降水格局对科尔沁沙地三种草本植物凋落物分解的影响
2018-03-04罗永清岳祥飞李玉强赵学勇丁杰萍闫志强中国科学院西北生态环境资源研究院甘肃兰州70000中国地质科学院岩溶地质研究所广西桂林500甘肃省环境科学设计研究院甘肃兰州7000兰州交通大学环境与市政工程学院甘肃兰州70070
罗永清,岳祥飞,李玉强,赵学勇,丁杰萍,闫志强(.中国科学院西北生态环境资源研究院,甘肃 兰州 70000;.中国地质科学院岩溶地质研究所,广西 桂林 500;.甘肃省环境科学设计研究院,甘肃 兰州 7000;.兰州交通大学环境与市政工程学院,甘肃 兰州 70070)
水分是干旱半干旱区陆地生态系统植物生长的限制性因子,也是影响我国干旱和半干旱区植被特征的关键因素。以往的研究多以年均降水量作为因子进行分析[1],如我国内蒙古半干旱草地的研究表明[2],年均降水量与地上植被生产力呈线性正相关关系;内蒙古锡林河流域羊草(Leymuschinensis)草原净初级生产力的研究同样发现[3],地上生物量与年均降水量呈明显的线性正相关关系。有关土壤呼吸的研究也有类似的结论[4]。但也有研究显示[5],年降水量与生态系统过程并非简单的线性关系,甚至有研究表明[6],年降水量不能很好地解释初级生产力、土壤呼吸等生态过程的年际变化。其原因可能是由于关键生态变化的产生与降水可能存在非同步性,如植被生长与土壤呼吸主要发生在生长季,非生长季的降水量可能对分析结果产生一定的影响。另一方面,生长季的降水格局也是影响生态过程的重要因素,如单次降水大小、降水间隔等对土壤和植被均有重要影响[7]。
凋落物分解是陆地生态系统植物体向土壤和大气环境中返还养分和碳元素的主要途径[8]。凋落物包括地上凋落物和地下凋落物,地上凋落物是由植物地上器官脱落后形成,地下凋落物主要来源于植物根系的死亡与根表脱落物的产生。对于一年生植物而言,生长季结束后的地上部分最终全部转化为地上凋落物,地下根系全部死亡形成地下凋落物。地上凋落物与地下凋落物在植物-土壤物质周转过程中均发挥着重要作用,如在森林生态系统[9],地上凋落物在土壤表层的积累和分解被认为是影响土壤物质累积和林下植被分布的主要因素。而在农田或沙地等生态系统[10],一方面,由于地上植被受风力搬运等环境因素和放牧、刈割等人类活动影响,地上凋落物向环境的返还受到不同程度的制约,另一方面,植物根系具有周转迅速、时间和空间上的连续性等特征,地下凋落物对土壤物质积累意义重大[8]。如农田生态系统的研究表明,通过植物根系向土壤的物质返还量甚至高于地上部分[11]。凋落物分解主要受凋落物质量、气候和土壤等因素影响。气候因素方面,气温和降水是影响凋落物分解的主要因素。已有的研究主要集中在温度和总降水量对凋落物分解的影响等方面[12],而对不同降水格局方面的研究未见报道。因此,本研究选取科尔沁沙地轻度退化草地中的3种常见一年生草本植物作为研究对象,基于多年降水特征设置不同降水格局处理,对供试物种地上和地下凋落物分解率进行了研究,旨在探索不同降水格局下草本植物的分解特征,为了解沙地物质循环对降水格局的响应过程和沙地可持续恢复机理提供理论支撑。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古自治区东部的通辽市奈曼旗境内(N 42°55′, E 120°42′)。该区位于科尔沁沙地中南部,平均海拔340~350 m,属温带大陆性半干旱气候,冬春季干旱,多大风天气,大风日数20~60 d,夏季多雨,降水主要集中在6-8月,年降水量364.6 mm,为典型的雨热同期型气候。地貌以平缓的流动沙丘、半流动沙丘、固定沙丘、丘间低地和甸子地镶嵌分布为特征。优势灌木为差巴嘎蒿(Artemisiahalodendron)、黄柳(Salixgordejevii)和小叶锦鸡儿(Caraganamicrophylla),草本优势植物包括黄蒿(Artemisiasacrorum)、狗尾草(Setariaviridis)、尖头叶藜(Chenopodiumacuminatum)、砂蓝刺头(Echinopsgmelini)、冷蒿(Artemisiafrigida)、糙隐子草(Cleistogenessquarrosa)、地锦(Euphorbiahumifusa)、苦荬菜(Ixetisdenticulate)、雾冰藜(Bassiadasyphylls)等。
1.2 试验方法
1.2.1样品采集与试验布设 2014年8月底采集凋落物样品,采样区为3种供试物种分布频度较高的轻度退化和 未退化草地。地上凋落物采用整株收割法采集;收割后进行地下凋落物样的采集,地下凋落物分0~10 cm层和10~20 cm层采用挖掘法进行,挖掘后将带有根样的土块放置于筛网中(网孔直径1 mm)用自来水冲洗,弃去泥沙后手动挑出非目标植物根系。每个物种共计获取凋落物样品3份(1份地上样品和2份地下样品),样品带回实验室后进一步用去离子水冲洗干净,然后65 ℃烘干48 h。样品烘干后剪成长度2~3 cm的小节,然后装入网袋(长宽均为10 cm)中,每袋装入干样5.00 g。
分解试验在透光率良好、四周敞开的遮雨棚(10 m长×5 m宽×3 m高)中进行。2015年4月初用直径36 cm、高45 cm的PVC管在轻度退化草地进行原状土芯采集,采集深度40 cm(保留5 cm的上沿),采集后转移至遮雨棚中预先挖好的土坑中,保证PVC管上沿与遮雨棚地面保持一致。4月15日进行分解样品的埋设,具体操作为:在每个PVC管中先挖出深度20 cm的土坑,然后依次将10~20 cm根样和0~10 cm根样埋入土坑中,回填所有挖掘出来的土壤,保证紧实度与原状土一致,然后在表层水平放置地上凋落物并用倒“U”型细铁丝固定。
1.2.2实验设计与处理 凋落物样埋入土壤中缓冲2周后于4月29日开始处理。根据研究区历史降水特征(1961-2014年5-9月中降水量280 mm左右),本次研究设置3个处理,即高频率-低强度降水(T1)、中频率中强度降水(T2)和低频率高强度降水(T3)3个处理。其中T1每5 d人工模拟降水一次,单次降水10 mm,T2每10 d降水一次,单次降水20 mm,T3每20 d降水一次,单次降水40 mm,试验从4月29日持续至9月10日,3个处理分别降水28,14和7次,累计降水量均为280 mm。
9月底结束本次分解实验,将各分解袋收集后带回实验室,分别倒出各分解袋中的样品,用自来水和去离子水分别冲洗干净后65 ℃烘干至恒重,然后计算分解率。
1.3 数据分析
供试物种的分解率采用如下公式计算:
分解率=(M0-Mt)/M0×100%
式中:M0为凋落物的初始质量;Mt为分解后的质量。
数据采用Microsoft Excel 2010和SPSS 19.0软件进行统计分析。用三因素方差(three-way ANOVA)分析不同物种、降水格局和凋落物分解率的差异性,方差分析后在0.05水平下用最小显著性差异(LSD)进行多重比较,采用Sigmaplot 12.5作图。
2 结果与分析
2.1 地上凋落物分解特征
研究表明,不同降水格局对地上凋落物的分解有显著的影响(图1),其中对于狗尾草和黄蒿,低频高量降水处理下的分解率显著低于其他两种处理,高频低量和中频中量处理间的差异均不显著。而对于尖头叶藜地上凋落物的分解,3种处理间的差异均达到显著水平,分解率随降水频率的减少显著下降。另外,不同物种间的分解率也表现出差异性,各处理下黄蒿的分解率均高于狗尾草和尖头叶藜,3种处理下的分解率分别为42.60%、43.17%和33.00%;狗尾草的分解率最低,3种处理下的分解率分别为26.40%、27.50%和17.00%。
2.2 地下凋落物分解特征
不同降水格局同样对地下凋落物的分解率产生影响,这种影响与地下凋落物埋深和物种类型有关。不同物种间,狗尾草的分解率随降水频率降低逐渐下降,尤其是0~10 cm层,这种差异达到显著水平;黄蒿的0~10 cm层地下凋落物分解率在3种处理下差异性也达到显著水平,低频率降水处理下的分解率显著低于中频率降水处理,而高频低量处理下的分解率与其他两个处理间的差异性不显著;而尖头叶藜地下凋落物分解率对不同降水格局处理的差异性不显著。不同深度凋落物分解率的分析表明,凋落物埋深也是影响其分解率的重要因子,这种影响在不同物种间表现出差异性。具体表现为:狗尾草10~20 cm层的分解率在各处理下均显著高于0~10 cm层,3种处理下10~20 cm层的分解率分别高出0~10 cm层18.75%、28.40%和67.98%;黄蒿在高频低量和高量低频处理下10~20 cm层分解率高于0~10 cm层,中频重量下0~10 cm层的分解率高于10~20 cm层,但差异性不显著;尖头叶藜在3种处理下的地下凋落物分解率均表现为10~20 cm层高于0~10 cm层的特征,但仅在低频高量处理下的差异性达到显著水平(图2)。
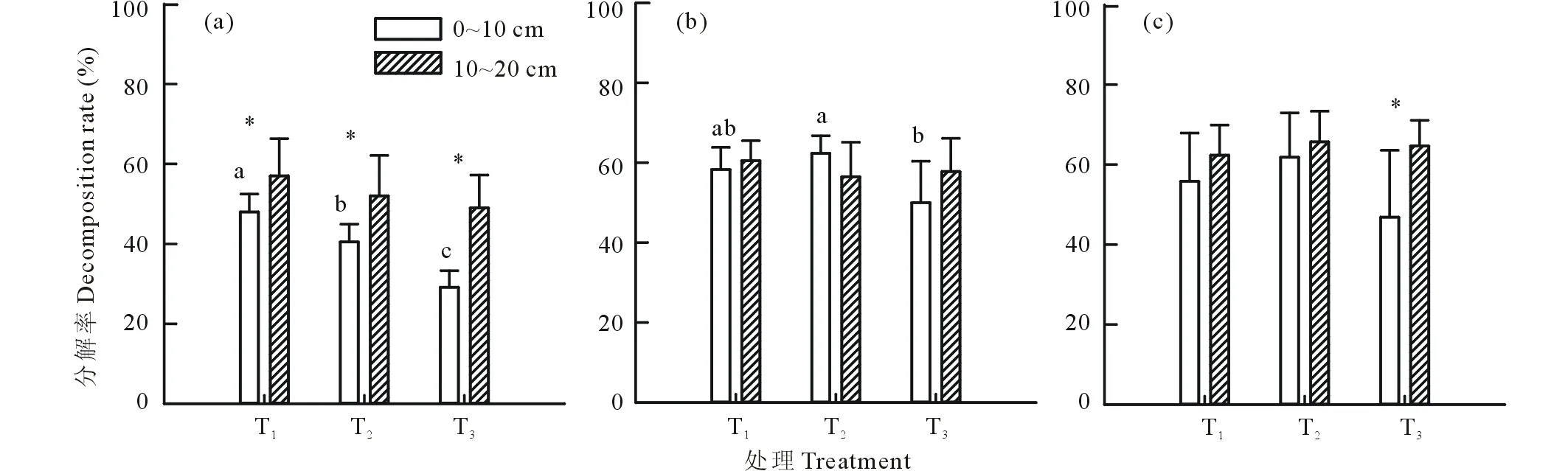
图2 不同降水格局处理下狗尾草(a)、黄蒿(b)和尖头叶藜(c)地下凋落物在0~10 cm埋深和10~20 cm埋深的分解率Fig.2 Plant belowground litter decomposition rate of S. viridis (a), A. sacrorum (b) and C. acuminatum (c) under treatments of different precipitation pattern at the depth of 0-10 cm and 10-20 cm不同小写字母表示差异显著(P<0.05); *表示显著性水平分别为P<0.05。The different letters mean significant difference at P<0.05; * indicate the significant level at P<0.05.
地上和地下凋落物分解率的对比发现,3种草本植物在不同降水格局处理下地上凋落物的分解率均显著低于地下凋落物,3种植物地上凋落物3种降水格局处理的平均分解率为地下凋落物(0~10 cm层和10~20 cm层)平均分解率的52.17%(狗尾草)、68.97%(黄蒿)和51.67%(尖头叶藜)。另外,三因素方差分析表明(表1),降水格局、凋落物埋深(地上凋落物视作凋落物埋深处理的一种)和物种类型均是影响科尔沁沙地一年生草本植物凋落物分解率的重要因素。此外,降水格局与凋落物埋深、物种类型与凋落物埋深均存在二因素交互作用。
3 讨论
凋落物分解是陆地生态系统物质循环的重要过程,凋落物分解率受本底质量和环境因素共同影响。水分和温度被认为是影响凋落物分解的主要驱动因子[8,11-13]。如土壤含水率与凋落物分解的相关研究表明[14],二者具有类似的特征,即凋落物分解率随土壤含水率的增加而提高。而已有的关于降水量和凋落物分解的研究表明,并非降水量的高低决定着凋落物的分解速率。如李雪峰等[15]的研究表明,年降水量450 mm处理下的分解率显著高于600和850 mm处理。土壤水分含量的高低除了受降水量影响以外,还与土壤质地、降水频次等因素有关。另外,降水和温度对凋落物分解和土壤呼吸的影响并非加和关系,而是更复杂的耦合关系。已有的分析表明[13],相对气候因子与凋落物分解的相关性更强。由此推断,在水热状况良好的科尔沁沙地,凋落物的分解可能主要发生在生长季。如在科尔沁沙地以黄柳和小叶锦鸡儿的研究表明[16],9月至次年5月的分解率仅为10%,而次年5月至9月的分解率分别为23.34%和21.08%。该分解率低于本次研究的结果,导致该差异的原因可能是分解起始时间的不同,本次研究起始于4月底,此时较高的土壤温度结合各种处理的土壤水分促进了凋落物的分解。
有研究表明[16-17],植物地上部分的分解率低于地下部分,本研究结果与此一致。一般认为导致该差异的原因是初始质量差异,即枝条的木质素等难分解物质含量高于根系[8,12,15]。刘文丹等[17]对比研究了粗根、细根、枝和叶片的分解率,结果表明,无论是石灰岩基质还是砂岩基质,常绿植物和落叶植物的枝和叶片的分解率均低于粗根和细根。普遍认为植物叶片的质量高于枝、根系等器官,但该研究结果显示叶片分解率低于根系。据此推断,土壤中适宜的微生物环境和较好的水分状况可能是导致地下凋落物分解率高于地上凋落物的重要原因。因此,在凋落物分解和植物-土壤系统物质循环方面的研究中,凋落物的分解基质是一个不可忽略的因素。如地下凋落物的分解主要受栖息于土壤中的微生物影响,而位于地表的地上凋落物除受地表土壤微生物和空气中的微生物影响以外,光照[18]也在一定程度上可促进凋落物的分解。
降水格局被认为是影响生态系统变化的主要因素。近年来的研究表明,无论从全球尺度[19]还是区域尺度[7],降水呈现总量年际波动较小、年内变异增大的趋势,极端降水和极端干旱事件频发[19]。因此,用总降水量来评价陆地生态过程变化的准确性降低。近年来一些研究逐渐开展了降水格局方面的研究,如极端干旱/降水、增减雨等方面的研究已有报道[19]。但目前的研究多集中在土壤呼吸、土壤微生物、植物个体发育与植被组成变化等方面,不同降水格局下凋落物分解方面的研究相对较少[20]。有关凋落物分解与降水方面的研究主要为大尺度多区域间凋落物分解差异的比较,在该过程中多以年均降水量或年总降水量作为因子进行分析[21]。而植物凋落物一方面是陆地生态系统物质循环和能量流动的关键环节,另一方面其分解率受降水影响,因此,开展不同降水格局对凋落物分解影响的研究可为准确预测区域尺度上凋落物分解和植被-土壤系统物质循环提供重要理论基础。
4 结论
作为陆地生态系统物质循环和能量流动的关键环节,凋落物分解受降水格局和凋落物埋深的影响,高频率和中频率降水格局下狗尾草、黄蒿和尖头叶藜的地上和地下凋落物分解率显著降低。降水格局对凋落物分解的影响与凋落物埋深存在耦合关系,该影响主要发生在地上凋落物和0~10 cm层地下凋落物。位于土壤表层的地上凋落物分解率低于位于土壤中的地下凋落物,在凋落物分解和植物-土壤系统物质循环方面的研究中,凋落物的分解基质是一个不可忽略的因素。
References:
[1] Epstein H E, Burke I C, Lauenroth W K. Regional patterns of decomposition and primary production rates in the U. S. great plains. Ecology, 2002, 83(2): 320-327.
[2] Ma W H, Yang Y H, He J S,etal. Above-and belowground biomass in relation to environmental factors in temperate grasslands, Inner Mongolia. Science China Life Sciences, 2008, 51(3): 263-270.
[3] Yuan F, Han X G, Ge J P,etal. Net primary productivity ofLeymuschinensissteppe in Xilin River basin of Inner Mongolia and its responses to global climate change. Chinese Journal of Applied Ecology, 2008, 19(10): 2168-2176.
袁飞, 韩兴国, 葛剑平, 等. 内蒙古锡林河流域羊草草原净初级生产力及其对全球气候变化的响应. 应用生态学报, 2008, 19(10): 2168-2176.
[4] Zhou X H, Talley M, Luo Y. Biomass, litter, and soil respiration along a precipitation gradient in southern Great Plains, USA. Ecosystems, 2009, 12(8): 1369-1380.
[5] Hu Z M, Fan J W, Zhong H P,etal. Spatiotemporal dynamics of aboveground primary productivity along a precipitation gradient in Chinese temperate grassland. Science China Earth Sciences, 2007, 50(5): 754-764.
[6] Yang Y, Fang J, Fay P A,etal. Rain use efficiency across a precipitation gradient on the Tibetan Plateau. Geophysical Research Letters, 2010, 37(15): 78-82.
[7] Yue X F, Zhang T H, Zhao X Y,etal. Effects of rainfall patterns on annual plants in Horqin Sandy Land, Inner Mongolia of China. Journal of Arid Land, 2016, 8(3): 389-398.
[8] Luo Y Q, Zhao X Y, Wang T,etal. Plant root decomposition and its responses to biotic and abiotic factors. Acta Prataculturae Sinica, 2017, 26(2): 197-207.
罗永清, 赵学勇, 王涛, 等. 植物根系分解及其对生物和非生物因素的响应机理研究进展. 草业学报, 2017, 26(2): 197-207.
[9] Tu Y, You Y M, Sun J X. Effects of forest floor litter and nitrogen addition on soil microbial biomass C and N and microbial activity in a mixedPinustabulaeformisandQuercusliaotungensisforest stand in Shanxi Province of China. Chinese Journal of Applied Ecology, 2012, 23(9): 2325-2331.
涂玉, 尤业明, 孙建新. 油松-辽东栎混交林地表凋落物与氮添加对土壤微生物生物量碳、氮及其活性的影响. 应用生态学报, 2012, 23(9): 2325-2331.
[10] Luo Y Q, Zhao X Y, Ding J P,etal. Dynamics of aboveground biomass and litters in different types of dunes under vegetation restoration processes in the Horqin Sandy Land. Journal of Desert Research, 2016, 36(1): 78-84.
罗永清, 赵学勇, 丁杰萍, 等. 科尔沁沙地不同类型沙地植被恢复过程中地上生物量与凋落物量变化. 中国沙漠, 2016, 36(1): 78-84.
[11] Bolinder M A, Kätterer T, Andrén O,etal. Estimating carbon inputs to soil in forage-based crop rotations and modeling the effects on soil carbon dynamics in a Swedish long-term field experiment. Canadian Journal of Soil Science, 2012, 92(6): 821-833.
[12] Guo J F, Yang Y S, Chen G S,etal. A review on litter decomposition in forest ecosystem. Scientia Silvae Sinicae, 2006, 42(4): 93-100.
郭剑芬, 杨玉盛, 陈光水, 等. 森林凋落物分解研究进展. 林业科学, 2006, 42(4): 93-100.
[13] Luo Y Q, Zhao X Y, Li Y Q,etal. Root decomposition ofArtemisiahalodendron, and its effect on soil nitrogen and soil organic carbon in the Horqin Sandy Land, northeastern China. Ecological Research, 2016, 31(4): 535-545.
[14] Zhang C H, Zhang L M, Liu X R,etal. Root tissue and shoot litter decomposition of dominant speciesStipabaicalensisin Hulunbuir meadow steppe of Inner Mongolia, China. Chinese Journal of Plant Ecology, 2011, 35(11): 1156-1166.
张彩虹, 张雷明, 刘杏认, 等. 呼伦贝尔草甸草原优势种贝加尔针茅根系组织和地上部分凋落物的分解. 植物生态学报, 2011, 35(11): 1156-1166.
[15] Li X F, Han S J, Zhang Y. Indirect effects of precipitation on litter decomposition ofQuercusmongolica. Chinese Journal of Applied Ecology, 2007, 18(2): 261-266.
李雪峰, 韩士杰, 张岩. 降水量变化对蒙古栎落叶分解过程的间接影响. 应用生态学报, 2007, 18(2): 261-266.
[16] Qu H, Zhao X Y, Zhao H L,etal. Litter decomposition rates in Horqin Sandy Land, Northern China: Effects of habitat and litter quality. Fresenius Environmental Bulletin, 2011, 20(12): 3304-3312.
[17] Liu W D, Tao J P, Zhang T D,etal. Decomposition of above and belowground organ litters of mid-subtropical woody plants. Acta Ecologica Sinica, 2014, 34(17): 4850-4858.
刘文丹, 陶建平, 张腾达, 等. 中亚热带木本植物各器官凋落物分解特性. 生态学报, 2014, 34(17): 4850-4858.
[18] Austin A T, Vivanco L. Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature, 2006, 442(7102): 555-558.
[19] Lemoine N P, Sheffield J, Dukes J S,etal. Terrestrial Precipitation Analysis (TPA): A resource for characterizing long-term precipitation regimes and extremes. Methods in Ecology and Evolution, 2016, 7(11): 1396-1401.
[20] Joly F X, Kurupas K L, Throop H L. Pulse frequency and soil-litter mixing alter the control of cumulative precipitation over litter decomposition. Ecology, 2017, 98(9): 2255-2260.
[21] Brandt L A, King J Y, Milchunas D G. Effects of ultraviolet radiation on litter decomposition depend on precipitation and litter chemistry in a shortgrass steppe ecosystem. Global Change Biology, 2007, 13(10): 2193-2205.
