染料木素的作用机制及在畜牧生产中的应用研究
2018-01-24甘麦邻曾仰双杨大洪张顺华
甘麦邻,杨 琼,李 强,曾仰双,杨大洪,张顺华★,朱 砺★
(1.四川农业大学动物科技学院,四川 成都 611130;2.成都农业科技职业学院,四川 成都 611130;3.四川省畜牧总站,四川 成都 610041;4.四川省雅安市名山区农业局,四川 雅安 625100)
植物雌激素是存在于植物中的具有雌激素作用的一类天然化合物,具有与雌激素类似的分子结构,能与雌激素受体(ER)竞争性结合(图1-Ⅰ)。雌激素在各种动物中的生理作用具有高度同源性,不同种动物体内的雌激素结构也高度类似,在动物机体中,植物雌激素因动物体内雌激素浓度高低、受体数目、结合程度以及用药的剂量,表现出对内源雌激素活性的促进或拮抗作用[1]。染料木素是一种主要的植物雌激素,又称为染料木黄酮、5,7,4′-三羟基异黄酮,是异黄酮类化合物中生物活性最高的一种,广泛存在于大豆、苜蓿、三叶草等豆科植物的茎叶和籽实中。
研究发现染料木素具有抗癌、防治骨质疏松症、保护心血管系统健康、提高机体免疫力和抗抑郁等作用[2-4]。由于其丰富的生物学效应和广泛存在引起了药物学家和营养学家的关注,目前广泛应用于制药、保健等领域[5-6]。已经有大量文献报道了关于植物雌激素的使用,但其中绝大多数研究中使用的是粗提物或者混合物,针对单一性有效成分的研究只占少数[7]。长期以来由于染料木素的提纯工艺存在耗时长、纯化难度大、价格昂贵等问题未能被大量应用,近年来随着提纯方法和提纯工艺的改进,获得高纯度的原料越来越容易,为其在畜牧生产中广泛应用打下了基础[8]。畜牧生产过程中生长性能、繁殖性能和抗病性能是生产者们关注的重点,直接影响着畜牧生产的收益。因此本文对近年来染料木素的作用机制及其在畜牧生产中的应用进行综述,以期为染料木素在畜牧生产中的应用提供较为准确的参考,为更好地利用我国的植物资源提供借鉴。
1 染料木素的作用机制
1.1与雌激素受体结合发挥雌激素样或抗雌激素作用染料木素与雌激素的结构相似,因此可与雌激素受体结合进而发挥作用。20世纪50年代首次报道了染料木素的雌激素样作用[9]。动物体内的雌激素受体,分为α型(ERα)和β型(ERβ),研究发现染料木素与ERβ的亲和性高于ERα,与雌激素相比染料木素与雌激素受体的亲和力更低,但可以双向调节,通常在低浓度时发挥雌激素样作用,而高浓度时发挥抗雌激素作用[10]。同时染料木素还可以增加受试动物体内雌激素受体不同亚型的表达,进而影响机体代谢[11]。
1.2抗氧化作用由于染料木素具有多羟基酚的结构(见图1-Ⅰ),因此具有一定的抗氧化活性。研究发现一定剂量的染料木素可以增加乳腺细胞内总超氧化物歧化酶(T-SOD)的含量[12],活体试验也证实了饲喂染料木素可以增加动物机体抗氧化能力[13]。同时染料木素可以减弱卵巢切除小鼠的氧化应激、脂质过氧化作用,对雌激素缺乏导致的神经退行性病变起到保护作用[14]。染料木素还可以抑制脑缺血再灌注损伤引起的氧化应激,从而起到脑保护作用[15]。

图1 染料木素(genistein)和雌二醇(Estradiol)分子结构(Ⅰ)及染料木素与EGFR-TK结合位点的微环境(Ⅱ,引自[17])Fig.1 The molecular structure of genistein and estradiol(Ⅰ),the binding site of genistein to EGFR-TK(Ⅱ,[17])
1.3天然的酪氨酸激酶抑制剂染料木素是酪氨酸激酶的高度特异的非竞争性抑制剂,可有效地抑制表皮生长因子受体(EGFR)等信号传导途径[16]。模拟对接试验发现染料木素可结合在EGFR-TK的活性腔中,通过干扰TK催化活性结构中Lys 721/Glu 738位点对非竞争性结合发挥抑制作用(见图1-Ⅱ)[17]。由于PTK活性与表皮生长因子(EGF)、胰岛素和胰岛素样生长因子Ⅰ(IGF-1)等受体相关联,因此染料木素的抑制作用可能会引起细胞中一些关键PTK的激活抑制,从而影响机体代谢[18-19]。
1.4拓扑异构酶抑制剂染料木素还可以通过抑制拓扑异构酶的活性发挥作用。拓扑异构酶通过催化DNA链的断裂和结合,从而调控DNA的拓扑状态,在基因复制、转录、重组、修复和染色体重塑过程中发挥着重要作用[20]。研究发现染料木素通过抑制拓扑异构酶Ⅱ的表达促进Hela细胞凋亡[21],还可以通过抑制DNA拓扑异构酶Ⅱ的活性来减少破骨细胞形成[22]。
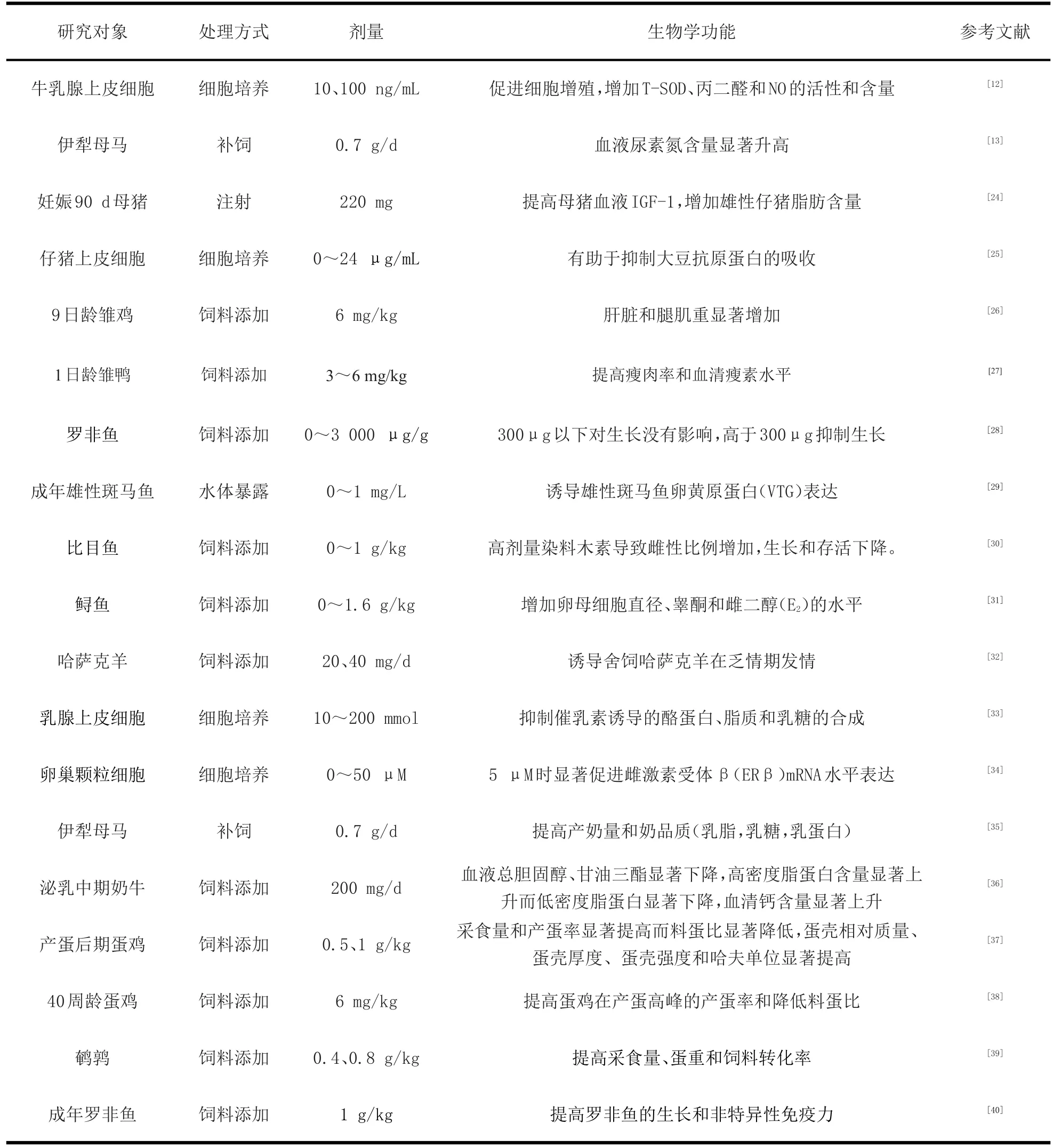
表1 染料木素在畜牧生产应用中发挥的生物学功能Table 1 The biological function of genistein in livestock production and application
除了上述被广泛研究的作用途径,染料木素还可以非竞争性结合11β-羟基类固醇脱氢酶1(11β-HSD1)、己糖-6-磷酸脱氢酶(H6PD)或通过抑制细胞周期,诱发细胞凋亡等途径发挥生物学功能[23]。在畜牧生产中可依据畜禽的生理状态,通过控制染料木素的剂量激发染料木素的不同作用途径来达成促生长、调控繁殖、预防和治疗疾病等目的(见表1)。
2 染料木素对动物生长发育的影响
动物生长发育过程中伴随着骨骼、肌肉、神经和脂肪等组织质和量的变化。动物体内雌激素水平不仅影响机体卵巢、子宫、乳腺等生殖系统器官的发育和功能,还可以通过与雌激素受体结合对下丘脑-垂体-性腺轴所涉及的器官和组织产生影响[41]。由于雌激素受体广泛存在于动物机体的各种组织和器官中,因此存在雌激素受体的部位都可能是染料木素的潜在靶点,并可能发挥相应的生物学作用。
C.Farmer等[24]对妊娠90 d的母猪每日2次注射220 mg染料木素,待其分娩后进行采血和选取1日龄小公猪进行屠宰测定,发现处理组母猪血液IGF-1水平及初生仔猪脂肪含量显著提高,同时发现处理组的断奶前死亡率降低为对照组的一半。张诗尧等[25]利用仔猪上皮细胞进行β-伴大豆球蛋白(β-Conglycinin)及其酶解产物吸收规律的研究,发现添加一定浓度的染料木素有利于仔猪肠道对大豆抗原蛋白的吸收。针对禽类的研究,耿志荣[26]发现给9日龄雏鸡饲料中添加6 mg/kg染料木素,饲喂2周后与对照组相比可以提高日增重8.21%,生长速率增加3.50%(第1周)和3.97%(第2周),同时料重比下降6.50%。2周后染料木素组肝脏重量显著增加,腿重增加11.24%。孟婷等[27]在1日龄雏鸭日粮中添加染料木素饲喂7周,发现3 mg/kg和6 mg/kg对雌性樱桃谷肉鸭均无显著影响,均能显著降低雄性肉鸭腹脂率和血清低密度脂蛋白和总胆固醇,6 mg/kg时还可显著提高雄性肉鸭瘦肉率和血清瘦素水平。陈栋[28]给罗非鱼(10.47 g)饲料中添加0、30、300、3 000 μg/g的染料木素,发现在300 μg/g浓度以下时对罗非鱼生长没有影响,而高浓度染料木素会抑制生长。同时高浓度染料木素会降低罗非鱼主要消化酶的活性并能干扰罗非鱼的蛋白和糖代谢,降低罗非鱼对营养物质的吸收和转化。另一方面高浓度染料木素抑制了GH/IGF-1生长轴线上游脑ghrelin、NPY R1的表达,进而降低GH合成与分泌;通过抑制肝脏GHR2、IGF-1R表达和促进IGFBP3表达,造成IGF-1合成的降低和游离IGF-1的减少,从而抑制了IGF-1介导的GH促生长作用。
在模式动物上研究,M.Velders等[11]发现,为去卵巢大鼠补充染料木素增加了大鼠骨骼肌中ERβ和Myh在mRNA水平和蛋白水平的表达量,并证明异黄酮可通过ERβ调节Myh的表达进而增强骨骼肌的收缩力。此前的研究结果也发现小鼠哺乳期饲喂含染料木素25 mg/kg的饲料可以促进后代仔鼠体重的增加,饲喂浓度达到50 mg/kg时则会影响仔鼠器官的发育,但当浓度提高到100 mg/kg以上时仔鼠生长就表现出抑制作用,同时严重影响仔鼠肝脏和性腺等敏感组织的生长发育[42]。
以上研究表明染料木素确实能够对动物生长产生影响,但同时这种影响表现出性别差异和剂量差异,在实际应用过程中仍需根据畜种及特定的生理状态进行前期投喂的摸索。
3 对动物繁殖生理的影响
3.1对种畜繁殖能力的影响本世纪初R.Ingham等[43]就报道了造纸厂排放废水中的染料木素及其他植物雌激素可能通过影响鱼类类固醇激素的水平,造成了工厂附近鱼类的生殖能力的下降。陈洁[29]将成年雄性斑马鱼暴露在不同浓度的染料木素中,发现染料木素可以诱导雄性斑马鱼卵黄原蛋白(VTG)表达,同时一部分剂量组睾酮含量显著下降,雌激素含量显著上升,ERβ mRNA的表达量也显著升高,并呈现出剂量依赖性,其结果揭示染料木素诱导雄性斑马鱼VTG的表达可能方式是通过雌激素β受体(ERβ)途径和直接提高雄鱼性腺内雌二醇的水平发挥作用。M.Dimaggio等[30]的研究发现日粮中的染料木素可以影响比目鱼的性别、生长发育和存活率,添加0、100、1 000 mg/kg染料木素时,发现高剂量组雌鱼更多,同时高剂量染料木素会导致比目鱼生长和存活下降。C.Bennetau等[44]以用作对照的鱼粉为基础的饮食制备500 ppm和1 000 ppm的富含染料木素的饲料,发现在500 ppm浓度时饲养鱼类的生殖能力就可能受损。A Jourdehi等[31]利用鲟鱼作为试验动物,在其日粮中分别添加0.2、0.4、0.8、1.6 g/kg染料木素,发现染料木素的添加可以增加雌鱼卵母细胞的直径,并提高体内睾酮和雌二醇(E2)的水平,同时还可增加体内钙、磷和胆固醇含量。
Li等[45]研究发现500 ppm染料木素处理可以增加小鼠的乳腺组织质量和提前雌鼠的阴道开口时间。此前对小鼠和哈萨克羊的试验中也发现染料木素能影响动物的发情及生殖器官变化。在雌性未性成熟的小鼠日粮中添加一定量的染料木素可以增加雌性小鼠性器官的湿重,对于性器官重量的增加可能主要是起雌激素样作用的结果[32]。在乏情期饲喂一定剂量的染料木素可以诱导哈萨克羊在乏情期发情,其作用机制可能通过雌激素样作用以及改善乏情期绵羊的体况诱导发情[46]。利用体外培养的颗粒细胞研究染料木素的雌激素效应作用,研究者发现5 μM浓度的染料木素可以显著提高ERβ的mRNA水平[33]。
3.2对繁殖相关经济性状的影响产奶和产蛋是分别是哺乳动物和禽类重要的繁殖性状,同时也是乳畜和蛋禽最主要的经济性状。早在1992年Fan等发现染料木素可以以剂量依赖的方式抑制催乳素(Prolactin,PRL)诱导的乳腺上皮细胞中酪蛋白、脂质和乳糖的合成[34]。
臧长江等[13]对泌乳期的伊犁母马补饲0.7 g染料木素,发现其血液尿素氮含量显著升高,并提示泌乳期母马饲喂染料木素对马匹免疫力提高和机体抗氧化能力有一定促进作用。郭云等[35]选用泌乳期为第3个月的伊犁母马每日添加0.7 g染料木素,饲喂42 d时乳蛋白产量提高29.61%,乳脂产量提高50.25%,饲喂56 d时乳糖产量提高20.29%,饲喂70 d时产奶量提高20.00%,以上结果提示泌乳期补饲染料木素可以提高伊犁母马的产奶量和奶品质。刘春龙等[36]给与泌乳中期奶牛每天200 mg染料木素,发现染料木素添加组血液中总胆固醇、甘油三酯含量显著下降,高密度脂蛋白含量显著上升而低密度脂蛋白显著下降,血清钙含量显著上升,并在一定程度上降低了奶牛血液的黏稠度。通过体外培养试验,研究人员发现添加10、100 ng/mL染料木素可以显著促进牛乳腺上皮细胞增殖,可显著增加总超氧化物歧化酶(T-SOD)、丙二醛(MDA)和一氧化氮(NO)的活性和含量[12]。
对蛋禽的研究也发现添加一定剂量的染料木素可以提高蛋禽的生产性能。陈常秀等[37]发现在蛋鸡产蛋后期饲料中添加染料木素可提高蛋鸡产蛋性能和鸡蛋品质及抗氧化能力,研究人员在蛋鸡490~587日龄时以500 mg/kg和1 000 mg/kg的染料木素添加量进行饲喂,发现1 000 mg/kg组的采食量和产蛋率显著提高而料蛋比显著降低,蛋壳相对质量、蛋壳厚度、蛋壳强度和哈夫单位也显著提高,同时蛋中染料木素的含量也显著增加,蛋重和蛋比重则无明显变化。徐国银[38]在40周龄的蛋鸡饲料中添加6 mg/kg的染料木素,持续饲喂到46周,发现染料木素可以提高蛋鸡在产蛋高峰的产蛋率和降低料蛋比,对蛋重则无明显影响,进一步研究发现染料木素还可以显著降低血清总胆固醇和甘油三酯。针对鹌鹑产蛋的研究中发现在日粮中添加染料木素有利于鹌鹑产蛋性能的提升,添加800mg/kg染料木素时可以显著提高鹌鹑采食量、蛋重和饲料转化率等指标,而不影响蛋黄黄豆苷原,维生素A和维生素E水平[39]。
姜晶晶[47]对青春期雌性大鼠进行6 mg/100 g体重的剂量灌胃,发现灌胃染料木素后大鼠乳腺相对体重百分比较对照组提高35.41%,乳腺组织RNA和DNA含量分别提高27.24%和4.27%,而RNA/DNA升高22.07%,提示染料木素可以促进大鼠乳腺的发育。血清17β雌二醇含量升高26.68%,孕酮水平提高130.54%。同时在日粮中进行添加染料木素,发现灌胃和日粮添加效果基本一致。
4 对动物抗病及免疫功能的影响
S.Chakraborty等[40]给成年罗非鱼(39.55 g)饲料中添加1 g/kg的染料木素饲喂35 d,发现饲喂染料木素之后可以提高罗非鱼的生长和非特异性免疫力。Wan等[48]研究发现染料木黄酮能降低日本血吸虫卵诱导的肝肉芽肿和纤维化,至少部分是由于NF-κB信号传导下降,随后降低了MCP1、TNFα和IL10的表达。这意味着染料木素可能是抗血吸虫病的潜在天然药物。陆扬[49]发现与VOA致敏小鼠相比口服染料木素可以减小VOA致敏小鼠的血清IgG1水平和mMCP-7肥大细胞的数量,同时口服染料木素后致敏小鼠的肠道微生物菌群的丰富性和优势菌群没有发生显著变化,提示染料木素可以缓解食物过敏的进程。王海涛等[50]发现染料木素可以通过破坏细茵细胞壁及细胞膜的完整性、抑制细茵的呼吸代谢和抑制蛋白质的合成等抑制金黄色葡萄球菌的生长。
5 总结与展望
已有的文献表明染料木素在畜牧生产应用中可发挥双向调节作用,通常低浓度表现为对动物产奶、产蛋、生长及发育的促进和对细胞增殖及抗氧化能力的提升,高浓度则表现出对动物生长及繁殖性能的损伤和对细胞增值的抑制及促进凋亡。除此之外染料木素的靶向位点雌激素受体类型、受体酪氨酸激酶的分布和动物生理状态的差异也会影响染料木素的作用效果。由于其广泛的生理学作用,染料木素可应用在畜牧生产领域的多个方面,同时染料木素对动物机体的调控网络极其复杂,其实际生产应用中的作用机理和可能产生的不良影响仍需进一步探索。
[1]Wang H,Murphy PA.Isoflavone Content in Commercial Soybean Foods[J].Journal of Agricultural& Food Chemistry,1994,42(8):1666-1673.
[2]Sureda A,Silva AS,Sánchez-Machado D,et al.Hypotensive effects of genistein:From chemistry to medicine[J].Chemico-biological interactions,2017,268:37-46.
[3]Wang YX,Tian K,He CC,et al.Genistein inhibits hypoxia,ischemic-induced death,and apoptosis in PC12 cells[J].Environmental Toxicology&Pharmacology,2017,50:227-233.
[4]Liu J,Yu X,Zhong S,et al.Hepatic and renal metabolism of genistein:An individualbased model to predict glucuronidation behavior of genistein in different organs[J].Journal of Pharmaceutical&Biomedical Analysis,2017,139:252-262.
[5]Odle B,Dennison N,Al-Nakkash L,et al.Genistein treatment improves fracture resistance in obese diabetic mice[J].Bmc Endocrine Disorders,2017,17(1):1-8.
[6]de Oliveira MR.Evidence for genistein as a mitochondriotropic molecule[J].Mitochondrion,2016,29:35-44.
[7]石建,马静,石苗茜.基于文献计量的染料木素最新研究现状分析[J].现代生物医学进展,2010,10(24):4768-4773.
[8]蔡锦源,陈玲,朱炽雄,等.染料木素的制备方法研究进展[J].粮食科技与经济,2015,(4):69-72.
[9]Cheng E,Story CD,Yoder L,et al.Estrogenic activity of isoflavone derivatives extracted and prepared from soybean oil meal[J].Science,1953,118(3058):164-165.
[10]Liu X,Xue X,Wang H,et al.Genistein modulates MMP-26 and estrogen receptor expression in endometrial cancer cells[J].2017,3(4):242-247.
[11]Velders M,Solzbacher M,Schleipen B,et al.Estradiol and genistein antagonize the ovariectomy effects on skeletal muscle myosin heavy chain expression via ER-beta mediated pathways[J].Journal of Steroid Biochemistry&Molecular Biology,2010,120(1):53-59.
[12]刘春龙,李忠秋,张帆,等.大豆黄酮和染料木素对体外培养奶牛乳腺上皮细胞增殖及抗氧化水平的影响[J].畜牧兽医学报,2008,39(11):1517-1522.
[13]臧长江,赵芳,李晓斌,等.补饲植物雌激素对泌乳期伊犁母马血液生化指标及抗氧化力的影响[J].中国畜牧兽医,2015,42(12):3226-3230.
[14]Huang YH,Zhang QH.Genistein reduced the neural apoptosis in the brain of ovariectomised rats by modulating mitochondrial oxidative stress[J].Br J Nutr.,2010,104(9):1297-1303.
[15]Ma Y,Sullivan JC,Schreihofer DA.Dietary genistein and equol(4',7 isoflavandiol)reduce oxidative stress and protect rats against focal cerebral ischemia[J].Am J Physiol Regul Integr Comp Physiol.,2010,299(3):R871.
[16]Elmarakby AA,Ibrahim AS,Faulkner J,et al.Tyrosine kinase inhibitor,genistein,reduces renal inflammation and injury in streptozotocin-induced diabetic mice[J].Vascular Pharmacology,2011,55(5-6):149-156.
[17]袁江兰,刘晖,康旭,等.表皮生长因子受体酪氨酸激酶结构域与其抑制剂染料木素的模拟对接[J].生物工程学报,2008,24(10):1813-1817.
[18]陈璐瑶,杨洋,安输,等.G蛋白偶联受体与酪氨酸激酶受体之间的信号交流在肿瘤治疗中的作用[J].中国药理学通报,2017,33(4):454-460.
[19]Xiao GS,Zhang YH,Wu W,et al.Genistein and tyrphostin AG556 decrease ultra-rapidly activating delayed rectifier K(+)current of human atria by inhibiting EGF receptor tyrosine kinase[J].British Journal of Pharmacology,2017,174(6):454-467.
[20]秦尚尧,袁一旻,胡昕,等.拓扑异构酶Ⅱ的结构、功能及作用机制研究进展[J].生理学报,2016,(1):98-106.
[21]Zhou N,Yan Y,Li W,et al.Genistein Inhibition of Topoisomerase IIα Expression Participated by Sp1 and Sp3 in HeLa Cell[J].International Journal of Molecular Sciences,2009,10(7):3255-3268.
[22]Zhang JM.Advances in studies on phytoestrogens in the prevention and treatment of postmenopausal osteoporosis[J].Drugs&Clinic.,2012,27(1):65-69.
[23]Tagawa N,Kubota S,Kobayashi Y,et al.Genistein inhibits glucocorticoid amplification in adipose tissue by suppression of 11β-hydroxysteroid dehydrogenase type 1[J].Steroids,2015,93:77-86.
[24]Farmer C,Robertson P,Xiao CW,et al.Exogenous genistein in late gestation:effects on fetal development and sow and piglet performance[J].Animal An International Journal of Animal Bioscience,2016,10(9):1-8.
[25]张诗尧.仔猪小肠上皮细胞对β-Conglycinin及其酶解产物吸收规律的研究[D].吉林:吉林农业大学,2016.
[26]耿志荣.大豆黄酮和染料木素的RP-HPLC方法的建立及其在禽类体内蛋内含量的研究[D].南京:南京农业大学,2003.
[27]孟婷,左伟勇,王健,等.染料木素对肉鸭屠宰性能及脂肪代谢的影响[J].中国畜牧兽医.2009,36(12):207-209.
[28]陈栋.金雀异黄素对尼罗罗非鱼(Oreochromis niloticus)生长的影响及其机制研究[D].北京:中国海洋大学,2013.
[29]陈洁.金雀异黄素对雄性斑马鱼VTG的诱导及其机理研究[D].北京:中国海洋大学,2010.
[30]Dimaggio MA,Kenter LW,Breton TS,et al.Effects of dietary genistein administration on growth,survival and sex determination in southern flounder,Paralichthys lethostigma[J].Aquaculture Research,2016,47(1):82-90.
[31]Jourdehi AY,Sudagar M,Bahmani M,et al.Comparative study of dietary soy phytoestrogens genistein and equol effects on growth parameters and ovarian development in farmed female beluga sturgeon,Huso hus o[J].Fish Physiology&Biochemistry,2014,40(1):117-128.
[32]甘麦邻,赵宗胜,黄琳璇,等.染料木素对青年母鼠生殖器官和体重的影响[J].当代畜牧,2014,(18):50-51.
[33]Nynca A,Nynca J,Wsowska B,et al.Effects of the phytoestrogen,genistein,and protein tyrosine kinase inhibitor-dependent mechanisms on steroidogenesis and estrogen receptor expression in porcine granulosa cells of medium follicles[J].Domestic Animal Endocrinology,2013,44(1):10-18.
[34]Fan G,Rillema JA.Effect of a tyrosine kinase inhibitor,genistein,on the actions of prolactin in cultured mouse mammary tissues[J].Molecular& Cellular Endocrinology,1992,83(1):51-56.
[35]郭云,赵芳,邓海峰,等.补喂植物雌激素对泌乳期伊犁母马产奶量和乳成分的影响[J].中国饲料,2016,(5):5-8.
[36]刘春龙,李忠秋,张帆,等.大豆黄酮和染料木素对奶牛血清生化及血液流变学指标的影响[J].中国兽医学报,2009,29(4):460-463.
[37]陈常秀,李永洙.染料木素对后期蛋鸡生产性能及蛋品质的影响[J].西北农林科技大学学报:自然科学版,2011,39(9):53-57.
[38]徐国银.日粮中添加异黄酮对肉鸡和蛋鸡生产性能的影响及其内分泌机制的研究[D].南京:南京农业大学,2007.
[39]Akdemir F,Sahin K.Genistein supplementation to the quail:effects on egg production and egg yolk genistein,daidzein,and lipid peroxidation levels[J].Poultry Science,2009,88(10):2125-2131.
[40]Chakraborty SB,Molnár T,Ardó L,et al.Oral Administration of Basella alba Leaf Methanol Extract and Genistein Enhances the Growth and Non-Specific Immune Responses of Oreochromis niloticus[J].Turkish Journal of Fisheries&Aquatic Sciences,2015,15(1):167-173.
[41]王虹,樊官伟,王少峡,等.雌激素受体介导的快速信号通路的研究进展[J].现代生物医学进展,2008,8(3):555-557.
[42]甘麦邻,姜茹斌,陈焱森,等.哺乳期母鼠饲喂染料木素对仔鼠早期生长发育的影响[J].四川农业大学学报,2017,35(1):75-80.
[43]Ingham RR,Gesualdi DA,Toth CR,et al.Effects of Genistein on Growth and Development of Aquatic Vertebrates[J].Bulletin of Environmental Contamination&Toxicology,2004,72(3):625-631.
[44]Bennetau-Pelissero C,Breton B,Bennetau B,et al.Effect of genistein enriched diet on the sex steroid endocrinology and the reproductive efficiency of the rainbow trout Oncorhynchus mykiss[J].Revue De Medecine Veterinaire,2002,153(7):513-516.
[45]Rong L,Zowalaty AEE,Chen W,et al.Segregated responses of mammary gland development and vaginal opening to prepubertal genistein exposure in Bscl2-/-female mice with lipodystrophy[J].Reproductive Toxicology,2014,54:76-83.
[46]甘麦邻,努尔力,阿不利孜,等.染料木素对舍饲哈萨克羊乏情期诱导发情的作用[J].畜牧与兽医,2016,48(2):61-64.
[47]姜晶晶.染料木素和亚麻籽木脂素对青春期雌性大鼠乳腺发育及相关性激素影响的研究[D].南京:南京农业大学,2008.
[48]Wan C,Jin F,Du Y,et al.Genistein improves schistosomiasis liver granuloma and fibrosis via dampening NF-kB signaling in mice[J].Parasitology Research,2017,116(4):1165-1174.
[49]陆扬.大豆苷元对断奶和食物过敏动物消化道的影响及TLRs在感染进程中的表达[D].南京:南京农业大学,2009.
[50]王海涛,石姗姗,李银霞,等.染料木素的抑菌活性及其机制的研究[J].营养学报,2008,30(4):403-406.
