matter in managed forests. Environmental Reviews, 1996, 4(3): 238- 265.
[11] Ohlson M, Söderström L, Hörnberg G, Zackrisson O, Hermansson J. Habitat qualities versus long-term continuity as determinants of biodiversity in boreal old-growth swamp forests. Biological Conservation, 1997, 81(3): 221- 231.
[12] IPCC. Guidelines for national greenhouse gas inventories (volume 4): agriculture, forestry and other land use. [2017-01- 17]. http://www.ipcc-nggip.iges.or.jp/public/2006gl/pdf/4_Volume4/V4_01_Ch1_Introduction.pdf.
[13] Woodall C W, Heath L S, Smith J E. National inventories of down and dead woody material forest carbon stocks in the United States: challenges and opportunities. Forest Ecology and Management, 2008, 256(3): 221- 228.
[14] 陈华, 徐振邦. 粗木质物残体生态研究历史、现状和趋势. 生态学杂志, 1991, 10(1): 45- 50.
[15] 杨玉盛, 郭剑芬, 林鹏, 何宗明, 陈光水. 格氏栲天然林与人工林粗木质残体碳库及养分库. 林业科学, 2005, 41(3): 7- 11.
[16] Guo L B, Bek E, Gifford R M. Woody debris in a 16-year oldPinusradiataplantation in Australia: mass, carbon and nitrogen stocks, and turnover. Forest Ecology and Management, 2006, 228(1/3): 145- 151.
[17] Garrett L, Davis M, Oliver G. Decomposition of coarse woody debris, and methods for determining decay rates. New Zealand Journal of Forestry Science, 2007, 37(2): 227- 240.
[18] 刘妍妍, 金光泽. 地形对小兴安岭阔叶红松(Pinuskoraiensis)林粗木质残体分布的影响. 生态学报, 2009, 29(3): 1398- 1407.
[19] Rouvinen S, Kuuluvainen T, Karjalainen L. Coarse woody debris in oldPinussylvestrisdominated forests along a geographic and human impact gradient in boreal Fennoscandia. Canadian Journal of Forest Research, 2002, 32(12): 2184- 2200.
[20] Everham E M, Brokaw N V L. Forest damage and recovery from catastrophic wind. The Botanical Review, 1996, 62(2): 113- 185.
[21] Woodall C W, Nagel L M. Downed woody fuel loading dynamics of a large-scale blowdown in northern Minnesota, U.S.A. Forest Ecology and Management, 2007, 247(1/3): 194- 199.
[22] Hansen A J, Spies T A, Swanson F J, Ohmann J L. Conserving biodiversity in managed forests. BioScience, 1991, 41(6): 382- 392.
[23] Lee P C, Crites S, Nietfeld M, Nguyen H V, Stelfox J B. Characteristics and origins of deadwood material in aspen-dominated boreal forests. Ecological Applications, 1997, 7(2): 691- 701.
[24] 王文娟, 常禹, 刘志华, 陈宏伟, 荆国志, 张红新, 张长蒙. 大兴安岭呼中林区火烧迹地粗木质残体特征. 应用生态学报, 2009, 20(8): 1805- 1810.
[25] 袁杰, 蔡靖, 侯琳, 张硕新. 秦岭火地塘天然次生油松林倒木储量与分解. 林业科学, 2012, 48(6): 141- 146.
[26] 刘妍妍, 金光泽. 小兴安岭阔叶红松林粗木质残体基础特征. 林业科学, 2010, 46(4): 8- 14.
[27] 罗大庆, 郭泉水, 黄界, 潘刚, 辛学兵, 郑维列. 西藏色季拉原始冷杉林死亡木特征研究. 生态学报, 2004, 24(3): 635- 639.
[28] Woodall C W, Walters B F, Oswalt S N, Domke G M, Toney C, Gray A N. Biomass and carbon attributes of downed woody materials in forests of the United States. Forest Ecology and Management, 2013, 305(4): 48- 59.
[29] Ugawa S, Takahashi M, Morisada K, Takeuchi M, Matsuura Y, Yoshinaga S, Araki M, Tanaka N, Ikeda S, Miura S, Ishizuka S, Kobayashi M, Inagaki M, Imaya A, Nanko K, Hashimoto S, Aizawa S, Hirai K, Okamoto T, Mizoguchi T, Torii A, Sakai H, Ohnuki Y, Kaneko S. Carbon stocks of dead wood, litter, and soil in the forest sector of Japan: general description of the national forest soil carbon inventory. Bulletin of FFPRI, 2012, 11(4): 207- 221.
[30] Kukuev Y A, Krankina O N, Harmon M E. The forest inventory system in Russia: a wealth of data for western researchers. Journal of Forestry, 1997, 95(9): 15- 20.
[31] Fridman J, Walheim M. Amount, structure, and dynamics of dead wood on managed forestland in Sweden. Forest Ecology and Management, 2000, 131(1/3): 23- 36.
[32] Woldendorp G, Keenan R J. Coarse woody debris in Australian forest ecosystems: a review. Austral Ecology, 2005, 30(8): 834- 843.
[33] Heath L S, Smith J E, Birdsey R A. Carbon trends in U.S. forestlands: a context for the role of soils in forest carbon sequestration//Kimble J, Heath L S, Birdsey R A, Lal R, eds. The Potential of U.S. Forest Soil to Sequester Carbon and Mitigate the Greenhouse Effect. Boca Raton: CRC Press, 2003: 35- 45.
[34] 广东省林业厅. 林业概况. (2015-04- 30) [2017-01- 17].https://www.gdf.gov.cn/index.php?controller=front&action=view&id=10084.
[35] 闫恩荣, 王希华, 黄建军. 森林粗死木质残体的概念及其分类. 生态学报, 2005, 25(1): 158- 167.
[36] 国家林业局. LY/T 2263—2014 立木生物量模型及碳计量参数——马尾松. 北京: 中国林业出版社, 2014.
[37] 国家林业局. LY/T 2264—2014 立木生物量模型及碳计量参数——杉木. 北京: 中国林业出版社, 2014.
[38] 国家林业局. LY/T 2261—2014 立木生物量模型及碳计量参数——湿地松. 北京: 中国林业出版社, 2014.
[39] 国家林业局. LY/T 2658—2016 立木生物量模型及碳计量参数——栎树. 北京: 中国林业出版社, 2016.
[40] 国家林业局. LY/T 2660—2016 立木生物量模型及碳计量参数——木荷. 北京: 中国林业出版社, 2016.
[41] 国家林业局. LY/T 2661—2016 立木生物量模型及碳计量参数——枫香. 北京: 中国林业出版社, 2016.
[42] 李海奎, 雷渊才. 中国森林植被生物量和碳储量评估. 北京: 中国林业出版社, 2010.
[43] 国家林业局. 国家森林资源连续清查技术规定. 北京: 国家林业局森林资源管理司, 2014.
[44] 李凌浩, 邢雪荣, 黄大明, 刘初钿, 何建源. 武夷山甜槠林粗死木质残体的贮量、动态及其功能评述. 植物生态学报, 1996, 20(2): 132- 143.
[45] 杨方方, 李跃林. 鼎湖山粗死木质残体生物量特征. 应用与环境生物学报, 2011, 17(5): 750- 752.
[46] 张修玉, 管东生, 张海东. 广州三种森林粗死木质残体(CWD)的储量与分解特征. 生态学报, 2009, 29(10): 5227- 5236.
[47] 肖洒, 吴福忠, 杨万勤, 常晨晖, 李俊, 王滨, 曹艺. 高山峡谷区暗针叶林木质残体储量及其分布特征. 生态学报, 2016, 36(5): 1352- 1359.
[48] Sturtevant B R, Bissonette J A, Long J N, Roberts D W. Coarse woody debris as a function of age, stand structure, and disturbance in boreal Newfoundland. Ecological Applications, 1997, 7(2): 702- 712.
[49] 邓绍林, 韦西华, 韦继兴, 黄焕扬, 田永江, 余长青. 桂西北杉木立木腐朽的调查研究. 广西林业科学, 1998, 27(2): 80- 83.
[50] 谷会岩, 代力民, 王顺忠, 于大炮, 周莉. 人为干扰对长白山红松针阔叶混交林粗木质残体的影响. 林业科学, 2006, 42(10): 1- 5.
[51] 谷会岩, 金靖博, 陈祥伟, 王恩姮, 周一杨, 柴亚凡. 人为干扰对大兴安岭北坡兴安落叶松林粗木质残体的影响. 应用生态学报, 2009, 20(2): 265- 270.
[52] Williams M S, Ducey M J, Gove J H. Assessing surface area of coarse woody debris with line intersect and perpendicular distance sampling. Canadian Journal of Forest Research, 2005, 35(4): 949- 960.
[53] Krankina O N, Harmon M E, Kukuev Y A, Treyfeld R F, Kashpor N N, Kresnov V G, Skudin V M, Protasov N A, Yatskov M, Spycher G, Povarov E D. Coarse woody debris in forest regions of Russia. Canadian Journal of Forest Research, 2002, 32(5): 768- 778.
[54] Harmon M E, Franklin J F, Swanson F J, Sollins P, Gregory S V, Lattin J D, Anderson N H, Cline S P, Aumen N G, Sedell J R, Lienkaemper G W, Cromack K Jr, Cummins K W. Ecology of coarse woody debris in temperate ecosystems. Advances in Ecological Research, 1986, 15: 133- 302.
[55] 蔡慧颖, 邸雪颖, 金光泽. 小兴安岭谷地云冷杉林粗木质残体碳密度特征. 生态学报, 2015, 35(24): 8194- 8201.
[56] von Oheimb G, Westphal C, Härdtle W. Diversity and spatio-temporal dynamics of dead wood in a temperate near-natural beech forest (Fagussylvatica). European Journal of Forest Research, 2007, 126(3): 359- 370.
