微生境对华山松人工林天然更新的影响
2018-02-27张小鹏王得祥杜江涛张宋智
张小鹏,王得祥,杜江涛,张宋智
(1 西北农林科技大学 林学院,陕西 杨凌 712100;2 甘肃小陇山森林生态系统国家定位观测研究站,甘肃 天水 741022;3 甘肃省栎类次生林生态系统重点实验室,甘肃 天水 741022)
据全国第八次森林资源清查数据,我国人工林面积已达6 933万hm2[1]。人工林经营已成为生态恢复的重要手段[2-3],寻求适宜的管理措施,对促进人工林稳定持续地完成天然更新具有重要的现实意义。然而森林更新是一个极为复杂的生态学过程,不断受到树种类别[4]、自然环境[5-6]以及干扰竞争[7]等多种因素的影响。研究认为,在有充足种源的情况下,具有支持种子萌发、幼苗生长和幼树形成的环境条件,是维护森林能够顺利完成天然更新的基础,其中任何一个环节出现问题都会对更新进程产生影响[8]。但事实上,许多树种都存在更新不足的情况[9],林下“只见幼苗,不见幼树”的现象也颇为常见[10]。对其原因,相关研究多是从立地条件等方面加以解释,如Mühlenberg等[11]研究认为,阴坡环境更有利于针叶林的天然更新;Dobrowolska[12]发现,林木密度过大会抑制栎类树种幼苗幼树的生长。这些研究充分说明了森林更新问题的复杂性以及林下微生境可能发挥的作用,因而有必要进一步弄清天然更新与环境因子之间的关系,这对于阐明森林天然更新机理和指导人工林科学经营都具有极其重要的理论和现实意义。
华山松(Pinusarmandii)为松科(Pinaceae)松属(Pinus)常绿乔木,系我国的特有种,因其育苗简易、种子可食、木质优良等原因,已成为西部地区最重要的造林树种之一[13]。本研究选择位于秦岭西段的小陇山林区华山松人工林为研究对象,通过调查阴坡和阳坡环境不同间伐强度林下的微生境特性以及同一区域的幼苗幼树存活数量,旨在探讨影响华山松人工林天然更新的微生境因子及其作用,以及华山松幼苗向幼树的生长过渡有无障碍及其现象解释,以期为华山松的更新机制和人工林抚育措施优化提供一定的理论和实践依据。
1 试验区概况
试验区位于秦岭西段小陇山林区的党川林场(34°06′-34°26′N,106°04′-106°18′E),海拔为1 080.0~2 500.0 m,坡度为24°~38°。境内地势由西北向东南倾斜,山体相对高差100~550 m,年平均气温10.9 ℃,年平均降水量500 mm左右,年平均蒸发量1 420.2 mm,大气相对湿度69%,年有效积温2 659.8 ℃,年均日照时数2 098.7 h,无霜期180 d,生长期180 d。境内土壤以山地褐土和棕壤为主,土层厚度约50 cm,pH值5.0~7.0,酸性至中性反应,为土石侵蚀剥蚀中山山地地貌类型,腐殖质含量较少。林场地处小陇山林区中部,森林覆盖率达90%以上,各林分及其分布在林区具有代表性。
据全林调查资料,林区华山松人工林累计营造面积约为1×104hm2,其中郁闭成林占50%左右,是林区重要的林木商品资源。研究样地所在的华山松人工林林龄为34 a,初植密度3 000株/hm2,并先后于1994-1996年、2004-2006年和2013-2014年,在不同区域进行过3次间密留匀的间伐处理。
2 研究方法
2.1 样地设置
2016-07在海拔为1 600~1 850 m的华山松人工林分布区域,分别在阴坡和阳坡布设间伐强度为弱度(20%)、中度(30%)和强度(40%)的典型调查样地各4个,以未间伐的样地(郁闭度为0.9)为对照。为保证调查的准确性,尽量保持样地条件的一致性。每个固定样地的调查面积设置为20 m×20 m,用于记录样地特征(表1);同时沿对角线分别布设面积为5 m×5 m的次样方3个,用于林下微生境和更新幼苗幼树的调查。
2.2 调查方法与数据处理
2.2.1 调查内容及其方法 (1)微生境特性。选择调查前7 d晴朗无雨的天气,在各间伐处理样方内,由多人分别于10∶00-16∶00每隔2 h测定距地面0.2 m高处的光照强度(ZDS-10型照度计)、空气温度和湿度(DHM2型通风干湿温度计),尽量保证测定时间的同一性和即时性;用环刀法取0~30 cm土层土壤,每10 cm一层,密封带回用烘干法(65 ℃,48 h)测定土壤含水率;测量样方内地表凋落物的厚度,同时分物种调查灌草植物的盖度[14]。各生境因子取平均值。(2)幼苗幼树数量。在5 m×5 m的样方内,分别统计各样地林下华山松幼苗(Seedling,树高≤1 m)和幼树(Sapling,树高>1 m且胸径<4 cm)的更新数量,取平均值[15],并以幼苗幼树更新总数量占其种群总株数的比例衡量各样地华山松种群更新潜力大小。

表1 小陇山华山松人工林不同间伐强度样地基本特征Table 1 Basic characteristics of Pinus armandii plantation plots in Xiaolongshan under different thinning intensities
2.2.2 数据处理 使用Excel软件进行调查数据的初步计算,采用SPSS18.0软件对林内的微生境因子进行单因素方差分析(one-way ANONA),对幼苗和幼树的数量进行多因素方差分析(multiway ANOVA),采用通径分析方法对幼苗幼树更新数量与微生境的关系进行综合分析。
3 结果与分析
3.1 不同间伐强度华山松人工林下微生境特性
由表2可知,不同间伐强度华山松人工林下呈现出明显的生境异质特性,首先各样地林内光照强度有较大差异,随着间伐强度的增大,林下光照强度呈增大趋势,且阳坡环境显著优于阴坡环境。林下空气温度保持在22~23 ℃,各间伐样地之间差异不显著。林下空气湿度表现为强度间伐样地显著小于其他样地;同一间伐强度下阴坡大于阳坡样地,但差异并不显著。林下土壤含水率表现为中度间伐和强度间伐样地显著小于其他样地,但相同间伐强度下阴坡与阳坡环境之间差异不明显。阴坡环境下样地凋落物厚度随着间伐强度的增大而减少,而阳坡样地凋落物厚度则随间伐强度的增大呈波动状态。林下灌木层盖度表现为阴坡强度间伐样地最大、未间伐对照样地最小;阳坡各间伐样地之间差异不明显。林下草本层盖度表现为阴坡弱度间伐样地最大,阳坡未间伐样地最小。林下的灌草植物主要有平枝荀子(Cotoneasterhorizontalis)、鼠李(Rhamnusdavurica)、栓翅卫矛(Euonymusphellomanus)和胡枝子(Lespedezabicolor)等。
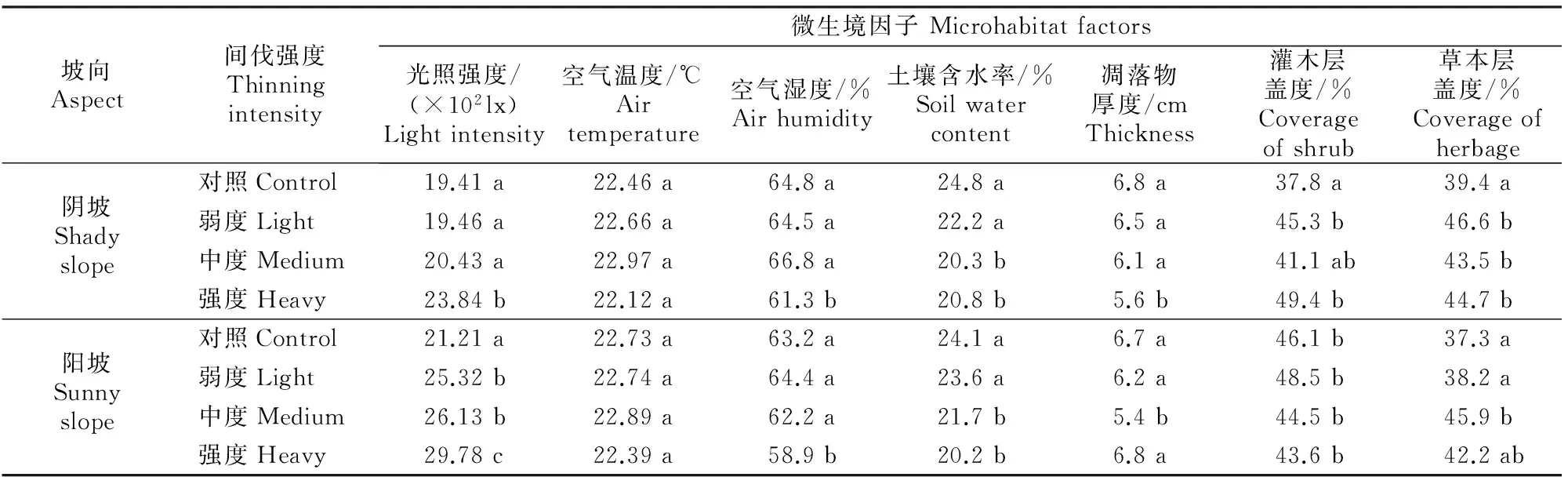
表2 不同间伐强度华山松人工林下的微生境特性Table 2 Microhabitat characteristics of Pinus armandii plantation under different thinning intensities
注:同列数据后标不同小写字母表示差异显著(P<0.05)。表3同。
Note:Different small letters in the same column mean significant difference atP<0.05 level.The same for table 3.
3.2 不同间伐强度华山松人工林下幼苗幼树天然更新数量
不同间伐强度华山松林下的天然更新状况可以反映幼苗幼树生长对微环境的适应特点。由图1可知,随着间伐强度的增大,不同坡向样地内的华山松幼苗更新数量均呈现明显的增大趋势,且间伐样地均显著(P<0.05)大于未间伐样地,如强度间伐样地内的幼苗数量是未间伐样地的4.69~5.28倍,但强度间伐与中度间伐样地之间幼苗数量差异并不显著。同一坡向的幼树数量均表现为未间伐的对照样地最少,但各样地之间并未表现出显著差异。多因素方差分析结果表明,坡向因子对幼苗和幼树的更新数量均未产生显著影响。由图1还可以看出,华山松幼苗向幼树生长过渡阶段存在明显的障碍现象,各样地内幼苗的数量均显著大于幼树数量,如强度间伐样地的幼苗数量是幼树的9.90~11.94倍,中度间伐样地的幼苗数量是幼树的10.85~13.59倍,弱度间伐样地的幼苗数量是幼树的8.06~9.14倍,未间伐样地幼苗数量是幼树的2.86~3.46倍。这说明华山松幼苗和幼树对微生境的生存适应机制可能有所不同,在幼树生长阶段死亡率较高。

图1 不同间伐强度下华山松人工林的更新数量Fig.1 Numbers of Pinus armandii plantation regeneration under different thinning intensities
3.3 不同间伐强度华山松人工林种群的天然更新潜力
相比天然林,华山松人工林处于先锋群落演替的初级阶段[16],其种群更新潜力指标是评价华山松林分更新能力大小的重要依据。由表3可知,阳坡和阴坡环境下,抚育间伐措施均对华山松种群的更新潜力有显著影响(P<0.05),且随着间伐强度的增大呈上升趋势。间伐后,样地内幼树幼苗的数量明显增加,但同时成年树的数量明显降低。阴坡环境下强度间伐、中度间伐和弱度间伐样地的种群更新潜力为对照样地的2.06,1.92和1.70倍,阳坡环境下则为2.04,1.98和1.83倍。可以看出,强度间伐对华山松人工林种群的更新潜力贡献最大,但阴坡与阳坡环境之间没有显著差异。
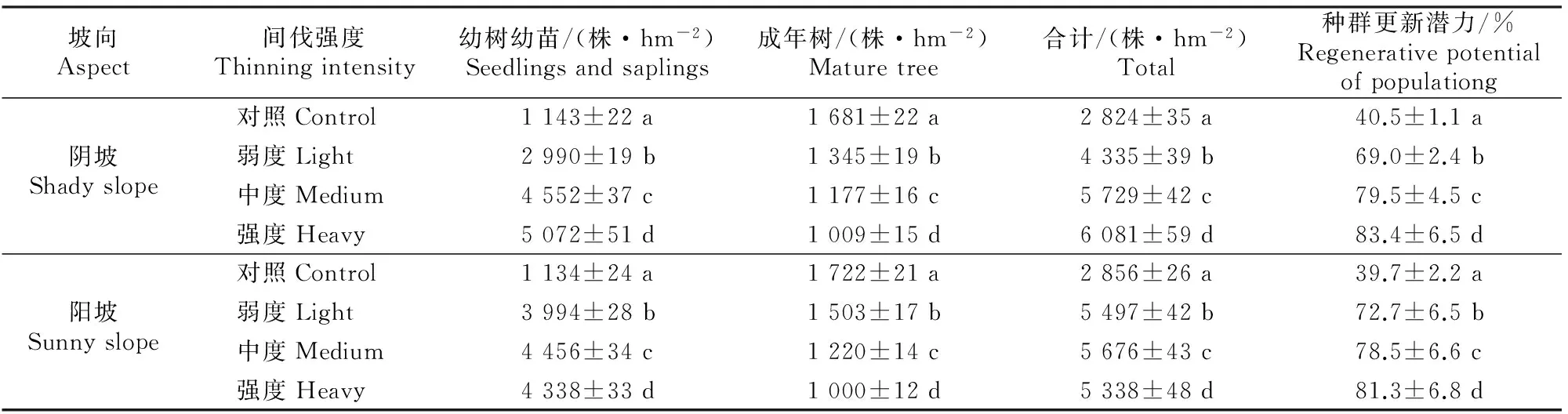
表3 不同间伐强度下华山松人工林种群的更新潜力Table 3 Regenerative potential of Pinus armandii population under different thinning intensities
3.4 不同间伐强度华山松人工林下更新与微生境间的关系
3.4.1 对幼苗更新数量的影响 通径分析可以反映出自变量(微生境因子)对因变量(幼苗数量)的解释程度及直接和间接重要性。由前述分析可知,不同坡向间伐处理林下呈现出明显的生境异质特性,这是促使林下幼苗更新数量产生差异的直接原因。通径分析结果(表4)表明,R2>0.95,说明表4中的7项微生境因子基本可以解释幼苗数量的变异现象,其中土壤含水率和地表凋落物厚度对幼苗的更新数量起主要负向作用,其相关系数分别为-0.847和-0.692,且这2项环境因子的直接通径系数也均为负数,分别为-0.507和-0.220,因而可以推断土壤含水率和地表凋落物厚度为影响华山松幼苗更新的主要消极因素。林下光照强度和灌草层盖度对幼苗数量起正向作用,有利于华山松幼苗的更新,其中光照强度的直接作用更大(直接通径系数为0.843),而灌草盖度的间接作用则较大,如灌木层盖度主要是通过影响林下光照强度(间接通径系数为0.293)和空气湿度(间接通径系数为-0.303)而综合发挥作用的。

表4 不同间伐强度华山松人工林下微生境因子与幼苗数量的通径分析结果Table 4 Path analysis of microorganism factors and seedlings in Pinus armandii plantation under different thinning intensities
注:X1.光照强度;X2.空气温度;X3.空气湿度;X4.土壤含水率;X5.凋落物厚度;X6.灌木层盖度;X7.草本层盖度。间接通径系数下的数据,从纵列看,X1与X2交汇点的数据为-0.175,其表示X2通过影响X1而对因变量(幼苗数量)所起的间接作用大小;从横列看,X1与X2交汇点的数据为0.071,其表示X1通过影响X2对因变量(幼苗数量)所起的间接作用大小;其余依此类推。表5同。
Note:X1.Light intensity;X2.Air temperature;X3.Air humidity;X4.Soil water content;X5.Thickness;X6.Coverage of shrub;X7.Coverage of herbage.The data of the indirect path coefficient from the column,the data at the intersection ofX1andX2is -0.175,indicating the indirect effect ofX2on the dependent variable number of seedlings by affectingX1;From the horizontal,the data at the intersection ofX1andX2is 0.071,indicating the indirect effect ofX1on the dependent variable number of seedlings by affectingX2.The rest and so on.The same for table 5.
3.4.2 对幼树更新数量的影响 运用同样的方法分析林下微生境对华山松幼树存活数量的直接和间接影响。通过前述的分析可知,华山松幼苗向幼树过渡阶段存在明显的障碍现象,幼树与幼苗对微生境的适应机制可能有所不同。通径分析结果(表5)表明,R2>0.95,说明这7项微生境因子同样可以解释幼树数量的变异现象,其中空气湿度、土壤含水率和凋落物厚度为华山松幼树生长的主要消极影响因素,但由于其直接通径系数均为正数,因而可推断这3个因子是通过影响其他环境因子而间接起负向作用的。光照强度对华山松幼树存活发挥着重要的正向作用,且直接通径系数(1.121)较大,从而对促进林下天然更新具有积极意义。除此之外,林下灌草层盖度也对幼树的存活发挥着正向作用,其相关系数分别为0.697和0.326,但直接作用较光照强度明显偏小(直接通径系数分别为0.487和0.366)。由此可以推断,林下光照强度条件制约是华山松幼树能否存活的关键,也是影响华山松幼苗向幼树正常生长过渡的重要环境因子。
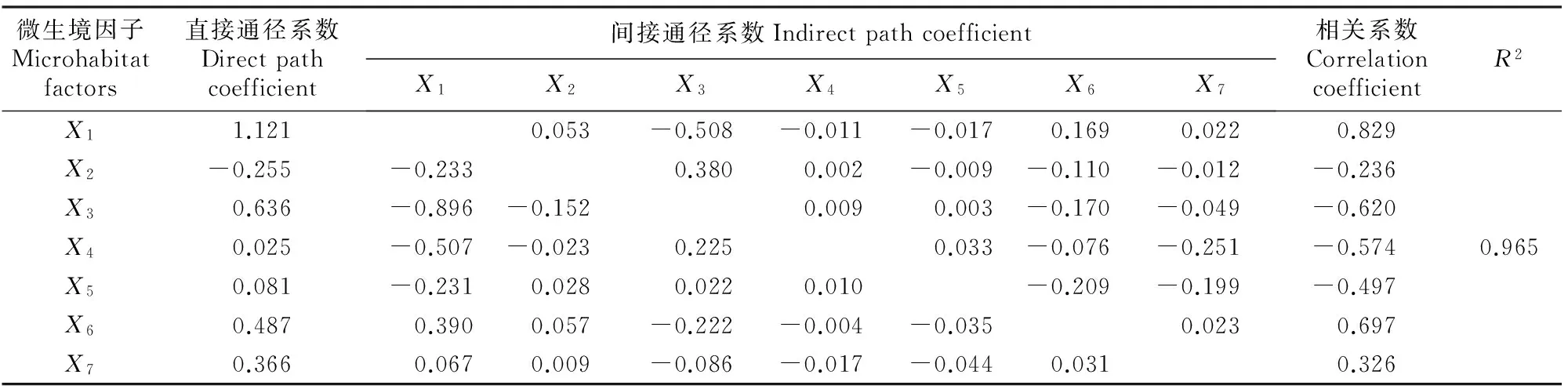
表5 不同间伐强度华山松人工林下微生境因子与幼树数量的通径分析结果Table 5 Path analysis of microorganism factors and saplings in Pinus armandii plantation under different thinning intensities
4 结论与讨论
4.1 微生境异质性对华山松幼苗数量有显著影响
幼苗的生长发育需要适宜的环境条件作为支撑,各微生境因子对华山松幼苗存活发挥着不同作用,以强度间伐林下所表现出的综合效应最为明显。本研究表明,土壤含水率和凋落物厚度是制约华山松幼苗数量的主要环境因子,较高的土壤含水率和较厚的地表凋落物对幼苗数量起主要的消极影响,且这两项因子的直接作用均较大。这可能是由于过高的土壤含水率会抑制华山松种子的正常萌发[17],从而影响其幼苗发生数量;而过厚的地表凋落物则会阻碍种子接触土壤[18],种子受真菌等微生物感染霉变的几率也相应增大[19],从而减少了幼苗定居的机会。调查过程中也确实发现在较厚凋落物层萌发的幼苗根系并未深插入土壤中。本研究同时表明,灌草层盖度和光照强度因子会对华山松幼苗的发生数量起积极作用,这可能是由于较大的灌草盖度为其种子萌发提供了较好的庇护条件,易形成稳定适宜的小气候环境,而充足的光照强度则直接促进了定居幼苗的进一步生长发育。
4.2 华山松幼苗向幼树生长过渡存在明显障碍现象
调查发现,各生境条件下华山松幼苗的数量均显著大于其幼树数量,说明幼苗和幼树生长对林下微生境的适应性有所不同,幼苗向幼树过渡阶段死亡率较高。通径分析表明,土壤含水率和凋落物厚度虽然对华山松幼树生长起着消极作用,但其直接通径系数反而为正数,说明这两项环境因子所起的消极影响仅是通过其他因子而表现出的间接作用;空气温、湿度因子对幼苗、幼树数量的相关系数虽然也为负数,但由于二者具有日周期性的特点(即早晚波动较大),因而也不能被解读为是制约华山松生长发育的关键因子。而松属植物通常被认为是群落生态系统中的先锋树种[20],其生长发育过程需要良好的光照条件,通径分析结果也表明光照强度对华山松幼树的存活发挥着重要作用,因而光照强度可以被认为是影响华山松幼苗向幼树过渡成功的关键因子。随着华山松幼苗的不断发育,其对光照条件的要求变得更为苛刻,林下相对较为郁闭的微生境环境不能满足幼树的进一步生长,天然更新过程因此受阻。
本研究表明,适宜的林下微生境条件对维持华山松天然更新至关重要,土壤含水率和凋落物厚度是制约幼苗定居与发育的关键因子,而充足的光照强度则是幼苗向幼树成功过渡的重要保障。抚育间伐能够大幅改善林内的光照强度,提高华山松幼树的成活率;通过清理林下凋落物,亦可为其幼苗的成功定居和生长发育创造良好条件,有利于种群更新潜力的提升。因此,在华山松人工林的抚育管理过程中,应注重采用适度间伐与清理林下凋落物相结合的经营措施,有助于林分持续、稳定地生长。但本试验仅是基于静态视角来探讨森林微环境对幼苗、幼树存活数量的影响,不能完全反映跨时间尺度下华山松人工林的天然更新动态规律特征,因而未来有必要进一步加强在此方面的跟踪研究。
[1] 管云云,费 菲,关庆伟,等.林窗生态学研究进展 [J].林业科学,2016,52(4):91-99.
Guan Y Y,Fei F,Guan Q W,et al.Advances in studies of forest gap ecology [J].Scientia Silvae Sinicae,2016,52(4):91-99.
[2] 胡会峰,刘国华.人工油松林恢复过程中土壤理化性质及有机碳含量的变化特征 [J].生态学报,2013,33(4):1212-1218.
Hu H F,Liu G H.Dynamics of soil physical-chemical properties and organic carbon content along a restoration chronosequence inPinustabulaeformisplantation [J].Acta Ecologica Sinica,2013,33(4):1212-1218.
[3] 李 龙,姚云峰,秦富仓,等.黄花甸子流域人工林土壤有机碳密度分布特征 [J].西北农林科技大学学报(自然科学版),2016,44(2):77-82.
Li L,Yao Y F,Qin F C,et al.Distribution characteristics of soil organic carbon density of different forests in Huanghuadianzi watershed [J].Journal of Northwest A&F University(Natural Science Edition),2016,44(2):77-82.
[4] Oberhauser U.Secondary forest regeneration beneath pine (Pinuskesiya) plantations in the northern Thai highlands:a chronosequence study [J].Forest Ecology & Management,1997,99(1):171-183.
[5] Hertel D,Wesche K.Tropical moistPolylepis,stands at the treeline in East Bolivia:the effect of elevation on stand microclimate,above- and below-ground structure,and regeneration [J].Trees,2008,22(3):303-315.
[6] Cierjacks A,Rühr N K,Wesche K,et al.Effects of altitude and livestock on the regeneration of two tree line formingPolylepisspecies in Ecuador [J].Plant Ecology,2008,194(2):207-221.
[7] D’Amato A W,Orwig D A,Foster D R.Understory vegetation in old-growth and second-growthTsugacanadensis,forests in western Massachusetts [J].Forest Ecology & Management,2009,257(3):1043-1052.
[8] 朱教君,刘足根,王贺新.辽东山区长白落叶松人工林天然更新障碍分析 [J].应用生态学报,2008,19(4):695-703.
Zhu J J,Liu Z G,Wang H X.Obstacles for natural regeneration ofLarixolgensisplantations in montane regions of eastern Liaoning Province,China [J].Chinese Journal of Applied Ecology,2008,19(4):695-703.
[10] 王中磊,高贤明.锐齿槲栎林的天然更新:坚果、幼苗库和径级结构 [J].生态学报,2005,25(5):986-993.
Wang Z L,Gao X M.The regeneration ofQuercusalienavar.acuteserrata:acorn status,seedling pool and size structure [J].Acta Ecologica Sinica,2005,25(5):986-993.
[11] Mühlenberg M,Appelfelder J,Hoffmann H,et al.Structure of the montane taiga forests of West Khentii,Northern Mongolia [J].Journal of Forest Science,2012(2):45-56.
[12] Dobrowolska D.Effect of stand density on oak regeneration in flood plain forests in Lower Silesia,Poland [J].Forestry,2008,81(4):511-523.
[13] 张明霞,王得祥,康 冰,等.秦岭华山松天然次生林优势种群的种间联结性 [J].林业科学,2015,51(1):12-21.
Zhang M X,Wang D X,Kang B,et al.Interspecific associations of dominant plant populations in secondaryforest ofPinusarmandiiin Qinling mountains [J].Scientia Silvae Sinicae,2015,51(1):12-21.
[14] 韩文娟,袁晓青,张文辉.油松人工林林窗对幼苗天然更新的影响 [J].应用生态学报,2012,23(11):2940-2948.
Han W J,Yuan X Q,Zhang W H.Effects of gap size on seedling natural regeneration in artificialPinustabulaeformisplantation [J].Chinese Journal of Applied Ecology,2012,23(11):2940-2948.
[15] 于 倩,谢宗强,熊高明,等.神农架巴山冷杉(Abiesfargesii)林群落特征及其优势种群结构 [J].生态学报,2008,28(5):1931-1941.
Yu Q,Xie Z Q,Xiong G M,et al.Community characteristics and population structure of dominant species ofAbiesfargesiiforests in Shennongjia National Nature Reserve [J].Acta Ecologica Sinica,2008,28(5):1931-1941.
[16] 彭舜磊,王得祥.秦岭火地塘林区华山松人工林与天然次生林群落特征比较 [J].西北植物学报,2009,29(11):2301-2311.
Peng S L,Wang D X.Comparison of community characteristics of plantation and secondary forest ofPinusarmandiiin Huoditang forest region of Qinling mountain [J].Acta Botanica Boreali-Occidentalia Sinica,2009,29(11):2301-2311.
[17] 兰国玉,雷瑞德,安 锋,等.秦岭中段华山松群落动态初步研究 [J].西北林学院学报,2005,20(3):48-52.
Lan G Y,Lei R D,An F,et al.Study on community dynamic ofPinusarmandiiin the Qinling mountains [J].Journal of Northwest Forestry University,2005,20(3):48-52.
[18] Brewer S W,Webb M A H.Ignorant seed predators and factors affecting the seed survival of a tropical palm [J].Oikos,2001,93(1):32-41.
[19] 王贺新,李根柱,于冬梅,等.枯枝落叶层对森林天然更新的障碍 [J].生态学杂志,2008,27(1):83-88.
Wang H X,Li G Z,Yu D M,et al.Barrier effect of litter layer on natural regeneration of forests: a review [J].Chinese Journal of Ecology,2008,27(1):83-88.
[20] 柴宗政,王得祥,郝亚中,等.秦岭中段华北落叶松人工林演替动态 [J].林业科学,2014,50(2):14-21.
Chai Z Z,Wang D X,Hao Y Z,et al.Succession dynamics ofLarixprincipis-rupprechtiiplantation in intermediate section of Qinling mountains [J].Scientia Silvae Sinicae,2014,50(2):14-21.
