红松人工林生物量碳密度
2018-03-07丁壮崔若光
丁壮 崔若光
(东北林业大学,哈尔滨,150040)
红松人工林生态系统生物量碳密度,其生物量是NPP减去当年的凋落物,即形成年生产量,用此数据计算生物量碳密度。生物量碳密度是生物量碳库的基本指标,生物量的贮碳器官由叶、枝、树干、果、根构成,具有吸碳、贮碳和转移碳的功能,与土壤碳密度、凋落物碳密度相比,属基础碳密度[1]。只有把生物量碳密度经营好,才能不断增加生物量碳密度,从而提高红松人工林碳汇水平。
现在很多学者对生物量碳密度及其碳汇研究十分重视,而且不断深入开展起来,但都偏重不同气候带,不同区域的研究,而对东北红松人工林及其天然阔叶树种混交林分的研究并不普遍。本文以东北林业大学老山人工林实验站营造红松人工纯林和白桦红松混交林,蒙古栎红松混交林生物量碳密度的研究作为重点,探索生物量碳密度的特点和规律,为提高红松人工林及其混交林生物量碳汇创造条件。
1 研究区概况
试验地点位于黑龙江省尚志市帽儿山镇东北林业大学老山人工林实验站,地理坐标为45°14′~45°20′N,127°29′~127°44′E。属长白山系北部张广才岭西坡,地貌为低山丘陵区,平均海拔300 m,平均坡度为12°,平均气温为2.8 ℃,平均湿度70%,年蒸发量为1 093 mm,地带性土壤为暗棕壤,非地带性土壤为白浆土等土种,约占土壤面积三分之一,原始植被为阔叶红松林,在历史上曾遭受严重破坏,现已基本恢复人工林和天然次生林的植被[2]。
2 研究方法
样地设置:在20年生和42年生红松人工纯林中各选3块样地,在同样林龄中,白桦红松混交林各选3块,蒙古栎红松混交林中各选3块,共计18块样地,其中6块固定样地,面积均为40 m×25 m。
乔木层生物量碳密度:在样地每木调查的基础上,按着径级大小排序分为5个数量相等的组合,并计算各组合的平均胸径、树高及活枝下高,按着这个标准的理论值,在样地中选择径级不等的样木,并使理论值与实际值的精度不超过5%,共选择5株样木,按此方法,两种林龄型3种类型样地,共选择30株样木。确定样木后逐一伐倒,量测全高,胸径及枝下高,并将树冠分为上层、中层和下层,在每层中测定叶、枝和果的生物量,树干是按1 m区分段测定生物量。
将每株样木生物量分别乘以所代表的组合株数,即得每组合生物量,再把样地5株样木组合生物量合计,即得样地生物量,减去含水量,即得样地生物量干质量。把样地生物量(干质量)乘以碳量,即得样地碳密度,再乘以公顷倍数,即得公顷碳密度。
乔木层根生物量碳密度测定:以每块样地5株样木的伐根为中心,挖掘1.0 m×1.0 m×1.0 m的方坑和0.5 m×2.0 m×1.0 m的长坑,分层取根量,分为粗根和细根,去掉含水量即根量干质量,将每5株的方坑和长坑根量分别乘以样木组合株树,即得样木组合生物量,再把5株样木的组合生物量,即得样地生物量。再乘以碳量,即得碳密度。再乘以公顷倍数,即得根生物量公顷碳密度。
下木层生物量碳密度:下木层包括林下灌木和更新幼树,在样地中以对角线的分布,设置2.0 m×2.0 m的样方5块,采取全收割的方法测定地上部叶和枝的生物量及地下根量,去掉含水量,即得生物量各干质量,计算样方平均生物量,再乘以样地面积,即得样地生物量,再乘公顷倍数,即得公顷生物量,把公顷生物量乘以碳量,即得碳密度。
草本层生物量碳密度:在样地内以对角线的分布设置1.0 m×1.0 m的样方5个,采取全收割的方法,测定地上地下生物量,去掉含水量,计算样方生物量平均值,再乘以各组分碳量,即得碳密度。采用样地、样方法测定红松人工林生物量,计算碳密度,效果比用平均样木法和数学模型法更近实际[3]。
数据分析:对不同林龄型,不同组分生物量的样品,按着德国耶拿公司生产的碳氮仪3 000分析碳含量及生物量样品含水量,以便计算干质量。对不同林龄型样木的组合生物量进行多重比较,对不同生物量各组分生物量进行方差分析。
3 结果与分析
3.1 红松人工林群落层生物量碳密度比较
由表1说明,20年生3种林分类型乔木层生物量碳密度属柞红林最大,为31.70 t·hm-2,超过红纯林41.96%,与桦红林持平,42年生3种林分类型乔木层生物量碳密度属柞红林最大,超过红纯林10.11%,与桦红林持平,由上述分析,乔木层碳密度占生物量碳密度的90.23%~96.10%,占生物量碳密度的主体部分。
白桦红松林20年生林分下木层碳贮量为1.92 t·hm-2,柞红林下木层碳贮量为1.70 t·hm-2;42年生桦红林林分下木层碳贮量为2.681 t·hm-2,柞红林下木层碳贮量为2.304 t·hm-2。在红松纯林中,由于林内透光系数小,没有下木层分布,而在混交林中,下木层均有分布,构成群落层的组成部分。20和42 d群落下木层分别占总碳贮量5.58%~5.76%。20年生林分草本层生物量碳密度属桦红林草本层碳密度最多,为1.43 t·hm-2,超过柞红林70.23%,42年生林分属柞红林草本层碳密度最多,为1.42 t·hm-2,超过桦红林20.34%。由上述分析看出,红纯林林分没有草本层发生,因为林分郁闭度大,光环境弱,再加林内凋落层厚,而没有发生草本层,20年生桦红林草本层生物量碳密度等,因为林内光环境好,促进了草本层增加[4],相反,柞红林由于叶量繁茂,影响光照条件,抑制了草本层增加。在42年生柞红林,由于树体增加,林内稀疏,增加了光照条件,促进了草本层的生长,从而增加碳密度[5]。

表1 不同红松人工林群落层生物量碳密度比较
注:表中数据是由每块样地样木的平均值计算的;进行均方差和不同林龄碳密度的方差分析,差异显著(P<0.05)。红松纯林简称红纯林,白桦红松混交林简称桦红林,蒙古栎红松混交林简称柞红林。
3.2 不同林分类型对红松人工林生物量碳密度的影响
由表2看出,3种林分类型同一林龄的生物量碳密度,经方差分析,20年生红纯林、桦红林和柞红林的碳密度相差显著,F值为4.762,临界值为4.26,(P<0.05),以百分数比较,桦红林比红纯林大53.52%,柞红林比红纯林大53.29%,两种混交类型生物量碳密度相比,差异甚小,仅差0.15%,42年生桦红林生物量碳密度和红纯林生物量碳密度相比,大10.12%。柞红林大13.10%,2种混交类型碳密度相比,相差2.66%。
总之,两种混交林类型生物量碳密度均大于同龄的纯林,因为纯林凋落物为针叶,分解慢,生物自肥力弱,影响红松纯林生长,生物量较少,相反,混交林结构多层,生物多样性高[6],生物自肥能力强,促进林分生长,生物量比针叶纯林大,所以混交林生物量碳密度大。

表2 3种林分类型各组分碳密度比较(含下木、草本层)
注:表中数据是由不同林分类型样地样木实测值的平均值±标准差;进行均方差和各组分之间的方差分析,差异显著(P<0.05)。
3.3 不同林龄林分生物量碳密度比较
由表2看出,红纯林生物量碳密度42年生林分比20年生林分大276.71%,桦红林生物量碳密度42年生林分比20年生林分大169.66%,柞红林生物量碳密度42年生林分比20年生林分大177.91%,同时对上述不同林龄型的生物量碳密度均做了方差分析,结果F值为5.516,临界值F为5.32(P<0.05),差异显著。因为生物量碳密度是由生物量因子和碳含量的乘积构成,在碳含量一般差异不大的条件下,生物量因子在乘积中起主导作用,即生物量越大,则碳密度也随着增加。林木生物量的大小受林龄和经营水平制约,林龄越大生长量亦大,这是林木生长的规律,故林龄大的林分生物量碳密度自然增大,当然在同一林龄的林分,由于立地条件好,经营水平高,也会有差异的[7]。由此说明,在培育碳汇林应考虑成熟林,以使碳汇生产稳定增长[8]。
3.4 不同林分类型生物量组分碳密度比较
由表2看出,两种林龄,3种林分类型生物量组分碳密度由叶量、枝量、树干量、果量和根量构成,由于组分所处位置和功能不同,其碳密度相差显著,经方差分析,各组织器官贮碳量,即组分碳密度相差显著,F值为4.321,临界值为3.86(P<0.05)。同时,又做了各组织器官贮碳量占总贮碳量百分数的比较,结果说明,20年生红松纯林碳量百分数由大到小树干量、叶量、枝量、根量、果量,42年生红纯林碳量百分数由大到小为树干量、枝量、叶量、根量、果量,20年生桦红林碳量百分数由大到小为树干量、根量、叶量、枝量、果量;42年生桦红林碳量百分数由大到小为树干量、根量、叶量、枝量、果量;20年生柞红林碳量百分数由大到小为树干量、根量、叶量、枝量、果量;42年生柞红林碳量百分数由大到小为树干量、根量、叶量、枝量、果量。
由上述分析看出,树干生物量碳密度占最多比例,其百分比为48.76%~62.34%,无论哪种林龄和林型均占最多,因为树干的生物量占树木各组织器官生物量中最多,是林木的主体部分,而碳密度也随着增多,即生物量越大,则贮碳密度也不断增加,几乎是成正比的趋势发展。
其次,根量贮碳量也比较多,仅次于树干部分,根量靠着导管和管胞,具有从森林土壤中吸收水分和无机盐的功能,混交林根系发达,功能强,生物量较大,因此根碳密度占较多比例,除20年生红松根量碳密度偏少外,其余均占11.81%~21.78%,是贮碳的较多器官。
叶量和枝量的贮碳功能,具有相互依存的作用。叶多,吸碳潜力大,支持枝的生长,而枝的生长又能促进叶的展开和分布,使叶量增多,概括认为叶量贮碳量占总量12.78%~18.63%,枝量贮碳量占总量10.35%~14.71%,叶量和枝量贮碳量虽然没树干和根量贮碳量多,但其功能是很重要的,是不可缺少的贮碳器官,应予培育,调整最佳状态,从而提高碳汇产量[9]。果量贮碳量最少,但碳含量高,随着林龄增大,结果还会逐渐增加,也是贮碳的有效器官。
3.5 不同生物量组分碳密度在树体中的空间变化
叶碳密度的空问变化:由表3看出不同林龄、不同林分类型一年生叶多分布在树冠中层,少量分布在下层和上层,由表3说明,多年生叶碳密度,42年生红纯林、柞红林均分布在林冠下层,少量分布上层和中层,20年生红纯林分布在下层、桦红林、柞红林分布在中层,少量分布上层和下层。因为一年生叶中层冠幅大,生物量、碳密度自然增大。与此相反,多年生叶碳密度分布在下层。而20年生的一年生叶多分布在中层,多年叶分布在中层。枝碳密度的空间变化:由表4看出,42年一年生枝碳密度主要分布在中层部位,其次为上层和下层,多年生枝碳密度分布随树种而异,红松纯林主要在下层和中层,白桦红松混交林分布在中层,红松柞树混交林分布在下层,总的说,一年和多年生枝的碳密度主要分布在中层和下层,上层较少,其变化随枝生物量的分布而分布。因为生物量是碳密度的主导因子,是物质基础,在碳含量相差不大的前提下,生物量大则碳密度亦大[10]。反映碳密度的组成因素特点。
由表4分析出,20年生枝碳密度主要分布于中下层部位,而上层较小,与42年生枝碳密度分布相似,但幼龄林自然稀疏比中龄林较弱,中下层活枝分布多,生物量大,所以碳密度较大。反映了枝的自然特点。

表3 3种林分类型不同树种的林龄叶碳密度
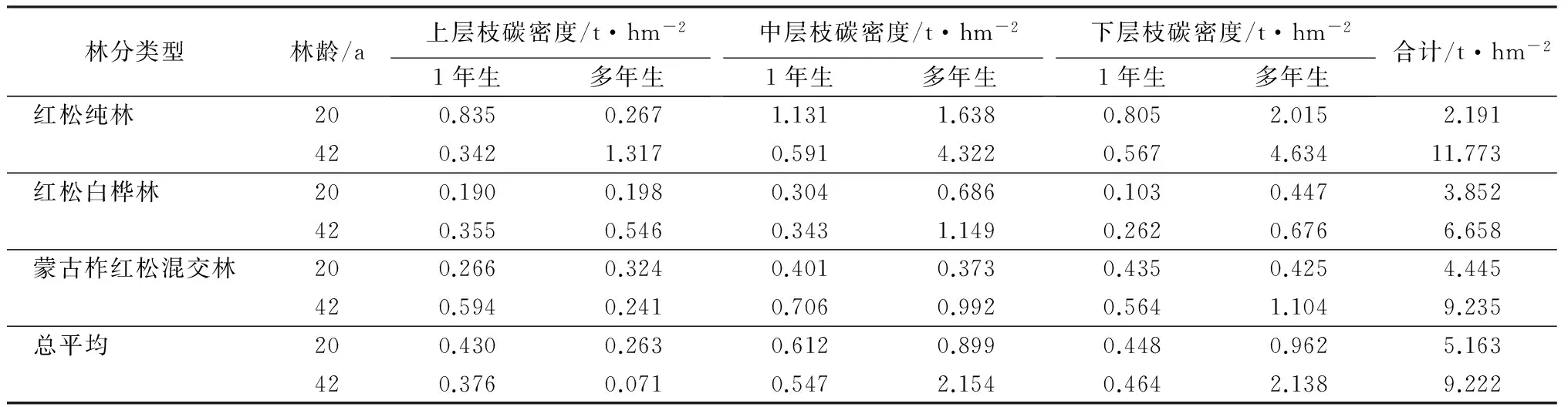
表4 3种林分类型不同林龄林各层分枝碳密度
树干碳密度空间的变化:由表5看出,42年和20年的红松人工林树干1 m区分段的碳密度随树干高度增加而降低,不同的类型和树种碳密度分布有所不同,红松树种在纯林或混交林中碳密度均比白桦和蒙古柞较高,但42年生从第6段,20年生从第5段以后,各树种碳密度分布接近一致,因为越上部树干径级细,碳密度接近一致。
根碳密度空间的变化:由42年生细根、20年生细根及42年生粗根、20年生粗根(见表6),看出42年生和20年生不同红松人工林类型根的碳密度随土层深度加深而减少的规律,呈递减的趋势。这种变化规律是随根的生物量分布而分布的,生物量越多,则碳密度越大,由此可以说明,根的碳密度主要分布在0~20 cm土层中,而以下的土层根系碳密度是很小的,红松纯林0~20 cm的碳密度比60~80 cm的碳密度大96.258%,红松柞树混交林大79.564%,差异是显著的,由此说明,根系的储碳能力主要分布在表层,而较深土层是很小的。0~20 cm的碳密度比60~80 cm的碳密度大79.56%,差异是显著的,由此说明,根系的贮碳量主要分布在土壤的表层,而较深的土层是很少的。

表5 不同林龄树干碳密度

表6 不同林龄根碳密度
4 结论与讨论
本文研究地区原始植被为阔叶红松林,在历史上曾遭严重破坏,建国后经过封山育林,荒山造林,天然次生林经营等多种措施,已逐渐演替成红松人工林和天然次生林混交的地带性植被,研究该植被生态系统生物量碳密度具有代表性,为培育碳汇林提供参考。
红松人工林及其混交林生物量碳密度是生物量碳库的主要指标,由树体贮碳器官叶、枝、树干、果和根的构成,具有吸碳、贮碳和转移碳的功能,与土壤、凋落物碳库比较,属基础碳库。生物量碳密度由生物量与碳含量两项因素乘积构成,在碳含量一定幅度内生物量起主导作用,即生物量越大,则乘积越大,碳密度随着增大,因此培育森林应以增加生物量为主要目标,促进碳汇生产。同时,在研究生物量碳密度的过程中,较深入认识到,红松人工林及其混交林的碳密度和碳汇生产,是构成有机体的基本成分,并且与有机体形成互为依存,互为条件,不可分割的统一体,这个统一体是再生资源,是有序的,有结构有功能的有机体,而通过自组织过程,达到新的平衡,进入稳定阶段,如果碳汇离开了有机体,就成为大气中的碳源,只有贮存于有机体中才能起到温室减排的作用,达到减缓气候变暖的目的。基于上述分析,只有经营好森林,实质就是经营好碳汇,才会增加森林碳汇。
两种林龄,3种林分类型,其碳密度相差显著,42年生红松纯林碳密度比20年生红松纯林碳密度大277.16%,42年生白桦红松混交林比20年生白桦红松混交林碳密度大169.69%,42年生蒙古栎红松混交林比20年生蒙古栎红松混交林大177.91%,因为林龄大的林分生物量亦大,碳密度随着增大。在同一林龄,3种林分类型碳密度比较,20年生白桦红松混交林碳密度比20年生红松纯林碳密度大53.52%,20年生蒙古栎红松混交林碳密度比20年生红松纯林碳密度大53.39%。42年生白桦红松混交林比42年生红松纯林大11.59%,42年生蒙古栎红松混交林比42年生红松纯林大12.95%。因为混交结构合理,养分丰富,能充分利用空间环境,生长量大,故混交林生物量比纯林大,碳密度自然增加。
经过计量,红松人工林生态系统生物量碳密度20年生为22.33~34.28 t·hm-2,42年生为84.12~95.13 t·hm-2;其中20年生叶量碳密度占总量12.52%~18.63%,枝量占10.35%~14.13%,树干量占48.76%~62.34%,果量占0.04%~0.06%,根量占6.14%~21.78%,以百分比多少排序为,树干量、根量、叶量、枝量、果量。其中树干量碳密度占一半以上是最多的,最少量为果量碳密度,因为树干量是林木的主体,生物量大,故碳密度增大,建议培育生态碳汇林。应着重树干量培育,从而增加生物量碳密度和碳汇生产。果量碳密度很小,只占总量1.51%,因为红松纯林和混交林,尚未达到成熟期,结实量有限,但碳含量高,随着林龄增大,结实会逐渐增加。
[1] 夏鑫.马尾松人工林的老龄林生态系统生物量和碳贮量研究[D].福州:福建农林大学,2008.
[2] 周玉荣,于振良,赵士洞.我国主要森林生态系统碳贮量和碳平衡[J].植物生态学报,2000,24(5):518-522.
[3] 王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究[J].应用生态学报,2001,12(1):13-16.
[4] XU M, QI Y. Soil-surface CO2efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California[J]. Global Change Biology,2001,7(6):667-677.
[5] KANG S Y, DOH S, LEE D, et al. Topographic and climatic controls on soil respiration in six temperate mixed-hardwood forest slopes, Korea[J]. Global Change Biology,2010,15(10):1427-1437.
[6] GIARDINA C P, RYAN M G. Evidence that decomposition rates of organic carbon inmineral soil do not vary with temperature[J]. Nature,2000,404:858-861.
[7] REY A E, PEGORARO V, TEDESCHI I, et al. Annual variaation in soil respiration and its components in a coppice oak forest in Central Italy[J]. Global Change Biology,2002,8(9):851-866.
[8] MARTIN J L, GOWERS T, PlAUT J, et al. Carbon pools in a boreal mixed wood logging chronosequence[J]. Global Change Biology,2010,15(11):1883-1894.
[9] 王绍强,周成虎,刘纪远.东北地区陆地碳循环平衡模拟分析[J].地理学报,2001,56(4):390-400.
[10] 郑帷婕,包维楷,辜彬.陆生高等植物碳含量及其特点[J].生态学杂志,2007,26(3):307-313.
