黄土丘陵区草本群落生物量空间分布格局及其影响因素
2018-03-04方昭张少康刘海威焦峰1张军中国科学院水利部水土保持研究所陕西杨凌712100中国科学院大学北京10009西北农林科技大学水土保持研究所陕西杨凌712100华中农业大学公共管理学院土地管理学院湖北武汉0070
方昭,张少康,刘海威,焦峰1,*,张军(1.中国科学院水利部水土保持研究所,陕西 杨凌712100;2.中国科学院大学,北京10009;.西北农林科技大学水土保持研究所,陕西 杨凌712100;.华中农业大学公共管理学院,土地管理学院,湖北 武汉 0070)
草地生态系统作为陆地生态系统的重要组成部分,在调节全球气候变化、碳循环以及水土保持方面具有重要作用[1-2]。生物量作为草地生态系统生产力的重要衡量标准[3],反映了植物生长发育状况及其形态特征的变化。草地生物量大部分都集中在地下部分,而且地下生物量的动态变化和地上生物量的变化息息相关,二者互相影响[4]。因此,草地地上地下生物量的关系研究一直是草地生态学研究中的重要环节[5]。近些年来,随着全球气候变化加剧,草地生物量的研究越来越受到关注[6]。国内外学者相继开展了草地生物量的大量研究,其中许多学者运用异速生长理论对生物量分配进行估测,相关研究表明在个体和群落水平上地上生物量和地下生物量均符合等速生长关系[7-8]。相比地上生物量而言,地下生物量的测定工作较为困难,导致地下生物量的相关研究进展缓慢,对于大尺度上地下生物量的研究通常采用模型法[9]和根冠比(R/S)法[10]间接地进行估算,其估测值还存在很大的不确定性。在我国各类草地总面积约为4×108hm2,面积辽阔[11]。草地是干旱半干旱地区黄土高原植被组成的主体,占植被总面积的65.0%[12]。草本植物发达的根系为草地提供了强大的水土保持功能,使得退耕还林还草政策成为西部大开发战略实施以来黄土高原恢复植被、改善生态的重要措施之一[13]。黄土高原从东南到西北呈现出明显的地带性水热梯度变化特征[14],为研究草本群落生物量分布变化规律及其影响因素提供了理想试验场地。
本研究利用黄土丘陵区天然的水热梯度变化背景优势,集中探讨了黄土丘陵区不同纬度梯度下草本群落地上、地下生物量变化规律及其地下生物量对环境因子的响应规律。旨在:(1)揭示黄土丘陵区纬度梯度下草本群落地上、地下生物量变化规律;(2)在群落水平上检验等速生长理论;(3)讨论环境因素对地下生物量的影响,同时为黄土丘陵区植被恢复建设提供理论指导。
1 材料与方法
1.1 样地概况
在黄土丘陵区纬度梯度下,由南向北依次选取富县、甘泉县、安塞县、靖边县、横山县以及榆林市榆阳区6个典型地区作为研究区,并根据不同的纬度梯度和植被类型特征将其分为4个不同植被带:森林带、森林-草原带、草原带和草原-荒漠带。该研究区位于107.97°—109.87° E, 35.95°—38.36° N,海拔1085~1553 m,属于暖温带大陆性季风气候,平均气温8.8 ℃,年平均降水量505.3 mm,日照时数2395.6 h,无霜期157 d。主要土壤类型为黄绵土、绵沙土和风沙土,土质松软,加之地表植被稀疏、降水集中等特点,土壤侵蚀和水土流失现象较为严重。研究区(表1)以禾本科(Poaceae)、菊科(Asteraceae)以及豆科(Leguminosae)植物为优势草本植物,主要有长芒草(Stipabungeana)、达乌里胡枝子(Lespedezadavurica)、铁杆蒿(Ariemisiagmelinii)、沙蒿(Ariemisiadesertorum)、猪毛蒿(Ariemisiascoparia)、风毛菊(Saussureaamurensis)、阿尔泰狗娃花(Heteropappusaltaicus)等。

表1 调查样地基本概况Table 1 Basic information of the experimental plots
Ⅰ:黄绵土Loessial soil, Ⅱ:绵沙土Loess sandy soil, Ⅲ:风沙土Aeolian sandy soil.
1.2 研究方法
1.2.1样品采集和处理 野外采样于2012年8月中旬进行,此时处于草本植物生长旺盛期,生物量达到较大值。在该研究区内共选取15块样地,每块样地内设置6个1 m×1 m的样方进行草本植物群落特征调查,对样方内草本植物的种类、多度、盖度、高度和频数进行统计;同时采用传统收割法,将6个样方内所有草本植物地上部分剪下,装入纸质信封带回实验室。地下生物量采用内径为7 cm的土钻在每个样地内随机选取10个点采集地下根系部分,分层方法为0~10 cm、10~20 cm、20~40 cm,将样品编号后装入密封袋带回实验室处理。同时在每块样地上按照S形选取5个样点用土钻采集土壤样品,按0~10 cm、10~20 cm、20~40 cm分层均匀混合后装于密封袋内带回实验室风干处理。
1.2.2地上生物量测定 将采集的植物样品带回实验室后去除表面黏附土壤、砾石等杂物后,经105 ℃条件下杀青处理后(约10 min),再置于70 ℃烘箱中烘干至恒重后称重。
1.2.3地下生物量测定 将样品用清水充分浸泡,使其表面黏附的土壤和杂质沉淀,然后置于0.5 mm筛网中,用大量清水反复进行冲洗,晾干后于70 ℃烘箱中烘干至恒重后称重。
1.2.4土壤养分理化性质测定 将土壤样品中细根等杂物拣出后研磨过筛进行元素分析。土壤有机碳采用重铬酸钾热容量法测定;土壤全氮经H2SO4-K2SO4:CuSO4:Se催化后用凯氏定氮仪(型号:ATN-300)进行测定;土壤全磷经H2SO4-HClO4消煮处理后采用钼蓝比色法测定,具体测定方法均参照参考文献[15]。
1.3 气象数据和地形数据
用全球定位系统(GPS)获取各个样地的地形数据(经度、纬度、坡向和海拔),并从国家气象局获取陕西省各县1990-2010年的气象资料,根据样地坐标使用ArcGIS软件进行Kriging插值法[16]提取所有样地年均温和年降水量数据,取其20年的平均值作为样地的年均温和年降水量(图1)。

图1 样地的年降水量和年均温Fig.1 Mean annual precipitation and mean annual temperature of sampling sites
1.4 数据处理
在进行数据分析之前,对各类数据进行正态分布检验(单个样本K-S检验),检验结果以上数据均符合正态分布。采用SPSS 22.0 软件分析不同纬度梯度下草本群落地上、地下生物量的变化特征及其对不同植被带草本群落生物量(地下、地上生物量和地下/地上生物量)进行单因素方差分析,并将地下生物量与各环境因子(温度、降水、土壤有机碳、全氮、全磷)进行相关分析和回归分析。草本群落地上与地下生物量之间的相关性采用Ⅱ类回归方法(Reduced Major Axis,RMA)分析,等速生长检验、异速生长指数(b)和截距(loga)的计算均使用SMARTR软件完成[17]。
2 结果与分析
2.1 草本群落地上、地下生物量和地下/地上生物量纬度梯度变化
表2中给出了黄土丘陵区不同纬度梯度下草本群落地下、地上生物量及地下/地上生物量。在35.95°~38.36° N的研究范围内,草本群落地上生物量的变化范围为54.60~204.32 g·m-2,最大值出现在纬度约36.8° N的位置处,平均值156.968 g·m-2,变异系数为27.83%;草本群落地下生物量的变化范围为78.88~829.64 g·m-2,最大值出现在纬度约37.1° N的位置处,平均值469.21 g·m-2,变异系数为48.87%;草本群落地下/地上生物量变化范围为 0.93~4.49,平均值2.89,变异系数为39.18%。相关分析研究发现,草本群落地上、地下生物量随纬度梯度的变化规律均呈现出“先增加后减少”的趋势(图2),并不是简单的线性关系。
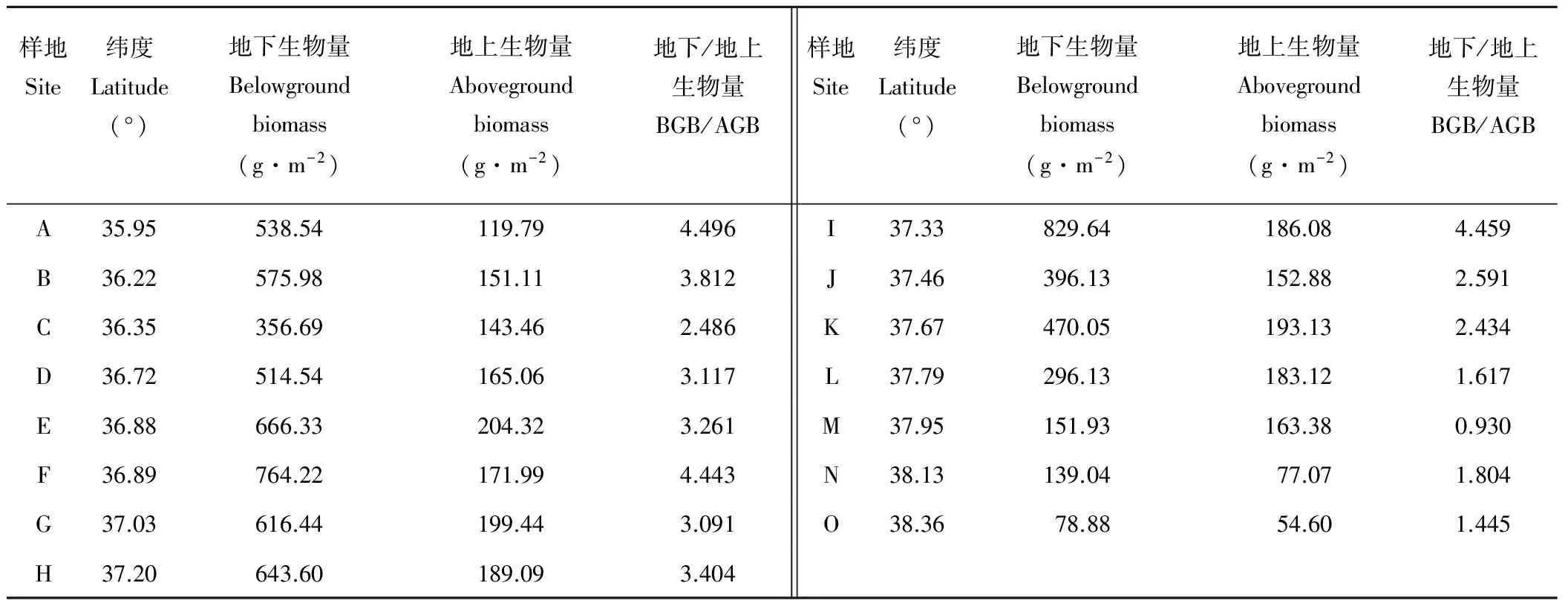
表2 草本群落地上生物量(AGB)、地下生物量(BGB)和地下/地上生物量(BGB/AGB)Table 2 Herbaceous community aboveground biomass (AGB),belowground biomass (BGB) and below/aboveground biomass (BGB/AGB)

图2 草本群落生物量随纬度的变化规律Fig.2 Variation of the herbaceous community biomass along the latitudinal gradientI:森林带Forest zone. Ⅱ:森林-草原带Forest-grass zone. Ⅲ:草原带Grass zone. Ⅳ:草原-荒漠带Grass-desert zone.
从表3中还可以看出,研究区内4个不同植被带的平均草本群落地下生物量分别为496.44,672.65,497.99,123.28 g·m-2,最大值和最小值相差750.76 g·m-2;草本群落地上生物量分别为144.86,191.21,178.80,98.35 g·m-2,最大值和最小值相差149.72 g·m-2;草本群落地下生物量均大于地上生物量,地下生物量是地上生物量的0.930~4.496倍。草原-荒漠带生物量(地下、地上)相对较低,且与森林-草原带和草原带差异显著(P<0.05)。草本群落生物量(地下、地上)大小顺序均为:森林-草原带>草原带>森林带>草原-荒漠带。

表3 不同植被带草本群落地下、地上和地下/地上生物量方差分析结果Table 3 Variance analysis of BGB,AGB,BGB/AGB of herbaceous community in different vegetation zones
注:同列数据后不同字母表示不同植被带间差异显著达5%显著水平。
Note: Values followed by different letters in a column are significant among vegetation zones at the 5% level.
2.2 地下和地上生物量的相关生长关系

图3 地上生物量和地下生物量的关系Fig.3 Relationship between belowground biomass and aboveground biomass
黄土丘陵区草本群落地上生物量与地下生物量之间存在极显著的线性相关关系(P<0.01),决定系数达到0.59,所以可以利用地上生物量来间接地预算和估测黄土高原草地生态系统生物量的大小(图3)。根据地上-地下生物量模型可知,草本群落地上和地下生物量之间的关系可用方程y=axb来表示,将方程两边同时取对数后转变为log(y)=loga+blog(x),b表示相关性的斜率,loga表示截距。通过标准主轴法(standardized major axis,SMA)计算回归直线的斜率和截距。根据相关生长理论,植物的地上生物量和地下生物量呈等速生长关系(回归直线的斜率与1.0差异不显著)或异速生长关系(回归直线的斜率与1.0
差异显著)[7,9,17-18]。拟合结果为:草本群落地上、地下生物量水平拟合直线的斜率b为1.07573 (95%置信区间: 0.83685~1.31461),对斜率b进行等速生长检验(即与1.0进行比较)发现,与1.0没有显著差异(P>0.05), 表明在群落尺度上,黄土丘陵区草本群落地上生物量与地下生物量呈等速生长关系。
2.3 地下生物量的影响因素
草本群落地下生物量与年均温、年均降水之间均呈现显著相关关系(图4)。随着年均温和年均降水量的升高,草本群落地下生物量均表现为“先增加后减少”的变化趋势(R2=0.683,P<0.001;R2=0.658,P<0.001)。由表4可知,不同土层的土壤有机碳、全氮、全磷含量与地下生物量之间均存在显著相关关系。从分析结果还可以看出,草本群落地下生物量与表层(0~20 cm)土壤有机碳含量的显著性(P<0.01)要高于草本群落地下生物量与深层(20~40 cm)土壤有机碳含量的显著性(P<0.05),而由于土层深度变化引起的显著性降低在草本群落地下生物量与土壤全氮、全磷含量之间没有得到体现,他们之间均是呈现出较为一致的显著相关性(P<0.05)。

表4 草本群落地下生物量(BGB)与土壤因子的相关关系Table 4 Correlations between herbaceous community belowground biomass (BGB) and soil factors
*P<0.05,**P<0.01.

图4 地下生物量对气候因子的响应Fig.4 Responses of belowground biomass to climatic factors
3 讨论与结论
3.1 地下、地上生物量变化及其与水热因子的关系
黄土丘陵区面积辽阔,地貌类型复杂多变,植被分布由南向北总体呈现出森林向草原过渡的明显地带性差异变化。不同植被带草本群落物种组成不同,而且不同草本植物对环境因子的适应能力和响应程度也各异,从而造成不同植被带草本群落生物量存在明显差异。有研究表明,水热因子是制约干旱-半干旱地区黄土高原草本群落生物量的重要因素,因纬度梯度变化所造成的水热分布不均和土壤条件差异对草本群落生物量状况及其空间分布有重要影响作用[19-21]。黄土丘陵区不同植被带草本群落生物量由大到小依次为:森林-草原带>草原带>森林带>草原-荒漠带,且地上、地下生物量随纬度梯度变化规律均呈现出“先增加后减少”的单峰型变化趋势,这与韩彬等[22]采用样带法对内蒙古草地植物群落生物量的研究结果相一致。本研究区域从降水稀少的草原-荒漠带过渡到温暖湿润的森林植被带,水热因子梯度变化明显。干旱-半干旱地区植物受到最普遍的环境胁迫就是水分供应不足,水分作为生物体的重要组成部分参与着各种新陈代谢过程,影响着植物体的生长发育情况。温度也是植物生命活动中不可或缺的生态因素,一方面通过酶活性变化影响着植物体内各种代谢过程,例如影响水分与矿质的吸收,物质合成、转化、运输和分配,进一步影响细胞功能的发挥从而影响植物体的正常代谢活动;另一方面温度升高也会加速土壤水分的蒸发使植物生长发育受到水分胁迫而影响植物体生命活动。草原-荒漠带和草原带降水量少、温度低,草本群落受水分和热量的限制作用明显,生理生化反应过程缓慢,生长发育迟缓,同时受人类活动干扰的可能性大,使其生长受到抑制,导致草本群落地上、地下生物量较小;而随着纬度的增加逐渐过渡到森林-草原带和森林植被带,水热条件发生明显改善,温度的升高虽然会加速土壤水分蒸发不利于植物生长,但是降水量的增多对水分的限制起到了一定程度的缓解作用,加之森林-草原带和森林植被带凋落物较多,容易进入土壤转化为有机质促进植物生长,较适宜的土壤养分、水资源以及温度条件,共同促进了草本植物植株和根系的生长,故形成草本群落地上、地下生物量随纬度不断上升的变化趋势,而随着温度的继续升高导致土壤水分蒸发量大于降水量,进而使得黄土丘陵区草本植物的生长受土壤水分胁迫作用增强,不利于植物的地上部分和地下细根的生长,引起地上、地下生物量下降。此外,本研究对象限定为草本群落,而没有将森林植被带和森林-草原带中的乔、灌木考虑在内,这也可能是引起地上、地下生物量降低的主要原因,从而表现出与邓蕾等[23]研究结果不同的变化趋势。
3.2 地下生物量与地上生物量的相生生长关系
深入研究地上-地下生物量关系可以加深对草地生态系统碳分配与储存过程的认识[24],而且对于研究草地生态系统土壤碳输入和碳循环也有着重要意义。目前,关于地上-地下生物量的分配机制研究主要聚焦于等速生长和最优分配两种假说,等速生长假说认为植物地上部分和地下部分生长存在一种平衡,地上与地下生物量呈现出相对一致的变化趋势;而最优分配假说则认为植物可以通过调整其分配模式来适应不同胁迫环境变化,从而达到保证自身生长的目的。黄土高原地区面积辽阔,地跨半湿润、半干旱和干旱3个气候带,从东南到西北因地带性水热梯度变化依次形成森林草原、温性草原和荒漠草原草地植被。森林草原作为森林和草原地带的群落交错区,特点是在草甸草原背景上分布着少量的旱中生灌木和矮乔木;与森林草原不同,温性草原多以旱生草本植物为主;而荒漠草原作为最旱生类型,气候干燥、少雨的极端气候形成以旱生丛生小禾草为主,并伴有少量的强旱生小半灌木和半灌木的植被组成。水热因子梯度变化引起不同类型草地植被物种组成,植物生态适应能力以及繁殖策略上均存在明显差异,但植物的相关生长性在植物界却是普遍存在的。目前,国内外相关学者越来越倾向于支持等速生长假说,认为植物在不同环境条件下地上和地下部分生长存在一种平衡。Niklas[18]研究表明藻类、苔藓乃至被子植物和裸子植物均存在一个相对的分配方式。程栋梁[25]对中国西北地区森林群落地上地下生物量分配研究发现森林植物均符合等速生长模型,但相关生长常数具有较大差异。而近年来王敏等[26]对黑河中游荒漠草地地上和地下生物量的研究也发现荒漠植物无论在个体水平还是群落水平上同样符合等速生长关系,但也有研究表明并不是所有荒漠植物都符合等速生长关系[27]。荒漠草原特殊的气候环境促使荒漠植物可能减少纵向生长而增加横向生长,并且这种趋势会随着环境胁迫的加剧变得更加明显,因此荒漠草原常见植被组成多为耐旱、叶比面积小且根深的低矮植物种类。本研究RMA分析结果显示,群落水平拟合回归直线的斜率与1没有显著差异(P>0.05),表明在群落水平上,黄土丘陵区草本群落地上生物量与地下生物量分配格局符合等速生长关系,此结论与王敏等[26]的研究结论相一致。由于本实验研究所选取的样本量较小,草本群落结构简单,使得拟合结果(R2=0.59)远高于全球草地中的对应值0.12[10]。黄土丘陵区草本群落地上和地下生物量通过幂函数方程拟合效果较好,体现了利用易获取的地上生物量数据估测地下生物量大小的可行性。但目前,不同植物以及同种植物不同发育阶段的相生生长关系方面还缺乏系统全面的认识。因此,大量的相关实验研究来充分验证相生生长关系的合理性及其适用条件也应作为当前该领域的主要研究任务。
3.3 地下/地上生物量变化与地下生物量对土壤因子的响应
地下/地上生物量反映了植物体光合作用产物的分配比例,是植物为应对环境变化做出的适应策略。本研究中地下/地上生物量在森林植被带→森林-草原植被带的过渡阶段出现较大的峰值,该结果与韩彬等[22]关于地下/地上生物量随纬度变化的响应趋势相同,其原因可能与研究区所处植被类型、物种组成和水热因子的差异有关。关于地下/地上生物量的影响因素目前还没有统一明确的定论,结果因植被类型、环境梯度、研究尺度和研究方法等不同存在很大差别[28-30]。不同群落间草本群落地下/地上生物量分配过程往往是十分复杂的,因此深入研究不同环境因子对地下/地上生物量的影响对于准确估算我国草地地下生物量大小和碳储量具有十分重要的研究意义。地下根系生长动态与环境因子的关系密不可分,环境变化势必会对地下生物量产生重要影响。由于3.1中水热因子对草本群落生物量的影响机理已经做了详细阐明,故本小结将仅探讨地下生物量对土壤因子的响应规律。戴诚等[29]和杨秀静等[30]认为地下生物量与土壤有机碳和全氮含量呈显著正相关,本研究结果与此较为一致。土壤C、N的空间分布受气候因素和成土作用机制影响变化较大,一般而言,从表层向深层的过渡中土壤C、N含量呈现出逐渐降低的分布格局,表层土壤较强的养分供应能力对细根生长起到了明显的促进作用,所以地下生物量与土壤有机碳、全氮含量相关性随着土壤深度增加呈现出逐渐下降趋势。而磷作为一种沉积型元素空间变异性较小,但却在植物生命活动中起着非常重要的作用。生长在黄土丘陵区的植物群落多数会受磷元素的限制作用,而低磷胁迫可能会促使植物根系产生一系列复杂的形态学变化。由此可见,土壤环境与植物地下根系生长变化关系密切,息息相关。 综上研究,气候因子和土壤理化性质对地下生物量都具有重要影响作用,但还需进一步结合植物生物学和生理生态方面的综合研究才能揭示其影响机理,做出准确可靠的结论。
References:
[1] Scurlock J M O, Johnson K, Olson R J. Estimating net primary productivity from grassland biomass dynamics measurements. Global Change Biology, 2002, 8(8): 736-753.
[2] Zhao W N. Community Carbon Sequestration of Grassland in the Loess Hill Region and Relationship with Environmental Factors. Yangling: Northwest A&F University, 2014.
赵婉伲. 黄土高原丘陵区典型草地群落碳储量及与环境因子的关系. 杨凌: 西北农林科技大学, 2014.
[3] Wang Y L, Ma Y S, Shi J J,etal. Investigation of biomass and soil nutrition of different vegetation type at alpine meadow in yellow river headwater area. Acta Agrestia Sinica, 2011, 19(1): 1-6.
王彦龙, 马玉寿, 施建军, 等. 黄河源区高寒草甸不同植被生物量及土壤养分状况研究. 草地学报, 2011, 19(1): 1-6.
[4] Sun L A, Liang Y M, Liu G B. A review of underground biomass of grassland. Grassland and Turf, 1993, (1): 6-14.
孙力安, 梁一民, 刘国彬. 草地地下生物量研究综述. 草原与草坪, 1993, (1): 6-14.
[5] Hu Z M, Fan J W, Zhong H P,etal. Progress on grassland underground biomass researches in China. Chinese Journal of Ecology, 2005, 24(9): 1095-1101.
胡中民, 樊江文, 钟华平, 等. 中国草地地下生物量研究进展. 生态学杂志, 2005, 24(9): 1095-1101.
[6] Zheng X X, Zhao J M, Zhang Y G,etal. Variation of grassland biomass and its relationships with environmental factors in Hulunbeier, Inner Mongolia. Chinese Journal of Ecology, 2007, 26(4): 533-538.
郑晓翾, 赵家明, 张玉刚, 等. 呼伦贝尔草原生物量变化及其与环境因子的关系. 生态学杂志, 2007, 26(4): 533-538.
[7] Enquist B J, Niklas K J. Global allocation rules for patterns of biomass partitioning in seed plants. Science, 2002, 295(5559): 1517-1520.
[8] Cheng D L, Niklas K J. Above- and below-ground biomass relationships across 1534 forested communities. Annals of Botany, 2007, 99(1): 95-102.
[9] Niklas K J. Modelling below- and above-ground biomass for non-woody and woody plants. Annals of Botany, 2005, 95(2): 315-321.
[10] Mokany K, Raison R J, Prokushkin A S. Critical analysis of root∶shoot ratios in terrestrial biomes. Global Change Biology, 2006, 12(1): 84-96.
[11] Ni J. Carbon storage in grasslands of China. Journal of Arid Environments, 2002, 50(2): 205-218.
[12] Chen F H, Huang W, Jin L Y,etal. Characteristics and spatial variation of precipitation in arid regions of Central Asia under the background of global warming. Scientia Sinica, 2011, (11): 1647-1657.
陈发虎, 黄伟, 靳立亚, 等. 全球变暖背景下中亚干旱区降水变化特征及其空间差异. 中国科学(地球科学), 2011, (11): 1647-1657.
[13] Jiao F, Wen Z M, Li R. Analysis on environment effect of the returning farmland into forest and grassland on the loess plateau. Research of Soil and Water Conservation, 2005, 12(1): 26-29.
焦峰, 温仲明, 李锐. 黄土高原退耕还林(草)环境效应分析. 水土保持研究, 2005, 12(1): 26-29.
[14] Chen Y M, Liang Y M, Cheng J M. The zonal character of vegetation construction on loess plateau. Acta Phytoecologica Sinica, 2002, 26(3): 339-345.
陈云明, 梁一民, 程积民. 黄土高原林草植被建设的地带性特征. 植物生态学报, 2002, 26(3): 339-345.
[15] Bao S D. Soil Agrochemical Analysis. 3 edition. Beijing: China Agriculture Press, 2000.
鲍士旦. 土壤农化分析. 3版. 北京: 中国农业出版社, 2000.
[16] Wang Y N, Xie J M, Guo X. Application of geostatistical interpolation method in ArcGIS. Software Guide, 2008, (12): 36-38.
王艳妮, 谢金梅, 郭祥. ArcGIS中的地统计克里格插值法及其应用. 软件导刊, 2008, (12): 36-38.
[17] Warton D I, Wright I J, Falster D S,etal. Bivariate line-fitting methods for allometry. Biological Reviews of the Cambridge Philosophical Society, 2006, 81(2): 259.
[18] Niklas K J. A phyletic perspective on the allometry of plant biomass-partitioning patterns and functionally equivalent organ-categories. New Phytologist, 2006, 171(1): 27.
[19] Coupland R T. Grassland ecosystems of the world. London: Cambridge University Press, 1979.
[20] Ji W Y. The Study of Biomass, Belowground Productivity and Their Relationship with Environmental Factors in Inner Mongolia Grassland. Beijing: Beijing Normal University, 2013.
纪文瑶. 内蒙古草原生物量、地下生产力及其与环境因子关系研究. 北京: 北京师范大学, 2013.
[21] Ma W H, Yang Y H, He J S,etal. Inner Mongolia temperate grassland biomass and its relationships with environmental factors. Scientia Sinica (Vitae), 2008, 38(1): 84-92.
马文红, 杨元合, 贺金生, 等. 内蒙古温带草地生物量及其与环境因子的关系. 中国科学(生命科学), 2008, 38(1): 84-92.
[22] Han B, Fan J W, Zhong H P. Grassland biomass of communities along gradients of the Inner Mongolia grassland transect. Acta Phytoecologica Sinica, 2006, 30(4): 553-562.
韩彬, 樊江文, 钟华平. 内蒙古草地样带植物群落生物量的梯度研究. 植物生态学报(英文版), 2006, 30(4): 553-562.
[23] Deng L, Shang G Z P. Distribution of natural grassland biomass and its relationship with influencing factors in Shaanxi. Acta Agrestia Sinica, 2012, 20(5): 825-835.
邓蕾, 上官周平. 陕西省天然草地生物量空间分布格局及其影响因素. 草地学报, 2012, 20(5): 825-835.
[24] Cairns M A, Brown S, Helmer E H,etal. Root biomass allocation in the world's upland forests. Oecologia, 1997, 111(1): 1-11.
[25] Cheng D L. Plant Allometric Study of Biomass Allocation Pattern and Biomass Production Rates. Lanzhou: Lanzhou University, 2007.
程栋梁. 植物生物量分配模式与生长速率的相关规律研究. 兰州: 兰州大学, 2007.
[26] Wang M, Su Y Z, Yang R,etal. Allocation patterns of above- and belowground biomass in desert grassland in the middle reaches of Heihe River, Gansu Province, China. Chinese Journal of Plant Ecology, 2013, 37(3): 209-219.
王敏, 苏永中, 杨荣, 等. 黑河中游荒漠草地地上和地下生物量的分配格局. 植物生态学报(英文版), 2013, 37(3): 209-219.
[27] Yang H T, Li X R, Liu L C,etal. Biomass allocation patterns of four shrubs in desert grassland. Journal of Desert Research, 2013, 33(5): 1340-1348.
杨昊天, 李新荣, 刘立超, 等. 荒漠草地4种灌木生物量分配特征. 中国沙漠, 2013, 33(5): 1340-1348.
[28] Ma W H, Fang J Y. RBS ratios of temperate steppe and the environmental controls in Inner Mongolia. Acta Scientiarum Naturalium Universitatis Pekinensis, 2006, 42(6): 774-778.
马文红, 方精云. 内蒙古温带草原的根冠比及其影响因素. 北京大学学报(自然科学版), 2006, 42(6): 774-778.
[29] Dai C, Kang M Y, Ji W Y,etal. Responses of belowground biomass and biomass allocation to environmental factors in central grassland of Inner Mongolia. Acta Agrestia Sinica, 2012, 20(2): 268-274.
戴诚, 康慕谊, 纪文瑶, 等. 内蒙古中部草原地下生物量与生物量分配对环境因子的响应关系. 草地学报, 2012, 20(2): 268-274.
[30] Yang X J, Huang M, Wang J B,etal. Belowground biomass in Tibetan grasslands and its environmental control factors. Acta Ecologica Sinica, 2013, 33(7): 2032-2042.
杨秀静, 黄玫, 王军邦, 等. 青藏高原草地地下生物量与环境因子的关系. 生态学报, 2013, 33(7): 2032-2042.
