黄土高原草地和农田系统碳动态对降雨、温度和CO2浓度变化响应的模拟
2018-03-04郭丁郭文斐赵建特木其勒图李旭东傅华骆亦其草地农业生态系统国家重点实验室农业部草牧业创新重点实验室兰州大学草地农业科技学院甘肃兰州7000内蒙古阿拉善盟额济纳旗草原工作站内蒙古巴彦浩特75006内蒙古阿拉善盟草原监督管理所内蒙古巴彦浩特75006美国北亚利桑那大学工程林业和自然科学学院亚利桑那弗拉格斯塔夫860
郭丁,郭文斐,赵建,特木其勒图,李旭东,傅华*,骆亦其(.草地农业生态系统国家重点实验室,农业部草牧业创新重点实验室,兰州大学草地农业科技学院,甘肃 兰州 7000;.内蒙古阿拉善盟额济纳旗草原工作站,内蒙古 巴彦浩特 75006;.内蒙古阿拉善盟草原监督管理所,内蒙古 巴彦浩特 75006; .美国北亚利桑那大学工程林业和自然科学学院,亚利桑那 弗拉格斯塔夫 860)
全球气候变化已经成为各国政府和科学界关心的重大问题,其通常是多种环境条件(比如降水、温度和大气CO2浓度)同时连续地发生变化[1]。由于土地利用变化和化石燃料的使用,从工业革命后CO2浓度逐步上升,预计到21世纪末CO2浓度将增加到700×10-6[2]。CO2与其他温室气体浓度的升高,使得全球气温在过去150年增加了0.76 ℃,到21世纪末全球气温将增加1.5~6.4 ℃[2]。全球气温改变同时,全球降水格局也发生变化,北半球热带、亚热带降水量减少了3%, 而中高纬度降水量增加了5%~10%,同时极端气候事件发生频率增加[2]。
气候变化对生态系统的结构和功能产生了重要的影响,也深刻影响生态系统碳循环[1,3-6]。随着CO2浓度升高植物光合速率增加,在700×10-6时达到顶峰[7],但同时CO2浓度增加能导致土壤呼吸增加;增温能增加美国高草大草原植物在春天时的光合效率,而在早秋时光合效率降低,同时增温能显著增加植物的暗呼吸[8];降水的增加能促进土壤微生物活性[9],也有研究表明水分增加会导致土壤呼吸对温度敏感性的降低[10]。
全球气候变化多因子对生态系统的影响通常被假设为单因子作用的叠加[1],但已有研究表明其交互作用对生态系统过程的影响极其复杂。对内蒙古短花针茅(Stipabreviflora)草地的控制实验表明,增温和增雨协同作用能显著增加草地生物量[11]。Shaw等[12]对加利福尼亚州的一年生草地研究中发现CO2浓度的升高能够抑制由于温度升高和降水增加而引起的净初级生产力提高。内蒙古克氏针茅(Stipakrylovii)草原研究表明,增温增雨的交互作用抑制了土壤呼吸[13]。Zhou等[14]通过整合分析发现增温和提高CO2浓度交互作用显著增加了土壤呼吸,而降低了异养呼吸(Rh)。但多因子的控制实验并不能完全模拟环境的变化且非常昂贵,利用模型对气候变化进行模拟,分析其对生态系统的影响,是目前广泛采用的方法之一,其在加深理解全球变化对生态系统结构和功能的影响中发挥重要的作用[1,15-16]。应用LPJ-GUESS模型对鸡公山马尾松林研究表明,增温、增雨以及增加CO2能显著提高生产力和土壤呼吸[17]。利用CENTURY模型,模拟呼伦贝尔草甸草原地上净初级生产力的研究表明,增温、增雨和CO2浓度增加能显著提高地上净初级生产力[6]。Luo等[1]采用陆地生态系统模型(Terrestrial Ecosystem model, TECO model)研究温度、降水和CO2浓度变化对不同生态系统影响中发现,三因子交互作用对碳过程无显著作用,而二因子交互作用对净初级生产力(NPP),异养呼吸(Rh)和净生态系统生产力(NEE)有正反馈作用,并且每个生态系统的响应不一致。本研究应用Luo等[15]和Weng等[16]的TECO模型,并根据当地农田管理方式,对模型进行修正,增加有机肥添加和作物收获子模型,模拟黄土高原草地和农田生态系统在CO2浓度、温度和降水改变情况下生态系统碳动态。初步探讨黄土高原草地和农田生态系统碳过程对未来气候变化响应。
1 材料与方法
1.1 研究样地与数据来源
实验样地位于兰州大学干旱半干旱国际气候与环境监测站(SACOL),地理坐标为:东经104.137°,北纬35.946°,海拔为1965.8 m。该区属于大陆性半干旱气候,过去50年的年平均气温为6.7 ℃;年降水量为382 mm,主要集中在7月到9月;年蒸发量为1343 mm;日照时数约为2600 h;无霜期为90~140 d。土壤类型为灰钙土。
研究包含两种土地利用方式:1)农田(MC),在传统耕作方式下已经至少利用50年;2)围封草地(EG),部分农田从1986年后被弃耕自然恢复成草地。于2005年,每个土地利用方式随机设置3个50 m×50 m样地。植被类型是半干旱典型草原,草地主要植物有本氏针茅(Stipabungeana)、冷蒿(Artemisiafrigida)、铁杆蒿(Tripoliumvulgare)、赖草(Leymussecalinus)等。农业生产为雨养农业,主要的作物有马铃薯(Solanumtuberosum),谷子(Setariaitalica)和胡麻(Linumusitatissimum)等,作物一年一熟。农田于5月中旬施入尿素30 kg·hm-2,过磷酸钙60 kg·hm-2和有机肥3000 kg·hm-2(64 g C·m-2)。10月下旬采用连根拔起的方式收获作物,仅在土壤中残留少量的植物残体。
本研究中所用的气象资料(2007-2010年)由兰州大学干旱半干旱国际气候与环境监测站提供,气象资料包括每小时的气温、土温、饱和水势压差、相对湿度、降水、太阳辐射和CO2浓度。模型校正的数据为土壤呼吸[18]、地上生物量和土壤有机碳。
1.2 模型和参数描述
陆地生态系统(TECO)模型是从陆地碳固存模型[15]演变而来,其主要是用来检测生态系统对全球变化的响应。TECO模型包括冠丛模型、土壤水分动态模型、植物生长模型和土壤碳动态模型4个主要的组成部分(图1)。冠丛光合作用以及土壤水分动态模型模拟的时间步长为1 h,植物生长和土壤碳动态模型模拟的时间步长为1 d。模型具体的描述在Weng等[16]中,本研究在这里仅对模型进行简单的概括。
灌丛光合作用子模型是基于多层过程模型,其改进自Wang等[19]所写模型,子模型根据比尔定律通过计算冠丛辐射传输来模拟气孔导度、光合作用和能量分配。叶子分为被日光照射和庇荫两部分。光合作用根据Farquhar模型[20]和气孔导度模型[21]计算。土壤水分动态子模型将土壤分成10层,第一层为10 cm,其他9层为20 cm。各层土壤水分含量取决于水分的输入和输出。水分的输入包括降水输入到土壤表层以及由表层渗透到深层的水分。水分的输出包括蒸发、蒸腾和径流。蒸发决定于表层土壤含水量和大气对蒸发的需求。蒸腾由气孔导度、土壤水分和根系分布决定。植物生长子模型根据ALPHAPHA模型[22]和CTEM模型[23]模拟碳分配和物候。碳同化在根茎叶之间分配依赖于他们的生长速率和物候的变化。物候变化通过叶面积指数(LAI)变化体现。生长季节的开始由生长度日(GDD)决定。叶衰老由最低温度和最低土壤水分决定。当LAI低于一定的水平(LAI<0.1),生长季结束。碳转移子模型考虑碳通过凋落物从植物转移到土壤和凋落物及土壤有机碳的分解[15]。
根系深度和垂直分布决定植物对水分吸收潜力。草地中土壤根系主要分布在70 cm 土层以上,根系垂直分布呈动态,随根系的生长和死亡发生变化。根系在土壤中初始分配比率被设定为40% (0~10 cm)、40% (10~30 cm)、15% (30~50 cm)和5% (50~70 cm)[3]。
1.3 模型校正
由于研究区的农田每年5月耕种时每hm2土地施用3000 kg有机肥(64 g C·m-2),10月中下旬作物(谷子)收获仅留少许的植物残体留在土壤中,所以农田系统的模型在原模型的基础上,添加了作物移除和有机肥添加模块。模型运行时在5月中旬向凋落物碳库中添加64 g C,10月中下旬向凋落物库增加15 g C[24]以及将地上和地下植物活体碳库变为零。
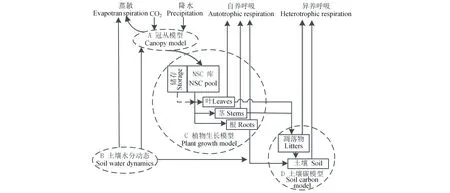
图1 陆地生态系统模型示意图[16]Fig.1 Schematic diagram of Terrestrial Ecosystem (TECO) model[16]方形表示各库;虚线的环表示4个子模型(A~D);NSC:非结构性碳。Boxes present pools; the dashed circles represent four submodels (A-D); NSC: Nonstructural carbon.
模型在气候改变条件下模拟运行之前,先将模型运行1000年使之达到稳定状态[3,16],然后将模拟值与观测值对比进行校正,调整模型的参数。
用土壤呼吸、土壤有机碳储量和地上生物量对模型进行校正。土壤呼吸采用LI-COR 6400配以9号呼吸室,草地土壤呼吸在每年的3-10月每月测定一次,农田土壤呼吸为每年的5-10月每月测定一次[18];草地地上生物量每年测定3次,为5,7和9月;农田地上生物量每年测定2次,为5和10月。
1.4 模拟情景
用模型模拟未来气候条件改变,包括改变温度,CO2浓度,降水以及三者之间的交互作用(表1)。所有气候改变均为每年持续地逐渐地发生100年,即从2011年到2110年结束。本研究所展示的结果为模拟第100年时NPP、Rh和NEE对气候变化响应的结果。
选取表2的情景计算气候变化因子之间的交互作用。

表1 本研究中模拟情景Table 1 Modeling scenarios in this study

表2 计算温度、降水和CO2浓度交互作用的情景Table 2 Modeling scenarios for studying interactions of temperature, precipitation and CO2 concentration in this study
0表示气候条件为当前状况;-和+分别表示温度减少或增加以及CO2浓度和降水减少或增加的百分比。Zero (0) represents the current condition; negative (-) or positive (+) values represent absolute decreases or increases in temperature and percentages of decreases or increases for CO2concentration and precipitation.
单因子和多因子对NPP,Rh和NEE主效应根据以下的公式计算[1]:
P100=P100T0C0-P0T0C0T2=P0T2C0-P0T0C0
C84=P0T0C84-P0T0C0
式中:P100T0C0表示降水增加100%时NPP、Rh和NEE的值;P0T0C0表示当前环境条件下NPP、Rh和NEE的值;P0T2C0表示温度增加2 ℃时NPP、Rh和NEE的值;P0T0C84表示CO2浓度增加84%时NPP、Rh和NEE的值;P100表示降水增加100%时主效应;T2表示温度增加2 ℃时主效应;C84表示CO2浓度增加84%时主效应。
双因子交互作用的主效应:
T2XP100=P100T2C0-P0T0C0-P100-T2
C84XP100=P100T0C84-P0T0C0-P100-C84
T2XC84=P0T2C84-P0T0C0-T2-C84
式中:P100T2C0表示降水增加 100%和温度升高2 ℃时NPP、Rh和NEE的值;P100T0C84表示降水增加 100%和CO2浓度增加84%时NPP、Rh和NEE的值;P0T2C84表示温度增加2 ℃和CO2浓度增加84%时NPP、Rh和NEE的值;T2XP100表示降水增加100%和温度升高2 ℃时交互作用;C84XP100表示降水增加 100%和CO2浓度增加84%时交互作用;T2XC84表示温度增加2 ℃和CO2浓度增加84%时交互作用;X指交互作用。
三因子交互作用:
T2XP100XC84=P100T2C84-P0T0C0-T2XP100-T2XP100-C84XP100-T2XC84-P100-T2-C84
式中:P100T2C84表示降水增加 100%、温度升高2 ℃和CO2浓度增加84%时NPP、Rh和NEE的值;T2XP100XC84表示降水增加 100%、温度升高2 ℃和CO2浓度增加84%交互作用。
由于计算不同样地交互作用,所以采用Luo等[1]方法计算相对幅度。如增温和增雨的交互作用为:
增温,增雨和增加CO2浓度三者交互作用为:
2 结果与分析
2.1 观测数据和模型对比
当模型达到平衡状态时,草地和农田生态系统碳库分别为4972.0和7110.4 g C·m-2,实测碳库分别为4935.5和7195.0 g C·m-2,具有较好的一致性。草地和农田生态系统的土壤呼吸和地上生物量模拟值与观测值都有较好的相关性(P<0.01);土壤呼吸的模拟值与观测值的线性关系分别为y=0.75x+0.28,R2=0.80(图2a和b)和y=0.72x+0.28,R2=0.62(图3a和b);地上生物量的线性关系分别为y=1.04x+6.69,R2=0.75(图2c和d)和y=0.90x+36.03,R2=0.97(图3c和d)。修正后的农田模型能很好地模拟农田系统碳动态。
2.2 单因子变化对草地和农田生态系统的影响
如图4(a, b, c)所示,随着温度上升,草地和农田生态系统NPP,Rh都表现为先增加后减少的趋势, NEE随着温度的升高呈现先减少后增加的趋势,除了农田生态系统的Rh在0 ℃时达最大值,其余的均在+4 ℃时达到最大值。农田生态系统的NPP和NEE对温度的响应高于草地生态系统,而Rh对温度的响应在草地生态系统较高。以+2 ℃为例,相比于0 ℃,草地生态系统NPP和Rh分别增加了28.7和28.0 g C·m-2,增加率为12.1%和11.9%;而农田生态系统NPP增加了73.8 g C·m-2,Rh降低了4.3 g C·m-2,增加率为19.7%和-1.9%;草地和农田生态系统分别增加了0.7和81.1 g C·m-2(图5)。
草地和农田生态系统的NPP、Rh都随着降水量(图4d、e和f)以及CO2浓度(图4g、h和i)的增加而增加,而NEE随其增加而减少。在降水量减少的情景下,农田生态系统NPP,NEE对降水量改变的响应比草地生态系统高,而在增加降水量的情景下趋势相反,例如当降水量增加100%时,草地生态系统NPP和Rh分别提高了14.9%和13.8%(图5);农田生态系统NPP和Rh分别提高了9.4%和4.0%(图5)。CO2浓度升高对草地和农田生态系统的影响低于温度和降水的改变。当CO2浓度从当前380×10-6升高到700×10-6,草地生态系统NPP和Rh升高了7.8%和6.7%(图5);农田生态系统NPP和Rh升高了4.1%和2.0%(图5)。
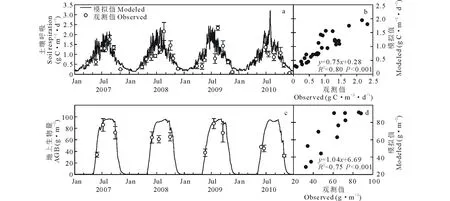
图2 草地生态系统土壤呼吸(a和b)和地上生物量(c和d)与模拟值对比Fig.2 Observed and simulated soil respiration (a and b) and aboveground biomass (c and d) in grassland system误差棒表示标准差,AGB表示地上生物量。Error bars denote±SD, AGB is aboveground biomass. 下同The same below.
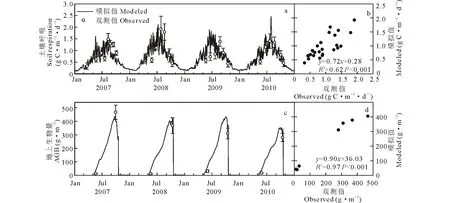
图3 农田生态系统土壤呼吸(a和b)和地上生物量(c和d)与模拟值对比Fig.3 Observed and simulated soil respiration (a and b) and aboveground biomass (c and d) in cropland system
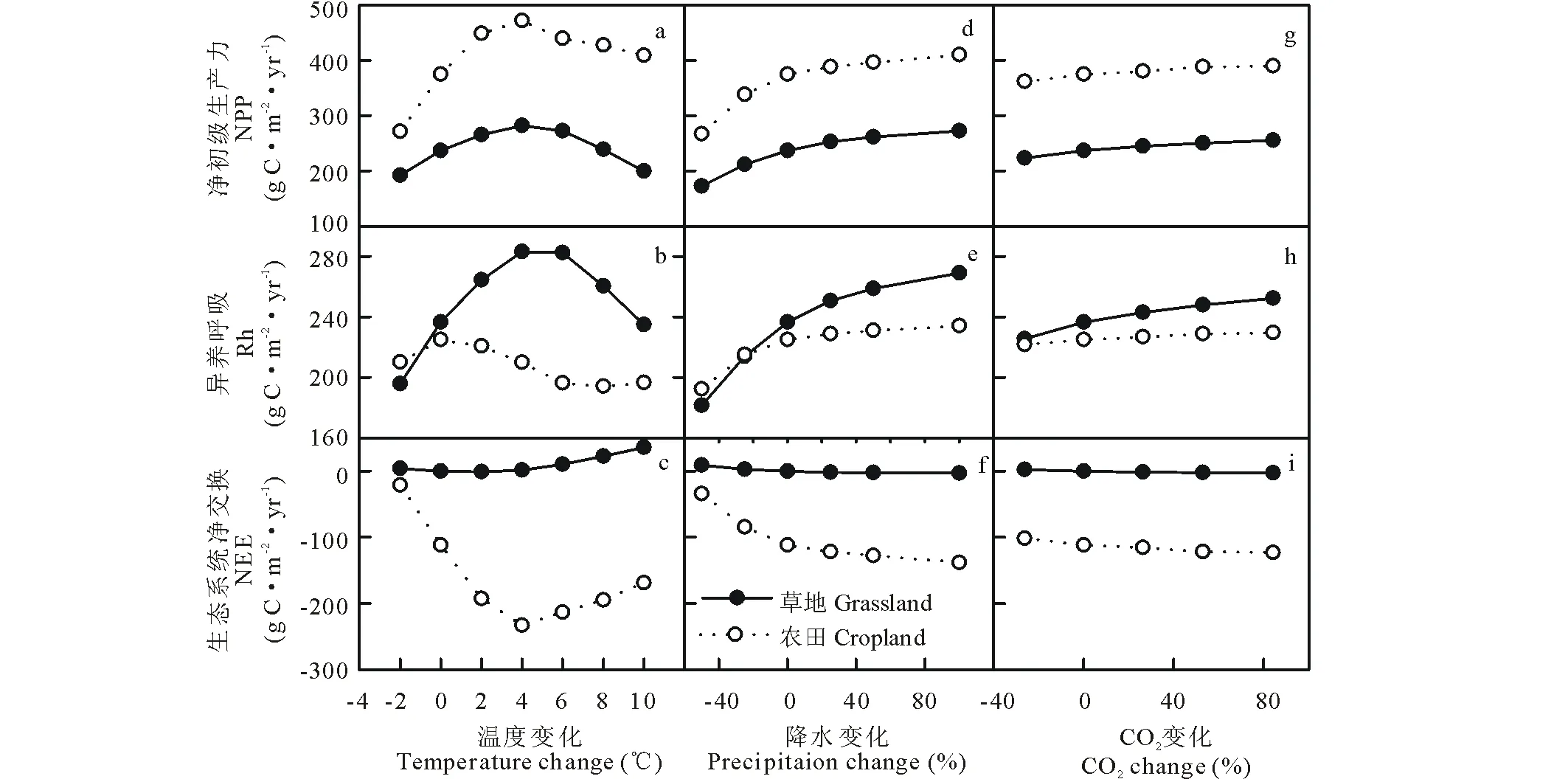
图4 单因子(温度,降水和CO2)改变对草地和农田生态系统NPP(净初级生产力),Rh (异养呼吸)和NEE (净生态系统交换)的影响Fig.4 Responses of NPP (net primary production), Rh (heterotrophic respiration) and NEE (net ecosystem exchange) to single-factor changes in temperature, precipitation and CO2
2.3 多因子变化对草地和农田生态系统NPP、Rh和NEE的影响

图5 相对于自然状态下,降水量减少50% (P-50),增加100% (P100),温度升高2 ℃ (T2)以及CO2浓度增加84% (C84)和各因子交互作用对NPP,Rh和NEE的影响Fig.5 Changes relative to control in NPP, Rh and NEE in response to treatments of halved precipitation (P-50), doubled precipitation (P100), elevated temperature (T2), elevated CO2 (C84) and their combination
降水和CO2浓度的改变影响生态系统NPP,Rh和NEE对温度变化的响应(图6和图7)。随着降水的增加,草地生态系统NPP和Rh不仅随之升高,NEE减少,而且其对温度响应的拐点发生改变,从降水减少50%温度0 ℃逐步变化到降水增加100%温度+8 ℃(图6a和b);而生态系统的NEE拐点变化较小,仅从0 ℃变化到温度+2 ℃(图6c)。农田生态系统NPP,Rh和NEE随着降水增加而增加,但是其随着温度变化的拐点并未发生改变(图6d,e和f)。同时,随着降水量从减少50%到增加100%,草地和农田生态系统NPP,Rh和NEE对温度响应更敏感。
相同温度下,CO2浓度升高增加了草地和农田生态系统NPP,Rh和NEE(图7)。在农田生态系统中,温度变化大于+4 ℃,CO2浓度升高使NPP和NEE变化趋势变缓,即对温度的响应不敏感(图7d和f)。在相同降水条件下,随着CO2浓度的升高,草地和农田生态系统NPP,Rh和NEE都有轻微提高,但是CO2浓度的改变并未改变草地和农田生态系统NPP,Rh和NEE对降水的响应趋势(图8)。
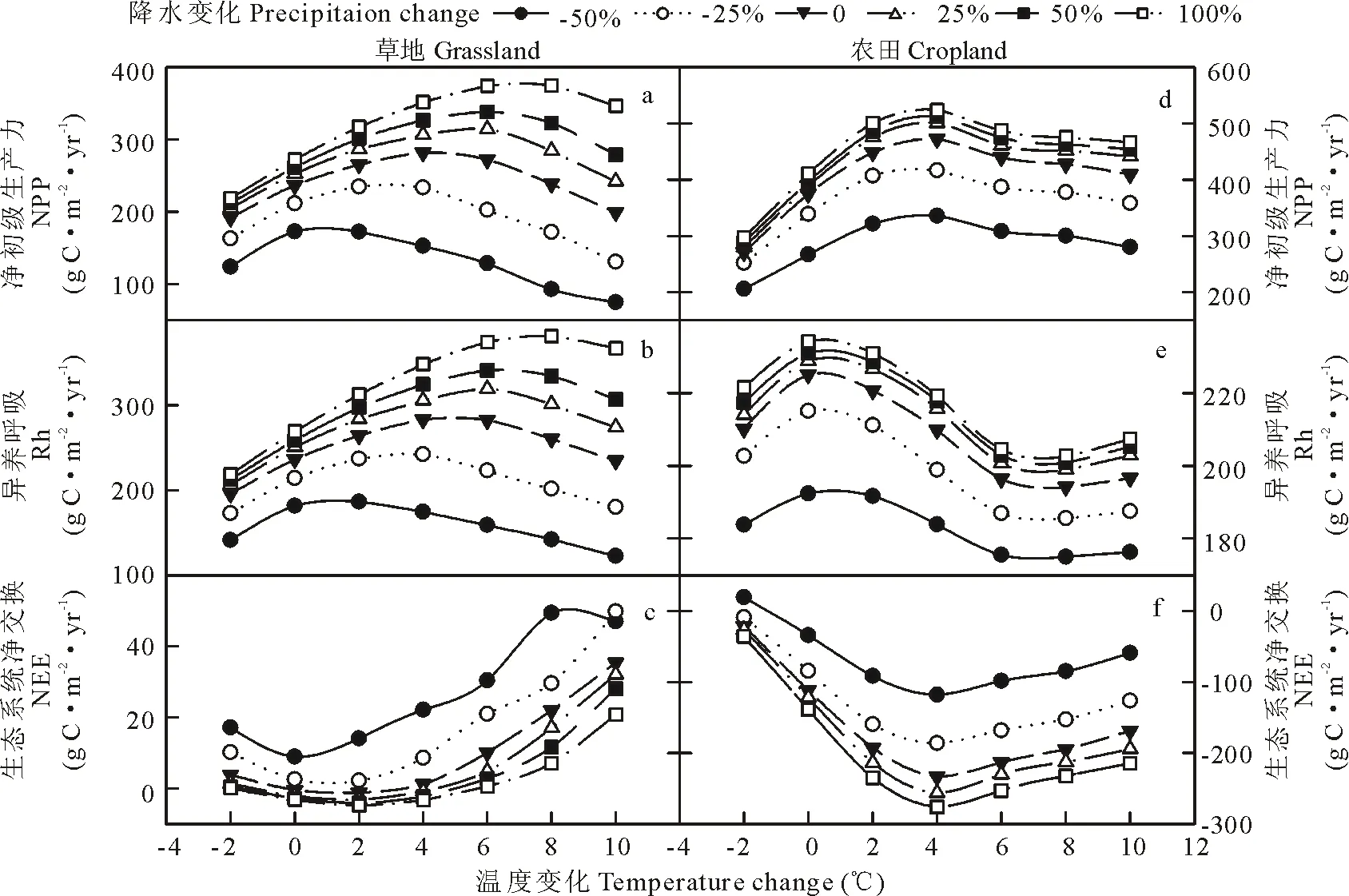
图6 草地和农田系统的NPP,Rh和NEE对温度和降水改变的响应Fig.6 Responses of NPP, Rh and NEE to changes in temperature and precipitation in grassland and cropland systems
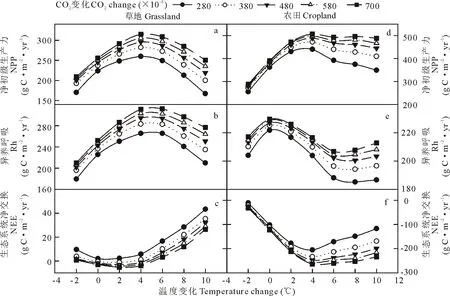
图7 草地和农田系统的NPP,Rh和NEE对温度和CO2浓度改变的响应Fig.7 Responses of NPP, Rh and NEE to changes in temperature and CO2 concentration in grassland and cropland systems
温度,降水和CO2浓度同时变化,改变生态系统NPP,Rh和NEE对气候变化的反应方式(图9和图10)。在CO2浓度280×10-6情境下,随着降水量改变,草地和农田生态系统的NPP对温度响应的拐点从0 ℃升高到+6 ℃(图9a)和+2 ℃到+4 ℃(图10a);在CO2浓度700×10-6情景下,其对温度响应拐点分别从0 ℃变化到+8 ℃(图9g)和没有变化(图10g)。草地生态系统Rh对温度,降水和CO2反应方式和NPP一致,农田生态系统Rh拐点没有变化。草地生态系统NEE对温度响应拐点在280×10-6下从0 ℃变化到+2 ℃(图9c),在700×10-6下从0 ℃变化到+4 ℃(图9i);农田生态系统NEE在不同CO2浓度下对温度响应拐点未发生改变。
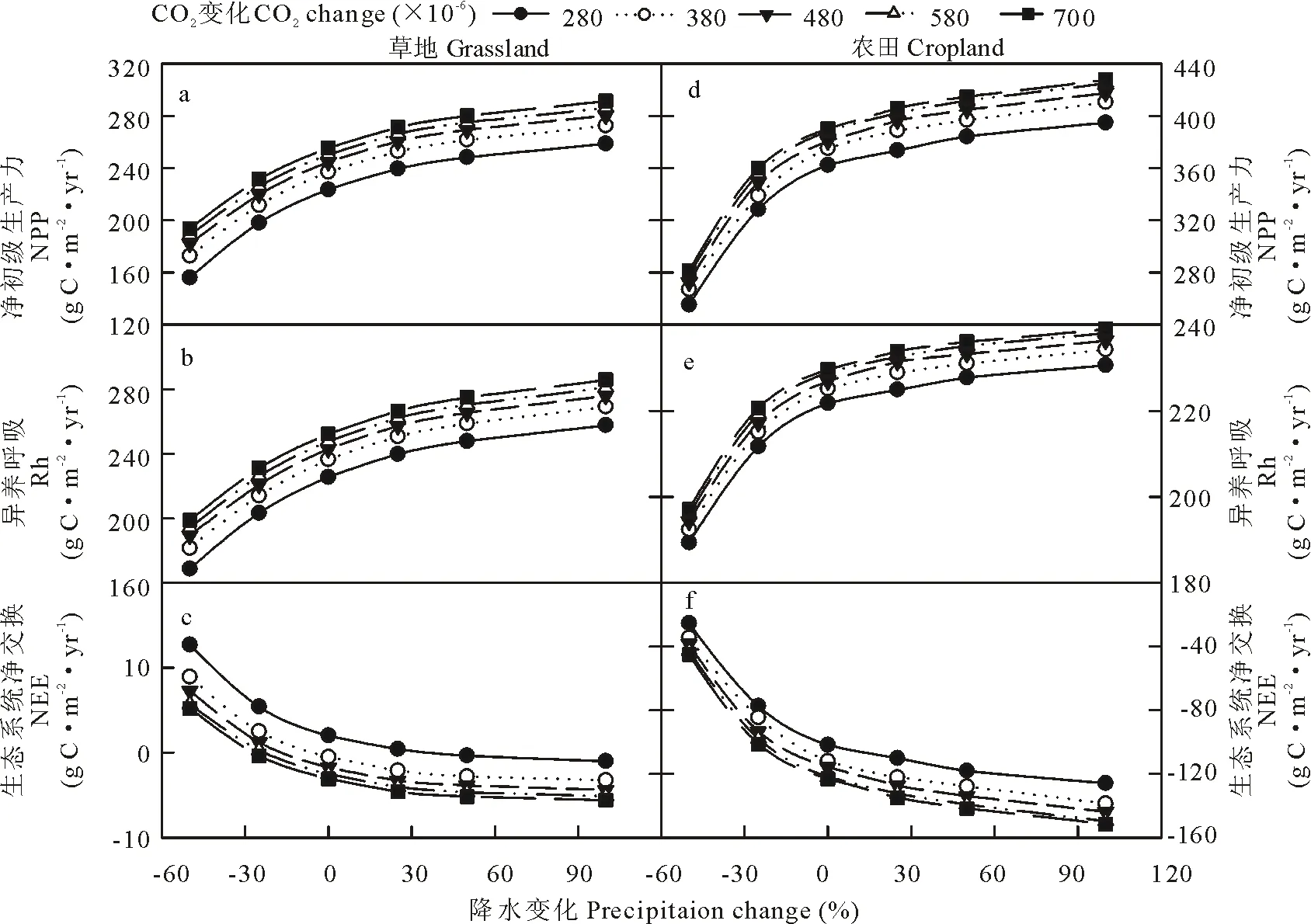
图8 草地和农田系统的NPP,Rh和NEE对降水和CO2浓度改变的响应Fig.8 Responses of NPP, Rh and NEE to changes in precipitation and CO2 concentration in grassland and cropland systems
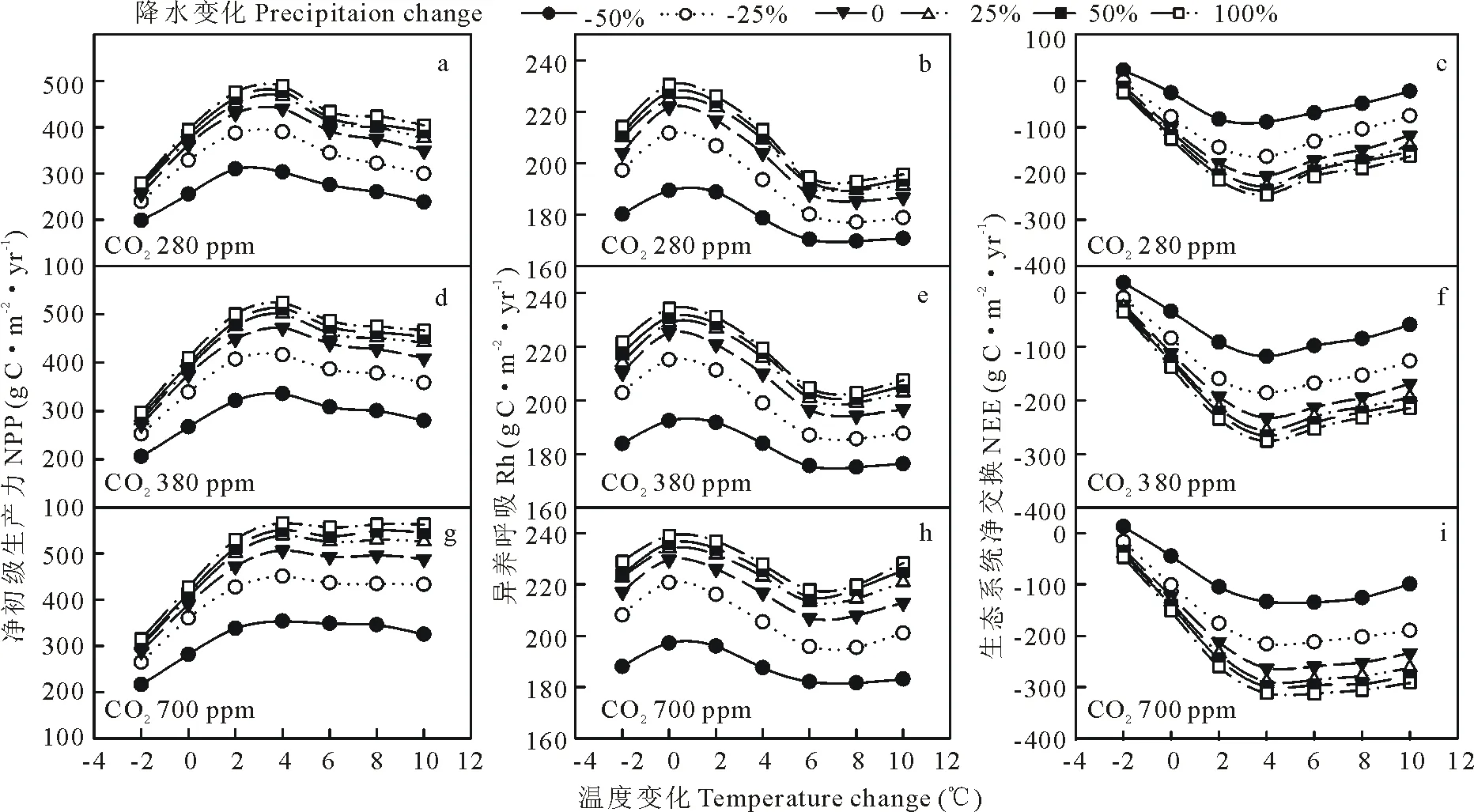
图10 农田系统的NPP,Rh和NEE对温度、降水和CO2浓度改变的响应Fig.10 Responses of NPP, Rh and NEE to changes in temperature, precipitation and CO2 concentration in cropland system
增温,增加降水和增加CO2浓度都可以增加生态系统NPP和Rh,降低生态系统NEE,降水减少表现相反的趋势(图5)。增温、增加CO2浓度和增加降水两因子或三因子同时改变可以增加生态系统NPP和Rh,降低系统NEE。而降水降低和增温或增加CO2浓度结合时,生态系统NPP和Rh降低,NEE升高(图5)。
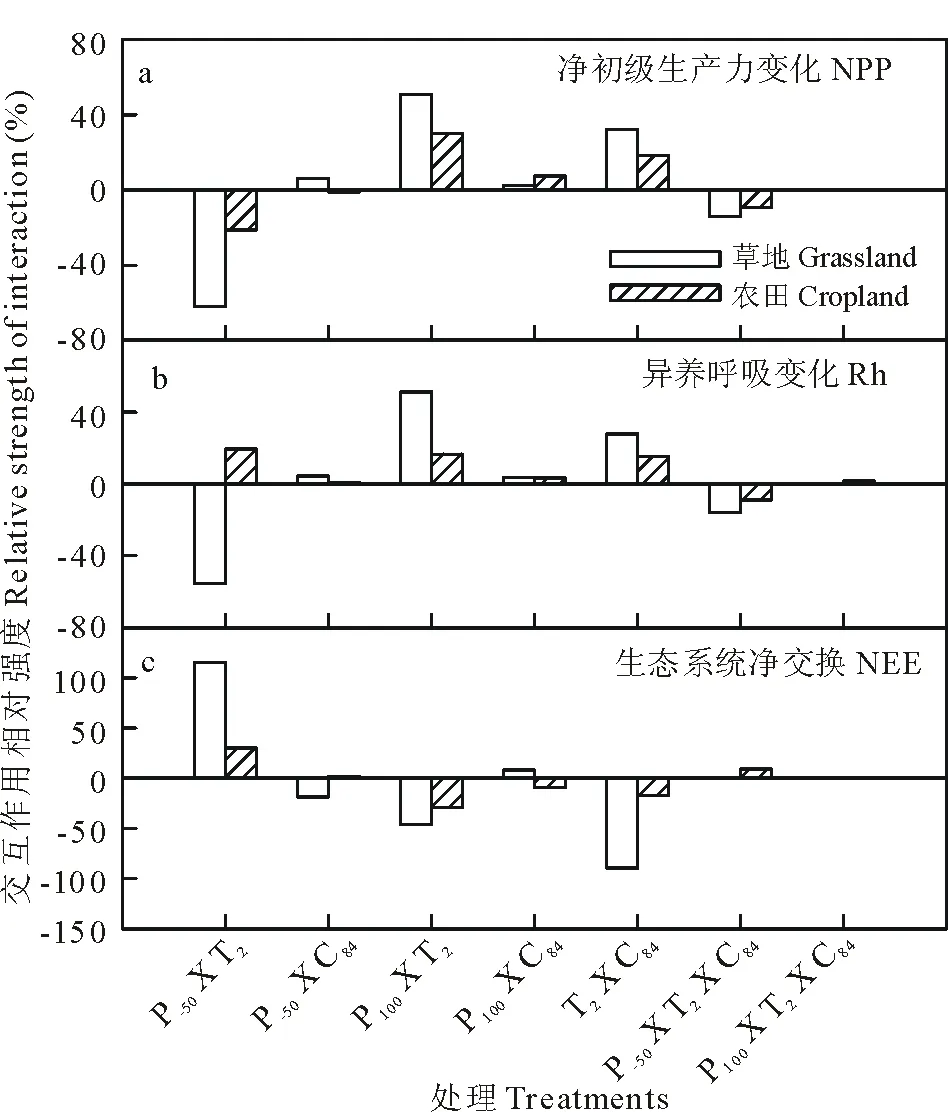
图11 双因子和三因子交互作用对NPP,Rh和NEE影响的相对强弱Fig.11 Relative strength of two- or three-ways interactive effects on NPP, Rh and NEE详细缩写解释见图5。Detailed information about abbreviations were shown in Fig.5.
如图11所示,总体而言,增温和增雨,增雨和增加CO2浓度,增温和增加CO2浓度的交互作用对草地和农田生态系统的NPP,Rh有着正反馈作用,而NEE负反馈作用;除农田生态系统Rh外,增温和减少降水量的交互作用对草地和农田生态系统的NPP,Rh有着负反馈作用,而NEE正反馈作用。减雨,增温和增加CO2浓度和增雨,增温和增加CO2浓度三者之间交互作用对生态系统作用并不明显(图11)。
3 讨论
3.1 单因子对生态系统NPP,Rh和NEE的影响
在较低的温度情况下,增温能显著地刺激植物生长和土壤呼吸,由于延长了其生长季节和增强了土壤N的矿化[25],表现为碳的固定(图4c)。亓伟伟等[26]利用BIOME-BGC模型模拟增温对生态系统碳通量的结果表明,青藏高原增温1.2~1.7 ℃能增加生态系统生产力和呼吸,使生态系统碳微弱增加。Xu等[4]野外增温实验研究也表明当温度增加3 ℃时,生态系统NPP提高。Luo等[27]研究表明,北美高草草原在温度增高2 ℃下,生态系统NPP增加14%,土壤呼吸增加8%。在温度过高情况下,增温减少了土壤水分,导致土壤水分的限制[28],温度的增高依赖与水分之间的交互作用。在高温时,Rh对温度的响应更明显,所以在温度升高4 ℃后,生态系统对碳固定逐渐地减少(图4c)。Zhou等[3]采用此模型模拟美国高草草原对气候变化响应结果表明,NPP,Rh和NEE对温度响应的拐点分别为5,6和7 ℃,本研究对于温度升高的耐性低于此,其主要原因可能是由于该研究地点的降水量高于黄土高原,提高了生态系统对温度升高的耐性。未来气温升高2 ℃,以C4植物为主的农田生态系统NPP增加率高于草地生态系统(图5),主要是由于增温更有利于C4植物的生长[29]。
通过增加降水或减少降水的实验表明,地上的NPP和植物密度随着降水呈现线性增加的趋势[30]。在内蒙古典型草原、荒漠草原[31]和内蒙古半干旱草原[32]的研究都表明NPP随着降水量增加而增加。对果林、裸地和农用地等不同土地利用方式土壤呼吸研究表明,降水能够使土壤呼吸产生激增效应[33]。Wu等[34]通过对控制实验的整合分析表明,降水的增加能显著地增加植物生物量,刺激土壤呼吸,最终增加生态系统碳固存。本研究也发现随着降水量的增加,生态系统NPP和Rh都显著增加,最终增加了生态系统碳固存(图4f)。相比起呼吸,生态系统的生产力对水分变化更加敏感。草地生态系统NPP和Rh对水分变化比农田生态系统更加敏感,其原因可能是由于农田土壤含水量高于草地所引起的。
生态系统对CO2浓度变化的响应与降水类似。CO2浓度升高能够增加植物光合作用、生长和地下碳输入,最终导致微生物活性的提高和呼吸的增加[35]。另外,高浓度CO2能减少植物气孔导度和蒸腾作用,从而提高土壤水分含量,加强土壤微生物活性和呼吸[36-37]。但长时间高浓度CO2导致植物对CO2敏感性降低[38]。同时,CO2浓度增加使植物对N的需求增加,最终将改变凋落物的C∶N和分解速率,从而影响生态系统的NPP,Rh和NEE[39]。由于光合适应性和不同物种的差异,植物光合作用对CO2浓度变化的响应是多样的[40]。本研究模型并没有反映生态系统对气候变化影响的同化过程,导致CO2对生态系统的影响为渐近线方式。C3植物光合速率对CO2浓度升高的响应高于C4植物[41],使得以C3植物为主要物种的草地生态系统NPP对CO2浓度升高的响应高于农田生态系统(图5)。
3.2 多因子交互作用对NPP,Rh和NEE的影响
草地和农田生态系统碳动态对温度、CO2浓度和降水改变的响应与其对单因子的响应具有不同的趋势(图5、6和7)。降水的增加或CO2浓度增加能抵消高温所引起的副效应,导致生态系统碳动态对温度响应的更高拐点[3];降水的减少加强了生态系统水分的压力[42],使生态系统NPP和Rh对温度响应不敏感,即随温度升高其变化曲线更加平缓(图5)。Zhou等[3]利用TECO模型对美国高草草原的研究中也发现了相同的趋势,而Cowling等[43]运用DGVM模型对亚马逊热带雨林的研究表明生态系统碳动态对温度响应的阈值随着降水的增加而减小,其可能是由于不同的生态系统类型所导致(热带雨林vs.草地)[3]。
草地和农田生态系统T2XP100,C84XP100和T2XC84的交互作用对NPP和Rh的影响较为一致,都能够增加NPP、Rh和碳的固定(图11)。在水分是主要限制因子的黄土高原区,增温和CO2浓度增加对生态系统的正交互作用,其原因可能是增加CO2浓度能减少气孔导度,提高水分利用效率[28],增加土壤水分的可利用性[36-37]。CO2浓度增加能够缓解由于增温而导致的土壤干旱及减轻水分压力[44]。同样,降水增加和温度升高对生态系统正的交互作用,主要是由于降水量的增加能够缓解由于温度升高而造成的水分损失,提高土壤水分的可利用性[45]。当降水量减少一半时增温能显著地降低生态系统NPP,导致生态系统碳释放增加。草地生态系统降水量减少和CO2浓度增加的交互作用对生态系统碳固定有正反馈作用;而在农田生态系统其对碳固定没有正反馈作用,但比起单独降低降水量或CO2浓度升高而言,其能够缓解单独减少降水量所造成生态系统碳损失。如上所述,CO2浓度升高能够缓解土壤干旱和降低水分压力。同时,本研究发现草地生态系统碳过程对多因子的交互作用响应更明显,其主要由于草地土壤含水量低于农田。Luo等[1]的研究也表明干旱地区碳过程对降水变化更加敏感。
4 结论
在黄土高原区,随着温度升高,草地和农田生态系统NPP和Rh都呈现先增加后减少的趋势,NEE表现出先减少后增加趋势。草地和农田生态系统NPP和Rh都随着降水增加和CO2浓度升高而增加,NEE随着降水增加和CO2浓度升高而减少,系统固碳能力增加。降水和CO2浓度增加将改变生态系统NPP,Rh和NEE对温度变化的响应。随着降水增加,生态系统对温度响应更敏感;草地生态系统NPP,Rh和NEE对温度变化响应拐点发生改变,而农田系统无变化。CO2浓度升高使农田生态系统NPP,Rh和NEE在>+4 ℃情景下,对温度变化响应不敏感。增温与增加CO2浓度和增加降水的2因子交互作用对生态系统NPP和Rh都有正反馈作用,对NEE有负反馈作用。三因子交互作用对生态系统NPP,Rh和NEE没有明显促进作用。草地生态系统碳动态对气候变化交互作用响应更显著。
References:
[1] Luo Y Q, Gerten D, Le Maire G,etal. Modeled interactive effects of precipitation, temperature, and [CO2] on ecosystem carbon and water dynamics in different climatic zones. Global Change Biology, 2008, 14(9): 1986-1999.
[2] IPCC. Climate Change 2007: the Physical Basis//Solomon D. Cambridge, UK: Cambridge Universtiy Press, 2007.
[3] Zhou X H, Weng E S, Luo Y Q. Modeling patterns of nonlinearity in ecosystem responses to temperature, CO2, and precipitation changes. Ecological Applications, 2008, 18(2): 453-466.
[4] Xu X, Sherry R A, Niu S,etal. Net primary productivity and rain use efficiency as affected by warming, altered precipitation, and clipping in a mixed grass prairie. Global Change Biology, 2013, 19(9): 2753-2764.
[5] Yan Z Q, Qi Y C, Dong Y S,etal. Nitrogen cycling in grassland ecosystem in response to climate change and Human activities. Acta Prataculturae Sinica, 2014, 23(6): 279-292.
闫钟清, 齐玉春, 董云社, 等. 草地生态系统氮循环关键过程对全球变化及人类活动的响应与机制. 草业学报, 2014, 23(6): 279-292.
[6] Zhang C H, Wang M J, Zhang L,etal. Response of meadow steppe ANPP to climate change in Hulunbeir, Inner Mongolia-a simulation study. Acta Prataculturae Sinica, 2013, 22(3): 41-50.
张存厚, 王明玖, 张立, 等. 呼伦贝尔草甸草原地上净初级生产力对气候变化响应的模拟. 草业学报, 2013, 22(3): 41-50.
[7] Silvola J, Ahlholm U. Photosynthesis in willows (Salix×dasyclados) grown at different CO2concentrations and fertilization levels. Oecologia, 1992, 91(2): 208-213.
[8] Zhou X H, Liu X Z, Wallace L L,etal. Photosynthetic and respiratory acclimation to experimental warming for four species in a tallgrass prairie ecosystem. Journal of Integrative Plant Biology, 2007, 49(3): 270-281.
[9] Yan Z Q, Qi Y C, Li S J,etal. Soil microorganisms to enzyme activity of grassland ecosystem affected by changes in precipitation pattern and increase in nitrogen deposition-a review. Micobiology China, 2017, 46(6): 1481-1490.
闫钟清, 齐玉春, 李素俭, 等. 降水和氮沉降增加对草地土壤微生物与酶活性的影响研究进展. 微生物学通报, 2017, 46(6): 1481-1490.
[10] Chen Q S, Li L H, Han X G,etal. Temperature sensitivity of soil respiration in relation to soil moisture in 11 communities of typical temperate steppe in Inner Mongolia. Acta Ecologicca Sinica, 2004, 24(4): 831-836.
陈全胜, 李凌浩, 韩兴国, 等. 典型温带草原群落土壤呼吸温度敏感性与土壤水分的关系. 生态学报, 2004, 24(4): 831-836.
[11] Lü X M, Wang Y H, Zhou G S,etal. Interactive effects of changing precipitation and elevated temperatures on plant biomass and its allocation ofStipabreviflora. Acta Ecologica Sinica, 2015, 35(3): 752-760.
吕晓敏, 王玉辉, 周广胜, 等. 温度与降水协同作用对短花针茅生物量及其分配的影响. 生态学报, 2015, 35(3): 752-760.
[12] Shaw M R, Zavaleta E S, Chiariello N R,etal. Grassland responses to global environmental changes suppressed by elevated CO2. Science, 2002, 298(5600): 1987-1990.
[13] Gu R, Chao L M, Zhang L X,etal. The influence of hydorthermal factors on soil respiration and soil temperature sensitivity ofStipakryloviisteppe, Inner Mongolia, China. Acta Prataculturae Sinica, 2015, 24(4): 21-29.
谷蕊, 潮洛蒙, 张立欣, 等. 水热因子对克氏针茅草原土壤呼吸及其土壤温度敏感性的影响. 草业学报, 2015, 24(4): 21-29.
[14] Zhou L Y, Zhou X H, Shao J J,etal. Interactive effects of global change factors on soil respiration and its components: a meta-analysis. Global Change Biology, 2016, 22(9): 3157-3169.
[15] Luo Y Q, Reynolds J F. Validity of extrapolating field CO2experiments to predict carbon sequestration in natural ecosystems. Ecology, 1999, 80(5): 1568-1583.
[16] Weng E S, Luo Y Q. Soil hydrological properties regulate grassland ecosystem responses to multifactor global change: A modeling analysis. Journal of Geophysical Research-Biogeosciences, 2008, 113(G3): G03003.
[17] Feng X H, Cheng R M, Xiao W F,etal. Productivity and carbon dynamic of the Masson Pine stands in Jigongshan region based on LPJ-GUESS model. Scientia Silvae Sinicae, 2013, 49(4): 7-15.
封晓辉, 程瑞梅, 肖文发, 等. 基于LPJ-GUESS模型的鸡公山马尾松林生产力和碳动态. 林业科学, 2013, 49(4): 7-15.
[18] Li X D, Fu H, Guo D,etal. Partitioning soil respiration and assessing the carbon balance in aSetariaitalica(L.) Beauv. cropland on the Loess Plateau, Northern China. Soil Biology & Biochemistry, 2010, 42(2): 337-346.
[19] Wang Y P, Leuning R. A two-leaf model for canopy conductance, photosynthesis and partitioning of available energy I: Model description and comparison with a multi-layered model. Agricultural and Forest Meteorology, 1998, 91(1): 89-111.
[20] Farquhar G, von Caemmerer S V, Berry J. A biochemical model of photosynthetic CO2assimilation in leaves of C3species. Planta, 1980, 149(1): 78-90.
[21] Ball J T, Woodrow I E, Berry J A. A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions//Biggins J, Editor. Progress in Photosynthesis Research. Martinus Nijhoff: Zoetermeer, Netherlands, 1987: 221-224.
[22] Luo Y, Meyerhoff P, Loomis R. Seasonal patterns and vertical distributions of fine roots of alfalfa (MedicagosativaL.). Field Crops Research, 1995, 40(2): 119-127.
[23] Arora V K, Boer G J. A parameterization of leaf phenology for the terrestrial ecosystem component of climate models. Global Change Biology, 2005, 11(1): 39-59.
[24] Li X, Fu H, Li X,etal. Effects of land-use regimes on carbon sequestration in the Loess Plateau, northern China. New Zealand Journal of Agricultural Research, 2008, 51(1): 45-52.
[25] Peng F. Initial Response of Alpine Meadow Ecosystem to Warming Manipulation in Permafrsot Area of Qinghai-Tibet Plateau. Beijing: University of Chinese Academy of Sciences, 2012.
彭飞. 模拟增温对多年冻土高寒草甸生态系统的影响. 北京: 中国科学院大学, 2012.
[26] Qi W W, Niu H S, Wang S P,etal. Simulation of effects of warming on carbon budget in alpine meadow ecosystem on the Tibetan Plateau. Acta Ecologica Sinica, 2012, 32(6): 1713-1722.
亓伟伟, 牛海山, 汪诗平, 等. 增温对青藏高原高寒草甸生态系统固碳通量影响的模拟研究. 生态学报, 2012, 32(6): 1713-1722.
[27] Luo Y Q, Sherry R, Zhou X H,etal. Terrestrial carbon-cycle feedback to climate warming: experimental evidence on plant regulation and impacts of biofuel feedstock harvest. Global Change Biology Bioenergy, 2009, 1(1): 62-74.
[28] Drake B G, Gonzàlez-Meler M A, Long S P. More efficient plants: a consequence of rising atmospheric CO2. Annual Review of Plant Biology, 1997, 48(1): 609-639.
[29] Chen Y Q. Effects of Simulated Warming on Growth and Water Utilization of C3and C4Plants. Hohehot: Inner Mongolia University, 2014.
陈宇琪. 模拟增温对C3、C4植物生长及水分利用的影响. 呼和浩特: 内蒙古大学, 2014.
[30] Yahdjian L, Sala O E. Vegetation structure constrains primary production response to water availability in the Patagonian steppe. Ecology, 2006, 87(4): 952-962.
[31] Mu S J, Li J L, Yang H F,etal. Spatio-temporal variation analysis of grassland net primary productivity and its relationship with climate over the past 10 years in Inner Mongolia. Acta Prataculturae Sinica, 2013, 22(3): 6-15.
穆少杰, 李建龙, 杨红飞, 等. 内蒙古草地生态系统近10年NPP时空变化及其与气候的关系. 草业学报, 2013, 22(3): 6-15.
[32] Ma W H, Yang Y H, He J S. Above- and belowground biomass in relation to environmental factors in temperate grasslands, Inner Mongolia. Science in China Series C: Life Sciences, 2008, 38(1): 84-92.
马文红, 杨元合, 贺金生. 内蒙古温带草地生物量及其与环境因子的关系. 中国科学(C 辑), 2008, 38(1): 84-92.
[33] Su H M, Li X Y, He B H,etal. Rainfall effects on soil microbial biomass and soil respiration under different land use conditions. Journal of Soil and Water Conservation, 2011, 25(6): 92-95.
苏慧敏, 李叙勇, 何丙辉, 等. 不同土地利用方式下降雨对土壤微生物量和呼吸的影响. 水土保持学报, 2011, 25(6): 92-95.
[34] Wu Z, Dijkstra P, Koch G W,etal. Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Global Change Biology, 2011, 17(2): 927-942.
[35] Anderson L J, Maherali H, Johnson H B,etal. Gas exchange and photosynthetic acclimation over subambient to elevated CO2in a C3-C4grassland. Global Change Biology, 2001, 7(6): 693-707.
[36] Morgan J, Pataki D, Körner C,etal. Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia, 2004, 140(1): 11-25.
[37] Hungate B A, Reichstein M, Dijkstra P,etal. Evapotranspiration and soil water content in a scrub-oak woodland under carbon dioxide enrichment. Global Change Biology, 2002, 8(3): 289-298.
[38] Jiang Y L, Zhou G S, Wang Y H,etal. Advances in the adaptability of zonalStipaplants to CO2and climate change in Inner Mongolia. Acta Ecologica Sinica, 2015, 35(14): 4559-4569.
蒋延玲, 周广胜, 王玉辉, 等. 内蒙古地带性针茅植物对CO2和气候变化的适应性研究进展. 生态学报, 2015, 35(14): 4559-4569.
[39] Johnson D. Simulated nitrogen cycling response to elevated CO2inPinustaedaand mixed deciduous forests. Tree Physiology, 1999, 19: 321-327.
[40] Luo Y Q, Sims D A, Griffin K L. Nonlinearity of photosynthetic responses to growth in rising atmospheric CO2: an experimental and modelling study. Global Change Biology, 1998, 4(2): 173-183.
[41] Wang X L, Xu S H, Liang H. The experimental study of the effects of CO2concentration enrichment on growth, development and yield of C3and C4crops. Scientia Agricultura Sinica, 1998, 31(1): 56-62.
王修兰, 徐师华, 梁红. CO2浓度增加对C3、C4作物生育和产量影响的实验研究. 中国农业科学, 1998, 31(1): 56-62.
[42] Ise T, Moorcroft P R. The global-scale temperature and moisture dependencies of soil organic carbon decomposition: an analysis using a mechanistic decomposition model. Biogeochemistry, 2006, 80(3): 217-231.
[43] Cowling S A, Shin Y. Simulated ecosystem threshold responses to co-varying temperature, precipitation and atmospheric CO2within a region of Amazonia. Global Ecology and Biogeography, 2006, 15(6): 553-566.
[44] Wall G, Garcia R, Kimball B,etal. Interactive effects of elevated carbon dioxide and drought on wheat. Agronomy Journal, 2006, 98(2): 354-381.
[45] Gerten D, Luo Y, Le Maire G,etal. Modelled effects of precipitation on ecosystem carbon and water dynamics in different climatic zones. Global Change Biology, 2008, 14(10): 2365-2379.
