沙柳的孔隙结构、微纤丝角和纤维素结晶度研究
2018-02-27周贤武邓丽萍王玉荣吕建雄赵荣军
周贤武,邓丽萍,王 滋,王玉荣,吕建雄,赵荣军
(中国林业科学研究院 木材工业研究所,北京 100091)
沙柳(Salixpsammophila)是生长在中国北方沙漠和草原的一种旱生落叶灌木或小乔木,它具有发达的根系,因此能够在缺水的环境下生长[1]。沙柳生长迅速,3年就可以长到3~5 m高,一般每间隔3年进行一次平茬,这有利于沙柳萌发大量的枝条[2]。据统计,内蒙古的库布齐沙漠每3年可收获5.2 t/hm2的沙柳材。然而,大部分沙柳木材被作为薪柴焚烧,造成大量浪费。近年来,已有研究将沙柳用于纤维板、刨花板、复合板、纸浆、纤维素产物和液化产物(如生物乙醇)等的生产[3-6],这为沙柳的高附加值加工利用开辟了道路。
孔隙结构、微纤丝角和纤维素结晶度对木材产品的质量有重要影响。孔隙结构对木材密度有显著影响,孔隙度小则密度大[7],而密度又是影响木材材性的重要因素,因此研究木材孔隙结构对深入了解木材性质具有重要作用。微纤丝角(MFA)是指木材细胞壁S2层中微纤丝方向与细胞主轴之间的夹角,它与木材的宏观物理力学性质密切相关,是评定材质、纸张强度、纤维复合材料性能等的重要指标,对于木质材料的加工利用有重要的研究价值[8]。纤维素结晶度(CrI)是指在具有结晶区和非结晶区(无定形区)的纤维素微纤丝中,结晶区占纤维素微纤丝整体的百分率,它与树木的生长期、纤维种类、纤维长度和纤维介电常数等均有密切关系,并对木材的纤维抗拉强度、杨氏模量、尺寸稳定性及化学反应性等具有重要的影响[9]。利用木质纤维素生物质制取燃料乙醇已经成为近年来研究的热点,比表面积可及性和纤维素结晶度是影响木质纤维素生物质酶水解的两大主要因素,其中比表面积可及性最为重要[10],提高比表面积可及性和降低纤维素结晶度可以有效提高酶解转化率[11]。
综上所述,研究沙柳孔隙结构、微纤丝角和纤维素结晶度,对于沙柳的水解、制浆造纸、人造板生产等资源高附加值加工利用具有一定的指导意义。目前仅冯利群[12]采用偏光显微镜法测量了沙柳的MFA,对于沙柳原料纤维素结晶度的研究也较少,而对于沙柳孔隙结构以及以上3个指标的径向变异研究则未见报道。大量研究已经表明,年轮对木材性质有显著的影响。因此,本研究采用氮气吸附法(NAD)和X射线衍射(XRD)研究了沙柳木材的孔隙结构、微纤丝角和纤维素结晶度,并深入探讨了年轮对沙柳木材孔隙结构、微纤丝角和纤维素结晶度的影响,以期为沙柳材的加工利用提供理论依据。
1 材料与方法
1.1 材 料
以采自内蒙古鄂尔多斯沙地的3年生沙柳为试验材料。沙区年平均气温6.0~8.5 ℃,年降水量250~440 mm,降水集中于7-9月份,占全年降水量的60%~75%,尤以8月份为多。
取3株沙柳,从每株地上1 m处开始沿树根到树梢方向先取100 mm长木段用于氮气吸附法(NAD)表征孔隙结构,再取170 mm长用于X射线衍射(XRD)测微纤丝角,最后取100 mm长用于X射线衍射(XRD)测纤维素结晶度。所有取下来的木段去除表皮、韧皮部和髓心后备用,具体取样和试样制备方法见图1。
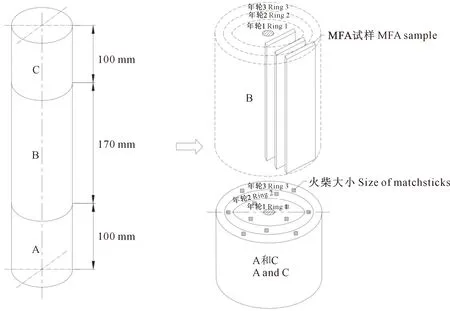
图1 沙柳材取样和试样锯解示意Fig.1 Sample sawing sketch of Salix psammophila wood
1.2 测定项目与方法
1.2.1 木材孔隙结构表征 将取下来的相应木段按年轮分开,劈成火柴大小,并磨成粒径0.175~0.246 mm(60~80目)的木粉,采用氮气吸附法表征沙柳不同年轮处木材的孔隙结构。样品在测量前置于80 ℃下真空脱气10 h,然后使用美国Micromeritics公司生产的ASAP 2020型比表面积及孔径分析仪获得不同年轮沙柳材的氮气吸附-脱附等温线,测定时以氮气作为吸附介质,试验温度为-196 ℃。最后依据苌姗姗等[13]提到的方法分别计算沙柳比表面积和孔体积,并根据沙柳材的氮气吸附-脱附等温线绘制孔径-孔容关系曲线。
1.2.2 微纤丝角(MFA)测定 首先将每株170 mm长的木段截成5段,然后在每段中由髓心向树皮方向按年轮依次截取尺寸为1.5 mm (R)×10 mm (T)×30 mm (L)的试样,如图1,共获得45个试样,每个年轮15个。试样制备好后,将它们置于恒温恒湿箱中,在温度为20 ℃、相对湿度为65%的条件下平衡1周。再用X′Pert-PRO型X射线衍射仪测量沙柳材微纤丝角(MFA),试验获得的每一张XRD双峰图用GaussAmp公式进行拟合,拟合范围为45°~135°和225°~315°,拟合方程为[14]:
式中:y表示衍射强度,x表示样品台旋转角度,a是常数,b1和b2是峰值,u和u+180是峰值所对应的中心横坐标,σ1和σ2是半峰宽。按照0.6T法计算沙柳材的MFA,即T=σ1+σ2,MFA=0.6T[15]。
1.2.3 纤维素结晶度(CrI)测定 用XRD对相对结晶度进行测定。首先将相应木段按年轮分开,劈成火柴大小,并磨成粒径0.175~0.246 mm (60~80目)的木粉,然后与MFA试样相同平衡含水率1周。测试时,在XRD模式下,记录2θ角范围为5°~40°,扫描速率为0.05 (°)/s。相对结晶度(CrI)按以下公式进行计算[16]:
CrI=100×(I002-Iam)/I002。
式中:I002代表晶格衍射角的极大强度,一般在2θ=22°出现;Iam表示衍射角为18°时非结晶背景衍射的散射强度。
1.3 统计分析
采用单因素方差分析法和多重比较(S-N-K test),对各年轮间的纤维素结晶度和微纤丝角进行差异显著性分析及两两比较。
2 结果与分析
2.1 沙柳材的孔隙结构
由表1可知,沙柳材的比表面积平均为1.01 m2/g。3个年轮处沙柳材的比表面积和孔体积从髓心到树皮依次增大,第3年轮木材比表面积是第1年轮的3.4倍,第3年轮木材的孔体积更是远大于第1年轮,为第1年轮的780倍。
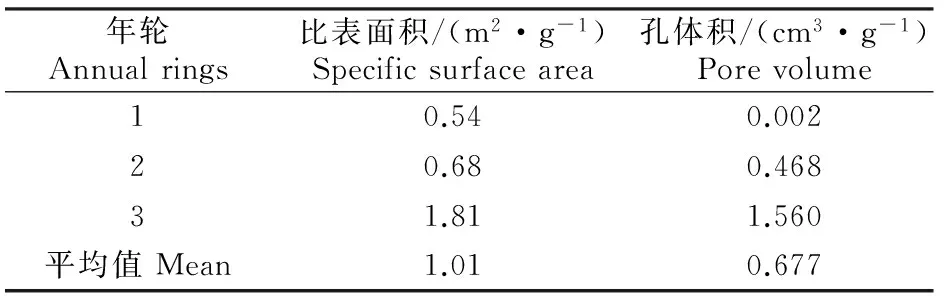
表1 不同年轮沙柳材的比表面积和孔体积Table 1 Specific surface area and pore volume of Salix psammophila at different annual rings
图2中3个年轮沙柳材均属于Ⅱ和Ⅳ混合型氮气吸附-脱附等温线。由图2可知,3个年轮木材氮气吸附量的上升速率明显表现为由髓心到树皮递增,说明靠近树皮的木材具有更多的孔隙结构。当相对压力为0~0.1时,3个年轮木材的氮气吸附量均迅速增加,此阶段主要发生微孔(孔径<2 nm)的填充,表明沙柳材中有一定的微孔结构;当相对压力为0.1~0.9时,第1年轮和第2年轮木材氮气吸附量随相对压力的增加仍缓慢增加,而第3年轮木材氮气吸附量增加明显比前2个年轮快,此阶段发生介孔(孔径为2~50 nm)和大孔(孔径>50 nm)的多层吸附,表明沙柳材中有一定的介孔和大孔结构,而且第3年轮木材介孔和大孔结构明显多于前2个年轮;在相对压力达到0.9时,等温线开始急剧上升,表明沙柳材中具有一定量的大孔。图3是沙柳不同年轮木材孔径大小分布图。由图3可知,第3年轮木材的孔体积明显大于其他2个年轮,第3年轮木材中大部分为介孔孔隙,且3个年轮木材大部分孔径都在2~10 nm。
2.2 沙柳材的微纤丝角(MFA)
图4为沙柳微纤丝角X射线衍射图,是典型的微纤丝角双峰图。由图4可以看出,从髓心到树皮,峰型越来越细长,使得MFA逐渐减小。表2为沙柳3个年轮木材的MFA计算结果。由表2可知,沙柳的MFA变化于10.71°~14.35°,平均为12.41°,略大于偏光显微镜法测得的沙柳MFA(11.09°),但低于一般正常阔叶树材15°~25°的MFA值[12]。
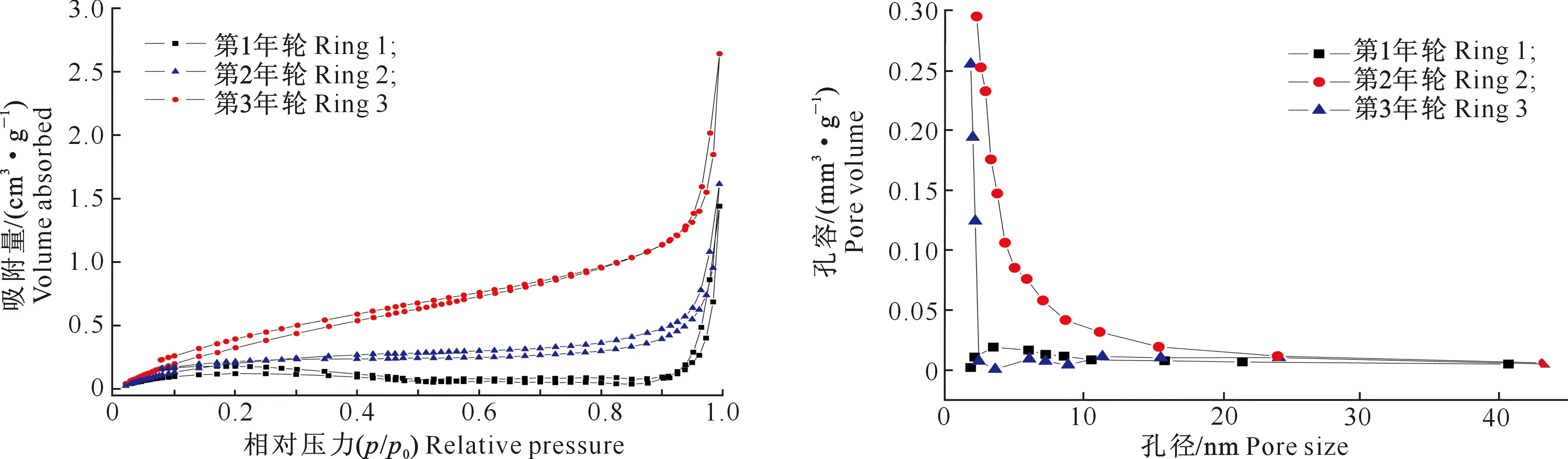
图2 不同年轮沙柳材的氮气吸附-脱附等温线Fig.2 Nitrogen adsorption-desorption isotherm of Salix psammophila at different annual rings
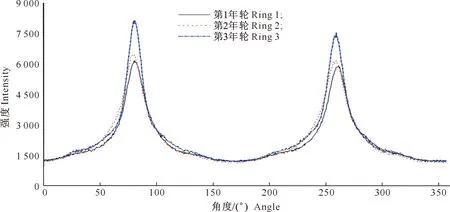
图4 不同年轮沙柳材微纤丝角的X射线衍射图Fig.4 X-ray diffraction spectrum of microfibril angle of Salix psammophila at different annual rings
2.3 沙柳材的纤维素结晶度
图5是沙柳不同年轮木材纤维素结晶度的X射线衍射图。由图5可知,3个年轮木材在2θ=18°附近的衍射极小峰值Iam基本重叠,但在2θ=22°附近的衍射极大峰值I002表现为从髓心到树皮缓慢降低。由表2可知,沙柳材纤维素结晶度变化于48.15%~49.58%,平均为48.99%。
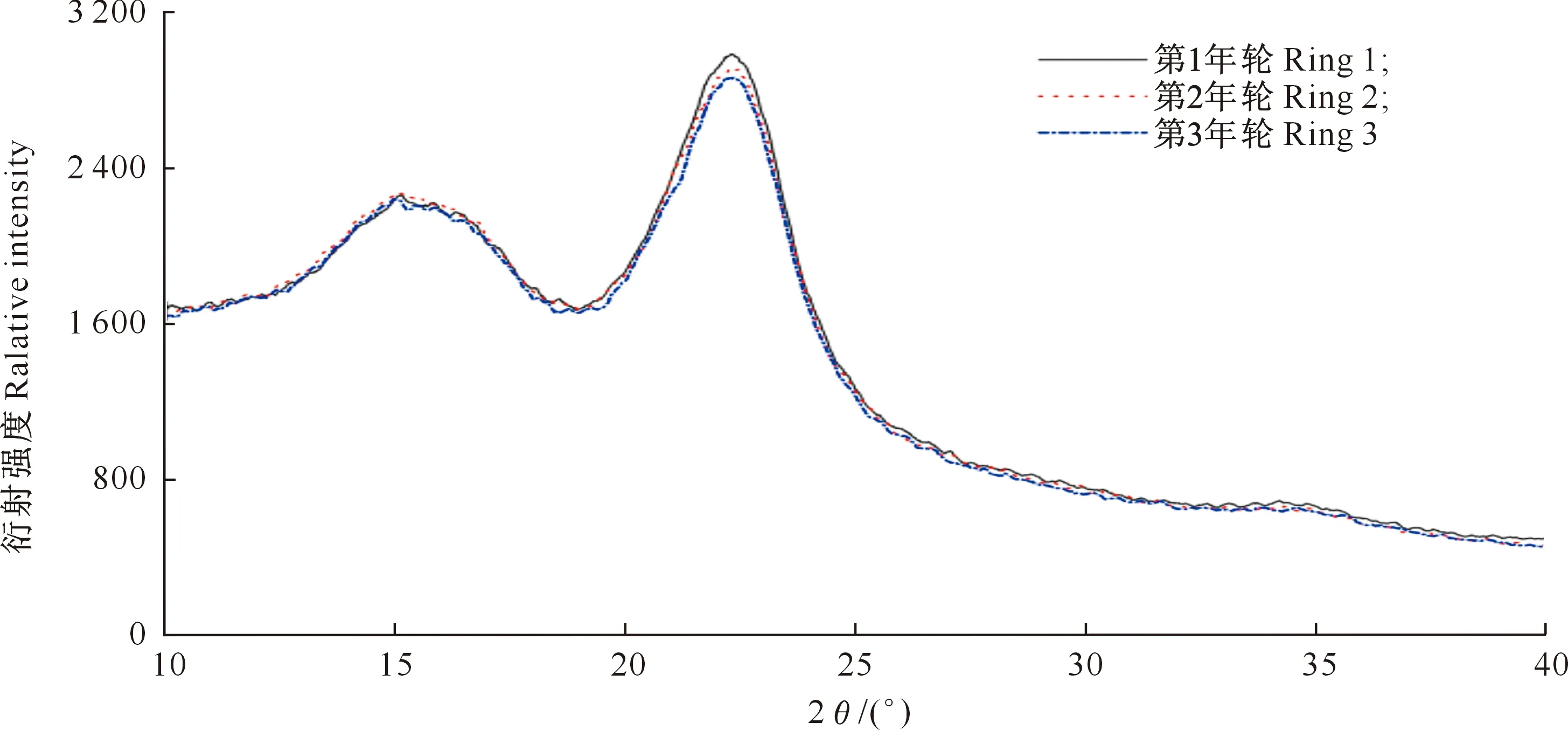
图5 不同年轮沙柳材纤维素结晶度的X射线衍射图Fig.5 X-ray diffraction spectrum of cellulose crystallinity of Salix psammophila at different annual rings

年轮Annualrings微纤丝角/(°)MFA纤维素结晶度/%CrI114.35±1.14a48.15±1.34a212.17±1.85ab49.23±0.68a310.71±0.98b49.58±1.94a
注:表中数据为“平均值±标准差”;同列数据后标不同小写字母表示在P=0.05水平具有显著差异。
Note:Digitals are “average±standard deviation”.Different lowercase letters in same line indicates significant difference between annual rings atP=0.05.
3 讨 论
孔隙结构直接影响木材细胞壁的物理力学性质,孔隙结构的增多会降低木材的细胞壁密度和细胞壁力学强度。目前对于木材细胞壁孔隙结构的研究极少。本研究首次采用NAD方法测量沙柳细胞壁的孔隙结构,发现沙柳前2个年轮木材的比表面积低于苌姗姗等[13]测得的正常杨木木材的比表面积(1.7 m2/g),因此沙柳材相比杨木具有更致密的胞壁结构。靠近髓心的沙柳木材因为具有更致密的胞壁结构,因而具有较高的木材细胞壁力学强度。纤维素原料的溶解是用木材生产纤维素材料和生物质能源的必要过程,原料孔隙结构越多,比表面积越大,越利于溶剂与原料的充分接触,原料溶解也越快[10-11]。由以上分析可知,靠近树皮处的沙柳材有较大的比表面积可及性,是制取纤维素材料和生物质能源较好的原材料。
前人虽采用偏光显微镜法对沙柳材的微纤丝角进行了测量,但本试验采用更先进的XRD技术对沙柳材微纤丝角的径向变异进行了研究,发现这种变异表现为从髓心到树皮逐渐减小,其中第3年轮木材的微纤丝角与第1年轮具有显著差异,相对第1年轮减小了25.4%。据前人研究可知,木材微纤丝角由髓心到树皮逐渐减小[17-19]。由此可见,沙柳材微纤丝角的径向变异符合一般木材微纤丝角的变异规律。MFA对木材性质和尺寸稳定性有很大影响,MFA小的木材顺纹方向强度高,尺寸稳定性好[20]。因此,可以预测沙柳顺纹方向的强度和尺寸稳定性应优于相同密度的一般阔叶树材;靠近树皮的沙柳木材微纤丝角小,可以增强木材顺纹方向强度和尺寸稳定性,其具体变化规律还需结合孔隙结构和纤维素结晶度等进一步研究证明。
本研究首次对沙柳不同年轮木材纤维素结晶度进行研究,发现其纤维素结晶度从髓心到树皮缓慢增加,与前人研究的木材纤维素结晶度径向变化趋势一致[21-22]。表现为这种趋势的原因是在植物生长发育过程中,纤维素结晶度也在不断提高[23]。通过方差分析发现,XRD测量的沙柳材纤维素结晶度在3个年轮间没有显著差异,因此在加工利用沙柳木材时可以忽略年轮对纤维素结晶度的影响。
4 结 论
沙柳3个年轮木材的比表面积从髓心到树皮分别为0.54,0.68和1.81 m2/g,孔体积分别为0.002,0.468和1.560 cm3/g,比表面积和孔体积从髓心到树皮均逐渐增大,尤其靠近树皮的木材比表面积和孔体积明显高于内部2个年轮的木材。沙柳材中存在微孔、介孔和大孔结构,大部分的孔隙直径集中在2~10 nm。沙柳材的微纤丝角平均为12.41°,低于一般阔叶材,而且从髓心到树皮逐渐减小,靠近树皮的木材微纤丝角明显低于靠近髓心的木材。沙柳材的纤维素结晶度从髓心到树皮缓慢增大,但趋势平缓,差异不大。靠近树皮的沙柳材具有较大的比表面积、孔体积、纤维素结晶度和较小的微纤丝角,是制取纤维素材料和生物质能源的较好原材料。
[1] Kubo S,Hashida K,Makino R,et al.Chemical composition of desert willow (Salixpsammophila) grown in the Kubuqi Desert, Inner Mongolia,China:bark extracts associated with environmental adaptability [J].Journal of Agricultural and Food Chemistry,2013,61(50):12226-12231.
[2] 许 凤,Jones-Gwynn L L,孙润仓.速生灌木沙柳的纤维形态及解剖结构研究 [J].林产化学与工业,2006,26(1):91-94.
Xu F,Jones-Gwynn L L,Sun R C.Fibre morphology and anatomical structure of sandlive willow (Salixpsammophila) [J].Chemistry and Industry of Forest Products,2006,26(1):91-94.
[3] Xu F,Geng Z C,Lu Q,et al.Salixpsammophilapulp obtained by alkaline hydrogen peroxide process [J].Cellulose Chemistry and Technology,2006,40(1/2):93-107.
[4] Sun X F,Xiao B,Baird M S.Modification and characterization of fibers of three sandy willow shrub species [J].Forestry Studies in China,2006,8(3):16-21.
[5] Jiang J X,Tang Y,Wang K,et al.Influence of steam pretreatment time on chemical composition and simultaneous saccharification and fermentation for ethanol from pruning shrub stalks [J].Journal of Biobased Materials and Bioenergy,2011,5(2):258-264.
[6] 薛 玉,杨桂花,陈嘉川,等.沙柳特性及其综合利用 [J].华东纸业,2011,42(4):57-60.
Xue Y,Yang G H,Chen J C,et al.Salixpsammophilacharacteristics and comprehensive utilization [J].East China Pulp and Paper Industry,2011,42(4):57-60.
[7] Ding W D,Koubaa A,Chaala A,et al.Relationship between pore structure,wood density and methyl methacrylate impregnation rate [J].Wood Material Science and Engineering,2008,3(1/2):62-70.
[8] 冯利群,马 蓓,侯玲艳.宽叶水柏枝材构造及微纤丝角研究 [J].安徽农业科学,2008,36(31):13636-13638.
Feng L Q,Ma B,Hou L Y.Study on the wood structure and microfibril angle ofMyricariaplatyphyllaMaxim [J].Journal of Anhui Agricultural Sciences,2008,36(31):13636-13638.
[9] 李 坚.木材波谱学 [M].北京:科学出版社,2003:20-27.
Li J.Wood spectroscopy [M].Beijing:Science Press,2003:20-27.
[10] Alvira P,Tomás-Pejó E,Ballesteros M,et al.Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis:a review [J].Bioresource Technology,2010,101(13):4851-4861.
[11] Sun Y,Cheng J.Hydrolysis of lignocellulosic materials for ethanol production:a review [J].Bioresource Technology,2002,83(1):1-11.
[12] 冯利群.4种灌木材微纤丝角及其变异性研究 [J].安徽农业科学,2008,36(7):2747-2748.
Feng L Q.Study on the microfibril angle in four species of shrubs and its variability [J].Journal of Anhui Agricultural Sciences,2008,36(7):2747-2748.
[13] 苌姗姗,胡进波,Clair Bruno,等.氮气吸附法表征杨木应拉木的孔隙结构 [J].林业科学,2011,47(10):134-140.
Chang S S,Hu J B,Clair B,et al.Pore structure characterization of poplar tension wood by nitrogen adsorption-desorption method [J].Scientia Silvae Sinicae,2011,47(10):134-140.
[14] 余 雁,王 戈,覃道春,等.X射线衍射法研究毛竹微纤丝角的变异规律 [J].东北林业大学学报,2007,35(8):28-29.
Yu Y,Wang G,Qin D C,et al.Variation in microfibril angle of moso bamboo by X-ray diffraction [J].Journal of Northeast Forestry University,2007,35(8):28-29.
[15] Cave I D.Theory of X-ray measurement of microfibril angle in wood [J].Forest Products Journal,1966,16(10):37-42.
[16] Segal L,Creely J J,Martin A E,et al.An empirical method for estimating the degree of crystallinity of native cellulose using the X-ray diffractometer [J].Textile Research Journal,1959,29(10):786-794.
[17] McMillin C W.Fibril angle of loblolly pine wood as related to specific gravity,growth rate,and distance from pith [J].Wood Science and Technology,1973,7(4):251-255.
[18] 汪佑宏,刘杏娥,江泽慧,等.利用X射线衍射技术分析黄藤材微纤丝角、结晶度及γ射线的影响 [J].光谱学与光谱分析,2010(5):1404-1407.
Wang Y H,Liu X E,Jiang Z H,et al.Analysis of the microfibrilla angles & crystallinity index ofDaemonoropsmargaritaewith X-ray diffractometry and the effects of γ-ray irradiation [J].Spectroscopy and Spectral Analysis,2010(5):1404-1407.
[19] 费本华,阮锡根.银杏木材微纤丝角及其与生长轮密度相关模型的建立 [J].木材工业,2000,14(3):13-15.
Fei B H,Ruan X G.Study on wood microfibril angle and the correlation models with growth ring density of ginkgo [J].China Wood Industry,2000,14(3):13-15.
[20] 江泽慧,费本华.长江滩地不同品系杨树木材纤维形态、微纤丝角和结晶度变异研究 [J].安徽农学院学报,1992,19(4):255-262.
Jiang Z H,Fei B H.The variation of fiber form,microfibril angle and crystallinity of different strains of poplar wood in the flood land along the lower Yangtze River,China [J].Journal of Anhui Agricultural University,1992,19(4):255-262.
[21] 崔 凯,孙庆丰,廖声熙,等.翠柏木材解剖性质和结晶度的径向变异及化学性质 [J].东北林业大学学报,2012,40(4):49-54.
Cui K,Sun Q F,Liao S X,et al.Wood anatomical properties ofCalocedrusmacrolepisand radial variation and chemical property of crystallinity [J].Journal of Northeast Forestry University,2012,40(4):49-54.
[22] 廖声熙,崔 凯,张 鹏,等.翠柏木材管胞形态和结晶度的株内变异 [J].西北林学院学报,2012,27(6):159-164.
Liao S X,Cui K,Zhang P,et al.Variation of wood tracheid character and crystallinity ofCalocedrusmacrolepis[J].Journal of Northwest Forestry University,2012,27(6):159-164.
[23] 杨淑敏,江泽慧,任海青,等.利用 X-射线衍射法测定竹材纤维素结晶度 [J].东北林业大学学报,2010,38(8):75-77.
Yang S M,Jiang Z H,Ren H Q,et al.Determination of cellulose crystallinity of bamboo culms with X-ray diffraction spectrum [J].Journal of Northeast Forestry University,2010,38(8):75-77.
