黄土丘陵区6种侧蒴藓类植物营养繁殖特征
2018-02-13赵允格高丽倩魏玉清郝正刚
孙 会,赵允格,高丽倩,魏玉清*,郝正刚
(1 北方民族大学 生物科学与工程学院,银川 750021;2 西北农林科技大学 黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨陵 712100)
苔藓是最低等的高等植物,是很多生态系统中的先锋物种,被誉为生态系统的拓荒者。其中,侧蒴藓类是苔藓植物门中形体较大、形态结构较复杂的一种类群[1]。侧蒴藓类以匍匐的形态密集分布,能够在基质上形成紧密交织状的覆盖层,具有强大的吸水能力,在水土保持和水源涵养中起着重要生态作用[2]。研究表明,中国贺兰山云杉林下侧蒴藓类饱和持水量为2 kg·hm-2;长白山高山冻原植被的苔藓层持水量为3~5 kg·m-2,在雨后可吸收3 mm左右的雨水,并通过缓慢向土壤中释放水分来维持湿润环境[3-4]。同时,发育良好的藓类植物能够减少雨滴对土壤的打击而减少土壤侵蚀[5-6],具有显著的水土保持功能。此外,苔藓植物在CO2固定、营养物质循环与贮存等方面也有重要的生态功能[7]。研究表明,在寒温带针叶林中藓类植物是主要的生产者,年净产量达200~900 g·m-2[8],而在温带森林中藓类植物的年固碳量可达200 g·m-2[9],中国长白山云冷杉暗针叶林中苔藓植物生物量达5~6 t·hm-2[10]。因此,苔藓植物及其生态功能一直是为人们所关注的重要课题,特别是近年来,随着人们对退化生态系统及其恢复的重视,很多学者就苔藓植物的多样性、生态适应性、生态功能及其人工培养等展开不少研究。
纵观已有研究,可见研究者多关注于苔藓植物的分布、分类、多样性[11-12]及其生物学特性和进化特征等[13-14]。而关于苔藓植物的应用研究,特别是其在水土保持及生态恢复方面的应用研究起步相对较晚,主要研究了人工培养对其生长发育及其水土保持功能的影响[15]。已有研究表明,经人工接种并赋予一定的抚育措施可加快苔藓植物的生长发育及其生态功能的恢复,在退化生态系统恢复方面具有巨大的潜力[16]。然而,目前已有研究主要针对形态较小的顶生蒴藓[16-18]。侧蒴藓类是苔藓植物门中广泛分布的种群,具有体型大、材料易于获取等优势,已有研究表明,侧蒴藓类具有很好的人工栽培潜力。如杜宝明等以大灰藓为对象,研究了不同光照对其培养的影响,结果表明60%的遮光有利于其盖度增加且盖度高达93%以上[19]。但是目前国内外研究对侧蒴藓类人工培养的关注较少,更缺乏不同藓种的对比研究,限制了侧蒴藓类植物的应用。
黄土丘陵区苔藓植物种类丰富,主要以温带性种类为主,其中青藓科(Brachytheciaceae)、丛藓科(Pottiaceae)、灰藓科(Hypnaceae)、真藓科(Bryaceae)、羽藓科(Thuidiaceae)、柳叶藓科(Amblystegiaceae)、紫萼藓科(Grimmiaceae)、金发藓科(Polytrichaceae)等所占比重较大[20,21]。笔者也通过调查发现黄土丘陵区厚角绢藓(Entodonconcinnus)、柳叶藓(Amblystegiumserpens)、青藓(Brachythecyumalbicans)、鳞叶藓(Taxiphyllumtaxirameums)、镰叶灰藓(Hypnumbambergeri)、荫地绢藓(Entodoncaliginosus)等广泛分布,是研究区的常见藓种。因此,本研究以黄土丘陵区常见6种侧蒴藓类植物(具体见1.1供试材料)为对象,通过室内培养,采用切茎撒播法,对比研究了6种侧蒴藓类植物营养繁殖及生长发育过程,以期为藓类植物在水土保持及生态恢复中的应用提供科学依据。
1 材料和方法
1.1 供试材料
研究采用典型黄土丘陵区陕西省安塞县纸坊沟流域的厚角绢藓、青藓和富县子午岭林区的荫地绢藓、柳叶藓、鳞叶藓、镰叶灰藓6种侧蒴藓类植物为实验材料,样品在第二作者的协助下并参考苔藓植物图鉴等书籍资料进行鉴定[22-27],其形态特征见表1。
1.2 样品采集与处理
1.2.1采样区概况供试侧蒴藓类植物主要采自位于典型黄土丘陵区的陕西省安塞县纸坊沟流域和富县子午岭林区。纸坊沟流域(E109°19′30″,N36°51′30″)属暖温带半干旱气候,年辐射总量为493 kJ·cm-2,年均气温8.8 ℃,≥ 0℃的积温 3 733.5 ℃;多年年均降水量为549 mm,降水年内分布不均,多暴雨,7~9月份的降水量占全年降水的60%左右,年蒸发量大于1 463 mm。该流域主要优势植物种为刺槐(Robiniapseudoacacia)、丁香(Syringaoblata)、狼牙刺(Sophoraviciifolia)、沙棘(Hippophaerhamnoides)、黄刺玫(Rosaxanthina)、铁杆蒿(Artemisiagmelinii)、茭蒿(Artemisiagiraldii)等。
富县子午岭林区(E109°08′~109°11′,N36°04′~36°05′)地貌类型属梁状黄土丘陵沟壑区,海拔高程920~1 683 m,相对高差100~150 m,沟谷密度4.5 km·km-2。年均气温9 ℃,多年平均降雨量577 mm,多集中在7、8、9三个月,占全年降雨的60%以上,属于暖温带半湿润气候。主要树种有山杨(Populusdavidiana)、白桦(Betulaplatyphylla)、辽东栎(Quercusliaotungensis)、榆树(Ulmuspumila)和杜梨(Pyrusbetulaefolia)等。林下草灌覆盖度达90%以上,主要有绣线菊(Spiraeasalicifolia)、胡枝子(Lespedezabuergeri)、栒子叶柳(Salixkarelinii)、黄刺玫、山桃(Prunusdavidiana)、沙棘(Hippophaerhamnoides)、铁杆蒿、白草(Pennisetumcentrasiaticum)、羽茅(Achnatherumsibiricum)、野菊(Dendranthemaindicum)等,林下枯枝落叶层厚度2~5 cm。阴坡及半阴坡的森林长势较阳坡好,梁峁顶部多旱生草灌植被,局部有散生的杜梨分布。
在安塞纸坊沟采样区,侧蒴藓类植物主要分布在沟坡的刺槐林下,盖度约为10%左右,以青藓科和绢藓科为主。在富县子午岭林区,侧蒴藓类植物主要分布在针叶林的云杉和油松林及阔叶林的辽东栎林下,以灰藓科、绢藓科、羽藓科、柳叶藓科等分布为主。
1.2.2样品采集供试藓样品采于2017年9月12~15日,其中厚角绢藓和青藓采自安塞县纸坊沟流域,荫地绢藓、柳叶藓、镰叶灰藓和鳞叶藓采自富县任家台子午岭林区。同一藓种采集时选取长势相近,生境相似的作为实验材料,采样后装入透气档案袋后统一装入黑色塑料袋(避光),样品于阴凉处风干带回实验室,在17 ℃环境中避光保存以备用。
1.3 研究方法
1.3.1接种及培养方法将采集到的6个藓种绿色部分用灭菌剪刀剪成4~5 mm长的节段作为接种材料。接种量70 g·m-2,采用切茎少量多次撒播方法进行接种,撒播后用平整的小木板轻压使其与基质充分接触。采用白色塑料盒(242 mm×168 mm×50 mm)为培养容器,以过0.25 mm河沙和采自陕北安塞的黄绵土按质量比1∶1混合,按1.35 g·m-3的容重填装,培养基厚度约4 cm。按田间持水量的80%控制培养基的含水量[36],待沙土完全被水浸湿开始接种。接种后,置于AGC~D003N型人工气候室(温度17 ℃,光照4 500~5 500 Lx,光周期昼12 h/夜12 h,空气湿度60%~70%进行培养,期间每天观察培养基含水量的变化并及时补水,确保培养基质含水量保持在田间持水量的80%。以不接种为对照,重复3次。
培养基所用黄绵土的理化属性:有效氮25.64 mg·kg-1、速效磷3.93 mg·kg-1、有机质0.66 g·kg-1、全氮24.50 g·kg-1、全磷46.12 g·kg-1、速效钾46 mg·kg-1、容重1.32 g·m-3、pH 8.76。
1.3.2观测指标与方法自培养之日起,每天观测新配子体的萌发情况,当新萌发的配子体长度1 mm,记为新配子体开始萌发时间。在观测配子体萌发的同时,用解剖针采取接种配子体和培养基表面可能萌发的原丝体用显微镜观察鉴定,一旦确定是原丝体,即认为其开始萌发并记录萌发时间。新配子体开始萌发后,每盘用大头针标记20个接种体,测量其新发枝数量和发枝长度,每7天观测记录1次,至萌发的新配子体变化趋于稳定时停止(本研究共观测7次)。用10 cm×10 cm样方观测藓盖度,每培养盘观测3个样方。

1.4 统计分析
用SPSS 19.0对各藓种的发枝数量、发枝长度和盖度以及新配子体和原丝体的萌发时间进行正态分布检验(Kolmogorov-Smirnov test),并采用ANOVA进行方差分析(P<0.05)。用Sigmaplot.v10.0作图。
2 结果与分析
2.1 不同藓种新配子体和原丝体萌发时间
6种供试藓接种材料的节段在接种1 d后均可充分吸水进入复活状态。复活后的接种材料均在叶腋部位萌发产生新配子体,但不同藓种新配子体开始萌发的时间差异显著(表2),表现为,青藓最易萌发新配子体,用时7 d,其次为厚角绢藓,用时9 d,镰叶灰藓最慢,用时13 d。
各藓种均于接种材料周边的基质表面部位衍生出原丝体,且不同藓种原丝体萌发时间差异显著,而对照未发现侧蒴藓类植物的生长发育。6个藓种中,鳞叶藓最易萌发原丝体,用时14 d,其次为荫地绢藓, 柳叶藓最慢,用时22 d。原丝体萌发后,由原丝体产生配子体托,由配子体托发育成新的配子体,这个过程需要4~5 d的时间。发育形成的新配子体的周边可继续生长原丝体,如此反复可产生大量的原丝体和新配子体。另外,供试6种藓均表现为新配子体萌发时间早于原丝体萌发时间,且开始萌发的新配子体都开始于配子体接种材料,后由原丝体萌发新配子体。6种侧蒴藓类植物新配子体萌发时间平均先于原丝体萌发的时间为6.5 d。其中相差最大的是柳叶藓,为12 d,相差最小的是镰叶灰藓,为3 d。6种侧蒴藓类植物新配子体萌发的快慢顺序依次为:青藓>厚角绢藓>荫地绢藓=柳叶藓>鳞叶藓>镰叶灰藓。其中,萌发时间最早的青藓7 d,与镰叶灰藓和鳞叶藓差异显著(P<0.05)。萌发时间最晚的镰叶灰藓用时13 d。而原丝体的萌发时间快慢顺序依次为:鳞叶藓>荫地绢藓>镰叶灰藓>青藓=厚角绢藓>柳叶藓。其中,萌发时间最早的是鳞叶藓,为14 d,与其他藓种存在显著性差异(P<0.05),萌发最晚的是柳叶藓22 d,与其他藓种差异显著。
2.2 不同藓种新配子体发枝长度
培养条件下,6种侧蒴藓类植物发枝长度变化如图1所示。可见,从开始萌发后第7天到培养结束的整个过程中,6个藓种的发枝长度均呈先增加后趋于平稳的趋势。其中,在第7天到21天是6个藓种发枝长度快速增加期,平均增长速率1.40 mm·d-1,21 d后趋于平稳。在前期快速增加过程中,增加速度最快的是荫地绢藓,增加值为4.74 mm,其次为青藓,增加值为4.33 mm,镰叶灰藓增加最慢,增加值仅为2.76 mm。待到培养结束即第49天时,发枝长度最长的是青藓,为11.94 mm,其次是荫地绢藓,为10.20 mm,最短的是镰叶灰藓,为8.17 mm。从整个观测过程来看,青藓的发枝长度具有明显的优势。荫地绢藓的发枝长度变化最大,由开始到最后的变化值为5.35 mm,其次为青藓,变化值为4.73 mm,变化最小的是厚角绢藓,变化值仅为2.25 mm。厚角绢藓的发枝长度在开始萌发的1周内快速接近最大值(9.90 mm),在接下来7周的培养过程中,发枝长度的变化仅为2.25 mm。而镰叶灰藓在整个观测过程中的发枝长度变化最小。

图1 6种侧蒴藓类植物发枝长度变化Fig.1 Variation of new shoot length of the six pleurocarpous mosses
2.3 不同藓种发枝数量
人工培养条件下黄土丘陵区常见6种藓发枝数量的变化如图2所示。从图中可以看出,从开始观测到观测周期结束,6种藓发枝数量均有不同程度增加,主要表现为前期的增加到后期趋于稳定。青藓在整个培养过程中的发枝数量最多,平均为4.97条,具有明显的优势,由开始萌发的第7天到第21天缓慢增加,增加速率为0.029条·d-1,第21天到28天是快速增加期增加速率为0.129条·d-1,之后趋于稳定。其次为厚角绢藓,但优势不明显,从开始萌发的第7天到14天有一个快速增加期,增加速率为0.04条·d-1,之后趋于稳定。镰叶灰藓的发枝数量最少,为2.05条,且在整个培养过程中变化幅度最小。到培养结束,发枝数量由多到少的顺序依次为青藓4.97条>厚角绢藓2.68条>柳叶藓2.57条>鳞叶藓2.52条>荫地绢藓2.28条>镰叶灰藓2.05条。
2.4 培养期不同藓种盖度变化
培养条件下黄土丘陵区常见6种侧蒴藓类植物盖度变化见图3,由图可见在培养周期结束后,藓盖度由大到小的顺序依次为青藓48.89% > 柳叶藓39.56% > 镰叶灰藓30.22% > 鳞叶藓26.67% > 厚角绢藓24.00% > 荫地绢藓22.26%。6个藓种在开始萌发的第7天到21天以接近0.67%的相似增长速率增长,第21天到第49天培养结束的过程中增长速率有所不同,其中青藓的增长速率显著加快,最快的增长速率出现在第21天到28天,为2.16%·d-1,第21天到42天是快速上升期,上升速率为1.54%·d-1,之后趋于稳定。柳叶藓在培养过程中一直缓慢增加,平均增长速率为0.94%·d-1,除此之外其余藓种至培养周期结束均趋于稳定。在培养期盖度增长速率最低的是荫地绢藓,平均增长速率为0.40%·d-1,其增长速率在整个培养过程中相对稳定。

图2 6种侧蒴藓类植物发枝数量Fig.2 Variation of new shoot number of the six pleurocarpous mosses
2.5 不同藓种生长速率的变化
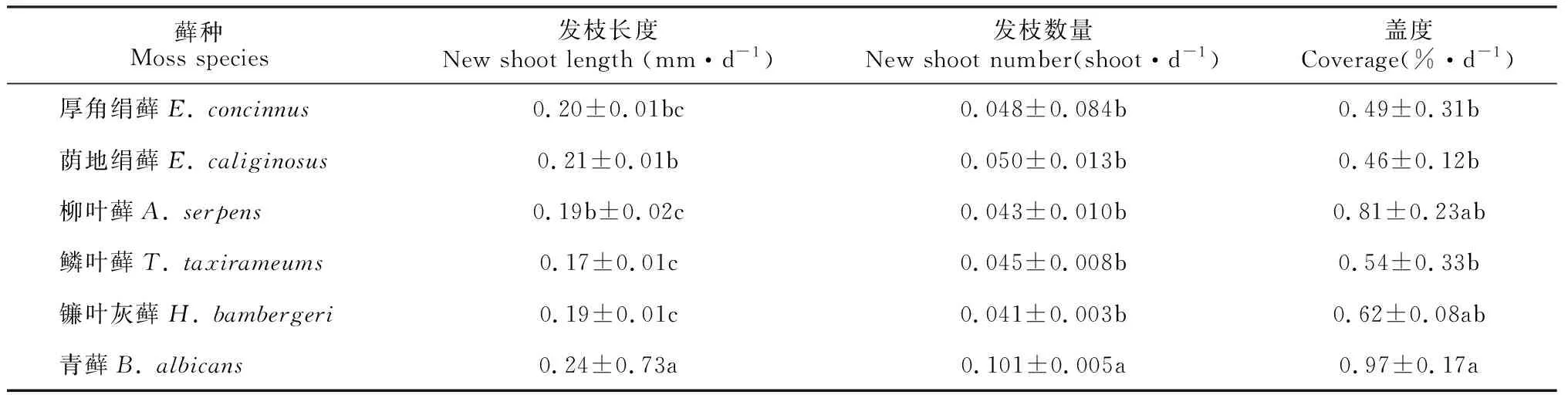
培养条件下黄土丘陵区6种侧蒴藓类植物的生长速率见表3,其中发枝长度、发枝数量和盖度的平均速率最大的均为青藓,分别为0.24 mm·d-1、0.101条·d-1、0.97%·d-1,且均与其他藓种差异显著(P<0.05)。荫地绢藓发枝长度的增加速率仅次于青藓,与鳞叶藓和镰叶灰藓差异显著(P<0.05)。发枝长度的平均变化速率最小的是鳞叶藓,为0.17 mm·d-1,与荫地绢藓也差异显著(P<0.05)。从发枝数量平均变化速率可知,镰叶灰藓的变化速率最小,仅有该物种与青藓存在显著差异(P<0.05),而其余藓种与青藓差异不显著(P>0.05)。盖度平均变化速率仅次于青藓的是柳叶藓,但与青藓差异不显著(P>0.05),最小的是荫地绢藓,与青藓存在显著差异(P>0.05)。

图3 6种侧蒴藓类植物盖度的变化Fig.3 Variation of coverage of the six pleurocarpous mosses
藓种 Moss species发枝长度New shoot length (mm·d-1)发枝数量New shoot number(shoot·d-1)盖度Coverage(%·d-1)厚角绢藓E. concinnus0.20±0.01bc0.048±0.084b0.49±0.31b荫地绢藓E. caliginosus0.21±0.01b0.050±0.013b0.46±0.12b柳叶藓A. serpens0.19b±0.02c0.043±0.010b0.81±0.23ab鳞叶藓T. taxirameums0.17±0.01c0.045±0.008b0.54±0.33b镰叶灰藓H. bambergeri0.19±0.01c0.041±0.003b0.62±0.08ab青藓B. albicans0.24±0.73a0.101±0.005a0.97±0.17a
3 讨 论
侧蒴藓类体型较大,结构较为复杂[1],且具有水源涵养、水土保持等多种生态功能[2],并在环境监测、医药、园林绿化、农业等方面也有着重要作用[26],因此被人们所重视。从开发利用的角度看,筛选藓类材料并研究其繁殖能力和方法将是今后研究的重点[27]。
苔藓植物的配子体是由原丝体、假根、茎、枝、叶5部分组成。苔藓植物的繁殖主要依靠配子体进行,并且形式多样[28]。本研究通过对黄土丘陵区常见的6种侧蒴藓类植物采用配子体繁殖的方式进行繁殖能力的对比,发现在培养过程中,6种侧蒴藓类植物均有新生配子体和原丝体的萌发,均表现出新生配子体先于原丝体出现。这可能是因为接种材料的配子体茎叶碎片具有极强的繁殖能力,茎的叶腋细胞经分裂、分化先产生小枝株,之后由新生小植株经细胞分裂产生原丝体原基,由原基产生原丝体的先后顺序决定的[29]。到培养结束的第49天6种侧蒴藓类植物的发枝长度、发枝数量和藓盖度均有明显变化,其中发枝长度最长的可达11.94 mm(青藓),最短的也可达8.83 mm(鳞叶藓);发枝数量亦有明显的变化,其中平均最大发枝数量为青藓的4.97条,最小发枝数量为镰叶灰藓(2.05条)。盖度的变化也很明显,最大盖度为青藓的48.89%,最小盖度为荫地绢藓的22.67% ,说明6种侧蒴藓类植物均可通过人工培养的方式进行繁殖,且具有很好的效果,具备应用潜力。
已有学者就该区的顶生蒴藓进行了不少人工培养研究,如杨永胜等研究了陕北安塞县的土生对齿藓(Didymodonvinealis)的人工培养,结果表明,在与本研究相同的培养条件下以接种量为250 g·m-2,经过60 d的培养藓结皮的盖度仍不到30%,培养50 d藓结皮的盖度不到25%。盖度的平均增长速率不到0.5%·d-1 [30]。陈彦芹以该区短叶扭口藓(Barbulaunguiculata)为对象,在与本研究相同条件下进行人工培养,结果表明,经过30 d的培养,藓盖度为9.8%、藓株高0.8 mm,盖度的平均增长速率为0.33%·d-1 [16]。本研究中6种侧蒴藓类植物在接种量为70 g·m-2经过49 d的人工培养,盖度最高可达48.89%(青藓),增长速率最高为0.97%·d-1,最小盖度为22.6%(荫地绢藓),增长率为0.46%。植株高度最高为11.94 mm(青藓),最低为8.17 mm(镰叶灰藓)。可见,侧蒴藓类的人工培养潜力相对于顶生蒴藓具有明显的优势。
另一方面,本研究结果表明,培养条件下,不同侧蒴藓类物种的营养繁殖存在明显种间差异。新生配子体的萌发和原丝体出现的时间,在培养过程中发枝长度、发枝数量和苔藓盖度的变化趋势有所不同,其中青藓的培养优势最为明显;该物种在培养过程中发枝长度、发枝数量和盖度变化均表现出明显的优势,生长速率(发枝长度、发枝数量、盖度的增长速率)高于其他藓种,且差异显著。培养周期结束青藓的发枝长度、发枝数量与盖度也均为最高。而镰叶灰藓在发枝长度和发枝数量增加量最小,盖度增加量最小的是荫地绢藓,其余藓种在3个观测指标中趋于两者之间。从生长速率来看,鳞叶藓和镰叶灰藓分别在发枝长度和发枝数量的生长速率变化最慢,在盖度的生长速率上变化最慢的是荫地绢藓。6种侧蒴藓类植物繁殖能力存在差异,可能与来自不同生境的不同藓种有关,其中厚角绢藓和青藓采自陕西北部的安塞县纸坊沟流域,该地区采样点林下郁闭度为15%~20%、年均气温8.8 ℃、年均降雨量为549 mm,而镰叶灰藓、柳叶藓、镰叶灰藓、鳞叶藓采自陕西北部的富县子午岭林区,该地区采样点林下郁闭度为30%~70%、年均气温9 ℃、年均降雨量576 mm。采自不同生境的不同藓种对光、温、水的需求有所不同[2, 31-35],这可能是造成不同藓种在相同培养条件下结果存在差异的原因。此外,不同藓种的形态特征和生理特性与其生态环境有关,其营养繁殖特征的差异可能还与其生物学特性有关[36-39],这可能也是造成其繁殖能力差异的原因之一。
4 结 论
对黄土丘陵区6种常见侧蒴藓类植物进行人工培养,通过对新生配子体和原丝体的萌发时间、发枝数量、发枝长度和盖度变化进行观测,得到以下结果:
(1)黄土丘陵区6种侧蒴藓类植物在人工培养条件下原丝体和新配子体均可萌发,且新配子体的萌发时间早于原丝体,新配子体发枝长度、发枝数量、藓盖度从萌发开始到培养周期结束均有明显的增加。6种藓均可进行人工培养,且与该地区顶生蒴藓的人工培养相比均表现出较好的培养潜力。
(2)黄土丘陵区常见6种侧蒴藓类植物的繁殖能力具有种间差异,其中青藓在发枝长度、发枝数量、藓盖度的生长速率增加最为明显,且与其余藓种存在显著性差异。青藓的营养繁殖能力,具有显著的优势。研究结果可为藓类植物在水土保持、水源涵养及修复退化的生态系统等方面应用提供科学依据。
