应用东乡野生稻回交重组自交系群体分析糙米矿质含量QTL
2018-01-22胡标林黄得润肖叶青何强生万勇樊叶杨
胡标林 黄得润 肖叶青 何强生 万勇,* 樊叶杨,*
应用东乡野生稻回交重组自交系群体分析糙米矿质含量QTL
胡标林1,2黄得润1肖叶青2何强生3万勇2,*樊叶杨1,*
(1中国水稻研究所 国家水稻改良中心/水稻生物学国家重点实验室, 杭州 310006;2江西农业科学院 水稻研究所/国家水稻改良中心南昌分中心, 南昌 330200;3江西兴安种业有限公司, 江西上饶 334300;*通讯联系人, E-mail: wanyong025@163.com, fanyeyangcnrri@163.com)
强化粮食作物的必需矿物质有利于缓解人们矿质营养缺乏症。从协青早B//协青早B/东乡野生稻BC1F5群体中挑选到1个单株A58,与协青早B回交,构建了BC2F4:5群体。采用电感耦合等离子体原子发射光谱仪ICP-AES测定132个BC2F4:5株系的糙米Mg、Ca、Zn、Fe、Mn和Cu含量,应用Windows QTL Cartographer 2.5进行糙米矿质QTL分析。共检测到17个糙米矿质含量QTL,分别位于第1、4、6、8、9和11等6条染色体上,包括Mg含量1个、Ca含量4个、Zn含量4个、Fe含量2个、Mn含量2个和Cu含量4个。这些QTL解释表型变异的5.0%~47.2%,其中8个QTL的增效等位基因来自东乡野生稻。12个QTL聚集于5条染色体上的5个QTL簇,表明不同矿质营养元素涉及到共同遗传生理机制,可通过分子标记辅助选择方法将有利等位基因应用于稻米营养品质改良。
东乡野生稻;糙米;矿质含量;QTL定位
矿质元素在人体内具有很多细胞代谢和生理生化功能,参与人体内的多种酶结构和激活[1]。随着人们膳食结构和生活方式的变化,矿质营养失衡日渐凸显,已成为影响人类健康的重要因素。在人类流行的镁(Mg)、钙(Ca)、锌(Zn)、铁(Fe)、锰(Mn)和铜(Cu)等元素缺乏症中[2],以Zn和Fe缺乏最为普遍[3]。由于矿质元素不能在体内合成,过去主要依靠保健品和药物补充等方法补充人体缺乏矿质元素以缓解矿质元素缺乏症[4]。然而这些传统方法覆盖面小、费用高,由此主食被认为是一种经济有效的矿质元素补充途径。因此,通过生物强化方法提高粮食作物可食用部位中的矿质含量日益受到研究者们的关注。
水稻作为超过50%全球人群赖以生存的主要粮食之一,是矿质元素生物强化的重要对象[5]。研究表明不同基因型水稻的稻米矿质含量存在广泛的遗传变异,且遗传成分在决定稻米矿质含量中起主要作用[6],这为筛选和选育富含矿质元素的水稻品种提供了可能性。
水稻传统育种方法主要通过表型间接对基因型进行选择,需要丰富的育种经验和较长时间;同时如稻米矿质含量等特殊性状的选择受到很多因素限制,选择效率低下。QTL分析是解析稻米矿质含量等数量性状的有效方法,广泛应用于作物数量遗传研究。小麦[7]、玉米[8]和水稻[9]等禾谷类作物中已有大量籽粒矿质含量QTL的研究报道。就水稻而言,已检测到稻米Mg、Ca、Zn、Fe、Mn和Cu含量QTL分别有23、24、64、52、41和29个,分布于水稻的12条染色体上,其中Mg含量QTL热点区域位于第9染色体上,Ca含量QTL热点区域聚集于第10染色体上,Zn含量QTL热点区域聚集于第6、7、8、9和10染色体上,Fe含量QTL热点区域聚集于第1、3和6染色体上,Mn含量QTL热点区域聚集于第1、3、7、8染色体上,而Cu含量QTL热点区域聚集于1、2和4染色体上[9-21]。这些稻米矿质含量QTL大多为初定位结果,精细定位的较少,目前仅有Yu等[18]利用珍汕 97/密阳46近等基因系将精细定位至29.9 kb区域,含有3个候选基因。上述QTL研究所使用的定位材料包括窄叶青 8 号/京系 17、Sasanishiki/Habataki、Ce258/IR75862、ZGX1/IR7586、Lemont/特青、春江06/TN1及Bala/Azucena等籼粳交[9-15]、Madhukar/Swarna、PAU201/Palman 579、珍汕97/明辉63及珍汕97/密阳46等籼籼交[16-19]、特青/云南野生稻[20]野栽交和红香1号/松98-131粳粳交[21]等不同类型的遗传群体,主要为亚种间组合群体,野栽交种间组合群体较少[20],说明野生稻稻米矿质含量基因发掘研究还应进一步加强。
由于过去水稻育种目标过分集中于提高产量,忽视其矿质元素含量[22],在一定程度上导致所育成水稻品种的稻米矿质营养含量较低。研究表明地方品种和野生稻等种质资源的稻米矿质含量高于栽培稻[23,24],是水稻矿质含量改良育种的重要资源。野生稻为水稻遗传改良提供了丰富的基因库[25],如Garcia-Oliveira等[20]研究表明31个稻米矿质含量QTL中26个(83.9%)增效等位基因来自野生稻,因此,从野生稻中发掘有利等位基因可为栽培稻的稻米矿质含量遗传改良提供有效途径。
东乡野生稻是全球分布最北(28°14' N)的普通野生稻,蕴含丰富的重要农艺性状有利等位基因[26],利用这一珍稀水稻资源开展稻米矿质含量QTL研究具有重要意义。本研究利用协青早B和东乡野生稻杂交衍生的BC2F4:5群体,开展糙米矿质含量QTL分析,旨在鉴定控制矿质含量QTL,并分析东乡野生稻有利等位基因在遗传改良上的前景。
1 材料与方法
1.1 水稻材料
本研究所用BC2F4:5株系材料的构建过程(图1)如下:以前期研究[26]的协青早B//协青早B/东乡野生稻BC1F5回交重组自交系群体中的1个单株A58为母本,以协青早B为父本,回交1次并自交1次,获得415个BC2F2单株,并自交产生415个BC2F2:3株系;利用169个SSR标记检测由30个BC2F2:3株系组成的3个DNA混池,发现47个SSR标记位点(表1)呈杂合,其余122标记位点均呈纯合。利用上述47个SSR标记检测415个BC2F2:3株系,结合农艺性状考查,得到4个候选株系;进一步检测4个候选株系各6个单株,挑选到农艺性状接近于亲本协青早B的5个BC2F3单株,自交收获种子;种植其自交产生的5个BC2F4分离群体,每个群体种植72个单株,经农艺性状考查和SSR标记检测后,挑选到132个BC2F4单株,自交产生132个BC2F4:5株系。

图1 BC2F4:5材料构建过程
Fig. 1. Procedure for developing the BC2F4:5population.
1.2 SSR标记多态性分析
前期构建的协青早B//协青早B/东乡野生稻BC1F5群体遗传图谱中共包含41个RFLP标记和108个SSR标记[26],针对41个RFLP标记所处区间,应用Gramene数据库(http://www.gramene.org)中的110对SSR引物,开展多态性筛选,以便应用SSR标记替代RFLP标记。由于协青早B//协青早B/东乡野生稻BC1F5群体的原始杂交亲本东乡野生稻单株丢失,故随机挑选30份BC2F2:3株系DNA等量混合,组成3个DNA混池,结合亲本协青早B共计4份DNA模板进行多态性检测。在协青早B和3个DNA混池间筛选到61个呈多态的SSR标记,结合BC1F5群体遗传图谱中108个SSR标记,总共169个SSR标记用于BC2F2:3群体基因型检测。
1.3 田间试验
2012年11月-2013年4月,在海南陵水种植415个BC2F2:3株系及亲本协青早B,每个株系种植12个单株。株行距16.7 cm´26.7 cm,正常田间管理。移栽后10 d,各株系混取中间10株幼苗叶片,用于DNA提取。 2013年5-10月在浙江杭州种植5个BC2F3单株自交产生的BC2F4群体,每群体种植72个单株。株行距16.7 cm´26.7 cm,正常田间管理。移栽后10 d,每株幼苗取约2 cm叶片,用于DNA提取。2014年5-10月在浙江杭州种植132个BC2F4:5株系及亲本协青早B,重复2次。每个重复中各株系种植1行12个单株,株行距16.7 cm´26.7 cm,完全随机区组设计,正常田间管理。移栽后10 d,选1个重复各株系混取10株幼苗叶片,约2 cm,用于DNA提取。采用简易法[27]提取所有试验材料总DNA。
1.4 标记检测
PCR扩增反应在PCR热循环仪上进行。PCR反应体系含10 μL溶液,包含5.0 μL 2´预混合溶液(CWBIO,北京康为世纪生物科技有限公司)、3 μL 去RNase酶水、1 μL DNA模板和3.3 ng/μL SSR引物各0.5 μL。PCR扩增条件:94℃下预变性2 min;94℃下变性30 s,55℃下退火30 s,72℃下延伸30 s,共循环29次;最后72℃下延伸2 min。PCR扩增产物检测视带型片段大小和清晰度,分别采用2.5%琼脂糖胶分离后GelRed染色和6%非变性聚丙烯酰胺凝胶电泳分离后银染显色进行检测。
基因型鉴定按如下进行:协青早B带型记为“1”,东乡野生稻带型记为“2”,杂合带型记为“3”,缺失带型记为“0”。
1.5 糙米米粉制备和矿质含量测定
在2014年浙江杭州试验点,待稻谷成熟后,2次重复中各株系混收中间5株稻穗,晒干脱粒后,置于常温下保存3个月。称取20 g稻谷,用THU-35A型砻谷机(日本佐竹公司)脱壳成糙米,再将糙米用1093型旋风式磨粉机(瑞典FOSS公司)磨制成粉,过0.18 mm孔径的不锈钢网筛(80目筛),得到糙米米粉样品,装入塑料密封袋中待测。
准确称取0.500 g(± 0.002 g)米粉试样置于25 mL聚丙烯塑料管中,加入8 mL 68%~70%硝酸(HNO3)。将聚丙烯塑料管中混合溶液放入置于通风柜中的DigiBlock ED54型石墨消解仪(LabTech中国公司)在110℃下加热消化,约2.5 h。溶液加热直至不再产生棕色气体且变为清亮无色,溶液剩余体积2 mL左右取出,切不可蒸干。再加入3 mL 30%双氧水(H2O2)混合摇匀,继续在110℃下消化0.5 h,溶液剩余体积1 mL左右取出,切不可蒸干。待消化残留液冷却至室温后,用去离子水定容至25 mL,上下摇匀。同时做空白试剂和大米米粉标准物质(GW10010)试验。
吸取10 mL消化溶液置于15 mL玻璃管中待测,利用去离子水、标准液制作测定标准曲线后,IRIS Intrepid Ⅱ XSP型原子发射光谱仪(Inductively coupled plasma atomic emission spectrometer,ICP-AES)进行测定,并在工作曲线上计算出米粉样品中的Mg、Ca、Zn、Fe、Mn和Cu等矿质含量。

表1 用于BC2F4:5群体基因型检测的47对SSR标记
1.6 数据分析
在前期的遗传图谱基础上[26],将检测到新的61个SSR标记加密到遗传图谱,利用MapMaker 3.0软件构建含169个SSR标记的遗传图谱。以2个重复中各性状数据的均值为基础进行数据分析,结合新构建的遗传图谱,应用Windows QTL Cartographer 2.5软件[28],采用复合区间作图法(composite interval mapping,CIM)进行QTL分析,以LOD=3.0作为QTL阈值,将LOD值最高处所对应的加性效应、显性效应和贡献率记为该QTL的效应。QTL命名遵循McCouch和CGSNL[29]提出的命名法则。
2 结果与分析
2.1 BC2F4:5群体的表型变异
BC2F4:5群体及亲本协青早B的糙米6种矿质含量基本统计参数列于表2。除Fe含量外,其余5种糙米矿质含量分布均呈正态分布。6种糙米矿质含量的遗传变异均较小,其中Mg含量的变异系数最小,为6.54%;而Ca含量的变异系数最大,为12.33%。与亲本协青早B相比,群体的Mg、Ca、Zn、Fe、Mn和Cu含量均值分别高9.21%、9.14%、0.70%、2.10%、11.40%和13.00%。

表2 协青早B3/东乡野生稻BC2F4:5群体糙米矿质含量

表3 协青早B3/东乡野生稻BC2F4:5群体糙米矿质含量间相关分析
*,**分别表示在0.05和0.01水平上显著相关。
*,**significant at 0.05 and 0.01 level, respectively.
2.2 糙米矿质含量间相关性分析
由糙米矿质含量间相关性分析(表3)可知,有较强的证据表明Mg、Ca、Zn、Fe和Mn之间显著性较高,而Cu含量与其余5种糙米矿质含量的相关性较低。具体表现为:除Ca和Zn以及Fe和Mn之间不呈现显著相关外,Mg、Ca、Zn、Fe和Mn含量两两之间均呈极显著正相关;Cu含量仅与Mg和Zn含量呈显著正相关,与Ca含量呈显著负相关。
2.3 糙米矿质含量QTL分析
采用CIM法对6个糙米矿质含量进行QTL分析,以LOD=3.0为QTL阈值,共检测到17个控制糙米矿质含量QTL(表4,图2),分别位于第1、4、6、8、9和11等6条染色体上,LOD介于3.12~18.58。单个QTL解释糙米矿质含量的表型贡献率介于5.0%~47.2%,其中表型贡献率大于10%的QTL有13个,8个QTL增效等位基因来自东乡野生稻。
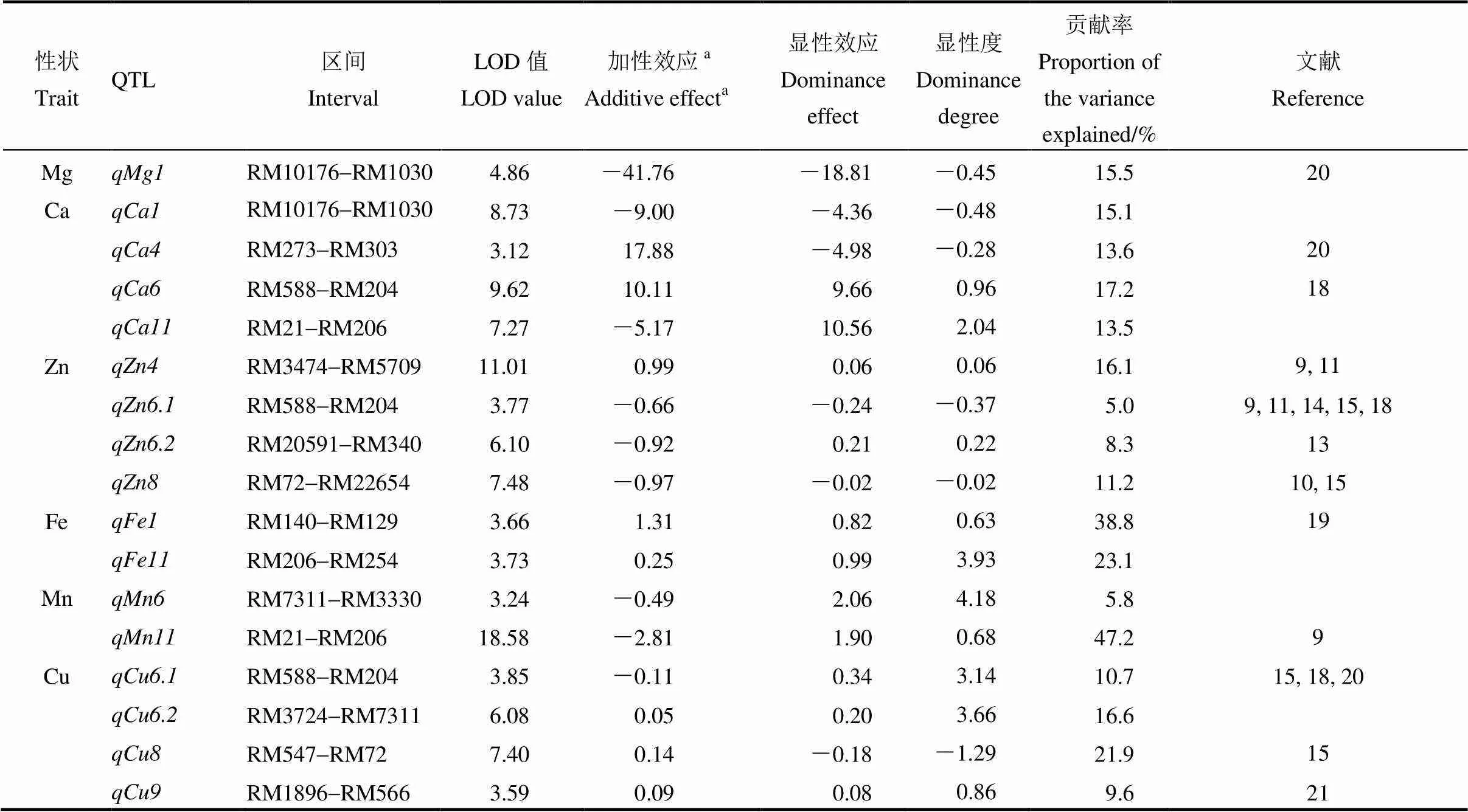
表4 协青早B3/东乡野生稻BC2F4:5群体糙米矿质含量QTL
a加性效应是指东乡野生稻等位基因取代协青早B等位基因。
aAdditive effect of replacing an Xieqingzao B allele by Dongxiang wild rice allele.
本研究仅检测到1个控制糙米Mg含量QTL,位于第1染色体上的RM10176-RM10300区间内,表型贡献率为15.5%,遗传作用模式为部分显性,其协青早B增效等位基因可增加糙米Mg含量41.76 mg/kg。
检测到4个控制糙米Ca含量的主效QTL,分布于第1、4、6和11染色体上,贡献率介于13.5%~17.2%。其中和的遗传作用模式均为部分显性,其增效等位基因分别来自协青早B和东乡野生稻,加性效应分别为9.00 mg/kg和17.88 mg/kg;表现完全显性,加性效应和显性效应分别为10.11 mg/kg和9.66 mg/kg;的遗传作用模式为超显性,增效等位基因来自协青早B,显性效应值为10.56 mg/kg。
共检测到4个糙米Zn含量QTL,分别位于第4、6和8等3条染色体上,贡献率介于5.0%~16.1%。其中和的遗传作用模式均为加性,其增效等位基因分别来自东乡野生稻和协青早B,加性效应为0.99 mg/kg和0.97 mg/kg;其余和的遗传作用模式为部分显性,其增效等位基因均来自协青早B,加性效应分别为0.66 mg/kg和0.92 mg/kg。
检测到2个控制糙米Fe含量QTL,和,表型贡献率分别为38.8%和23.1%,其中的遗传作用模式为部分显性,其东乡野生稻等位基因分别增加糙米Fe含量1.31 mg/kg;的遗传作用模式为超显性,增效等位基因来自东乡野生稻,显性效应值为0.99 mg/kg。
检测到2个控制糙米Mn含量QTL,分别位于第6和11染色体上,表型贡献率分别为5.8%和47.2%。其中,的遗传作用模式为超显性,增效等位基因来自协青早B,显性效应值为2.06 mg/kg;的遗传作用模式为部分显性,增效等位基因来自协青早B,加性效应值和显性效应值分别为2.81 mg/kg和1.09 mg/kg。
检测到4个控制糙米Cu含量QTL,分别位于第6、8和9等3条染色体上,其贡献率介于9.6%~21.9%。其中、和的遗传作用模式均为超显性,前者的增效等位基因来自协青早B,后两者的增效等位基因来自东乡野生稻,显性效应值分别为0.34、0.20和0.18 mg/kg;的遗传作用模式为显性,增效等位基因均来自东乡野生稻,显性效应值为0.08 mg/kg。
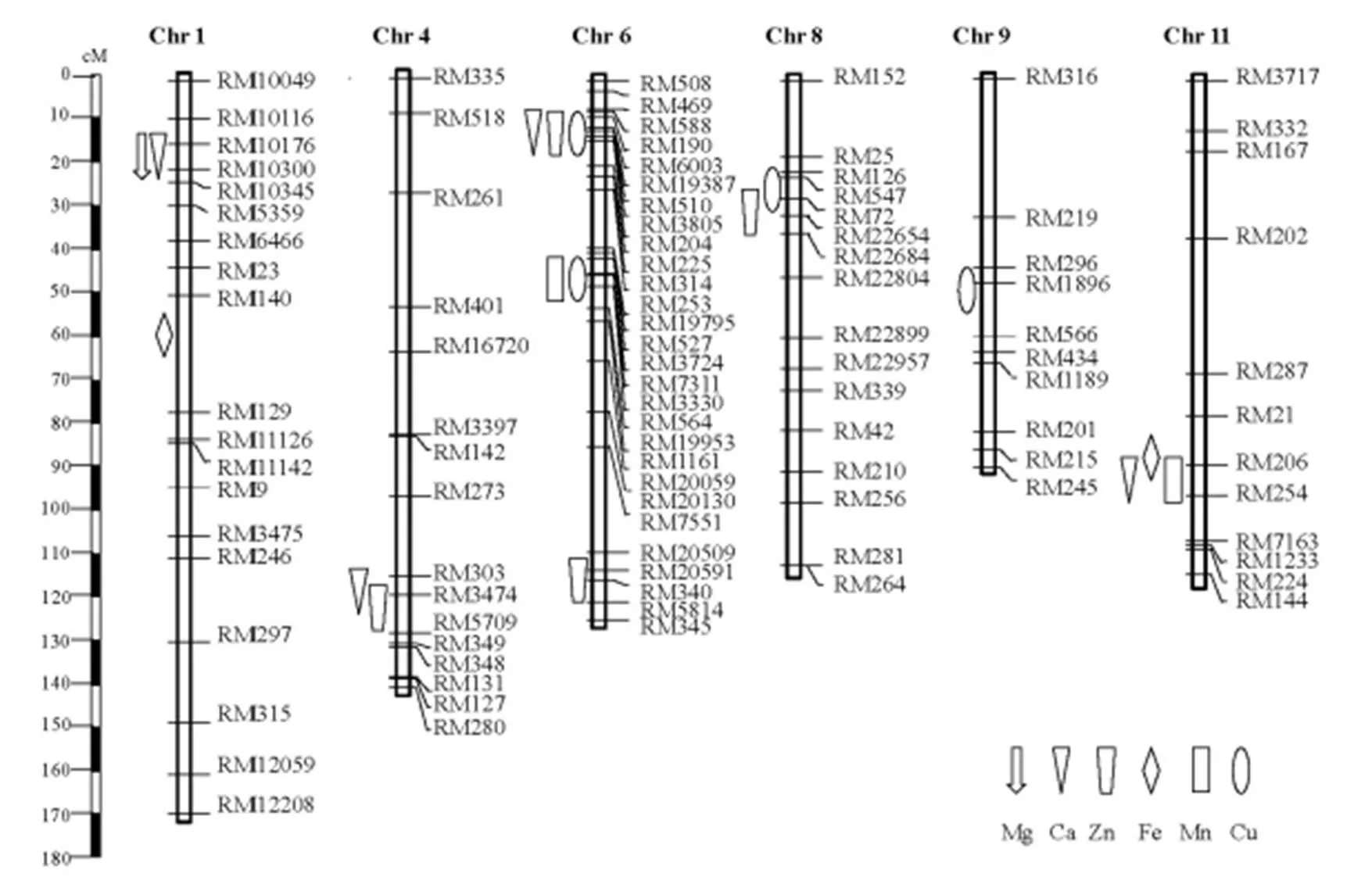
图2 糙米Mg、Ca、Zn、Fe、Mn及Cu含量QTL的染色体位置
Fig. 2. Chromosomal positions of the QTLs conferring Mg, Ca, Zn, Fe, Mn and Cu contents in brown rice.
3 讨论
近年来,随着人们膳食结构和生活方式的变化,矿质营养缺乏已成为影响人体健康的重要因素。由于稻米的全球消费量大,稻米矿质含量略微增加便可有效缓解人类矿质元素缺乏症[6]。理解稻米矿质含量遗传变异和矿质含量等位基因间的差异对稻米矿质含量的遗传改良非常重要。人们习惯食用稻米部位为精米,尽管其矿质含量低于糙米[30],但与糙米矿质含量密切相关;其次,高矿质含量稻米可以通直接食用糙米或加工成米制品以提高稻米矿质含量的利用效率。为此,本研究利用野生稻和栽培稻种杂交构建BC2F4:5群体开展糙米矿质含量QTL分析。
3.1 糙米矿质含量间的相关性
稻米矿质含量间的相关性已有较多报道。稻米Mg含量与Ca含量[18, 31, 32]、Zn含量[18,31]、Fe含量[31,33],Mn含量[18,31-33]及Cu含量[18]呈显著或极显著正相关。稻米Ca含量与Zn、Fe含量[18, 31, 33]及Mn含量[18, 31-34]呈极显著正相关,而与Cu含量[33]呈极显著负相关。稻米Zn含量与Fe含量[23, 31-33]、Mn含量[18,23]及Cu含量[18, 32, 33]均呈显著或极显著正相关。稻米Fe含量与Cu含量[34]及Mn含量[23, 31, 33]均呈显著或极显著正相关。稻米Mn含量与Cu含量均呈显著或极显著正相关[18,31, 33]。
本研究结果表明,Mg含量与Ca、Zn、Fe及Mn含量呈极显著正相关,与Cu含量呈显著正相关;Ca含量与Fe和Mn含量呈极显著正相关,而与Cu含量呈显著负相关;Zn含量与Fe和Mn含量呈极显著正相关,与Cu含量呈显著正相关。上述结果与前人结果较为一致,尤其是Mg、Zn和Mn含量间均呈显著或极显著正相关,表明这些矿质含量间存在稳定的协同效应,将有助于稻米不同矿质元素的聚合改良,应在育种实践中得到重视。
3.2 糙米矿质含量QTL
本研究共检测到17个糙米矿质含量QTL,其中8个QTL的有利等位基因来自东乡野生稻,这些有利的野生稻等位基因成功导入到协青早B背景将促进稻米矿质营养的遗传改良。
在17个QTL中,有13个QTL与前人报道的稻米矿质含量QTL区间重叠或相近(表3)。其中Mg含量和Fe含量QTL各1个QTL,即与Garcia-Oliveira等[20]报道的糙米Mg含量QTL重叠,位于Anuradha等[19]检测到的糙米Fe含量QTL区间内。Ca含量和Mn含量QTL各2个,即和分别与糙米含量QTL[20]和糙米及精米Ca含量QTL[18, 35]相近,和分别与糙米Mn含量QTL[9]和精米Mn含量QTL[18]部分重叠。Cu含量QTL有3个,即分别与2个报道中的糙米Cu含量QTL[15, 20]及Yu等[18]报道的精米Cu含量QTL重叠;位于Norton等[15]检测到的标记区间内;与黄莹莹等[21]报道的糙米Cu含量QTL相同。Zn含量QTL有4个,分别为与2个报道中的糙米Zn含量QTL[9, 11]区间一致;与3个研究中的糙米Zn含量[9, 11, 15]和2个研究中的精米Zn含量QTL[14, 18]部分重叠;位于Xu等[13]检测的精米Zn含量QTL区间内;与糙米Zn含量QTL[10, 16]和Xu等[13]检测的精米Zn含量QTL重叠。值得注意是,本研究检测到LOD值最高的矿质含量QTL均在前人相同元素含量QTL定位研究中被检测到。上述在不同环境和遗传背景中共同检测到的控制稻米矿质含量染色体区间,可能是有潜在利用价值的稻米含量QTL。此外,由于本研究利用BC2F4:5分离群体的遗传背景较为纯合(纯合率为72.19%),一定程度上也导致已报道的位于这些纯合区域的稻米矿质含量QTL在本研究中未被检测到。
在QTL定位研究中,相关性状QTL通常成簇分布于相同或相邻的染色体区间内,这可能是由基因的多效性和多个基因的紧密连锁引起的。本研究检测到的17个糙米矿质含量QTL分别聚集于6条染色体上的5个QTL簇中,这也证实了QTL多效性的存在。控制3种不同糙米矿质含量QTL分别位于第6和第11染色体上的2个QTL簇中,如控制Ca、Zn和Cu含量QTL位于第6染色体的RM588-RM204区间和控制Ca、Fe和Mn含量QTL第11染色体上的RM206-RM254区间。控制2个不同稻米矿质含量QTL位于第1、6和8染色体上的3个QTL簇,如控制Ca和Zn含量QTL位于第4染色体上RM273-RM5709区间,且这2个QTL的有利等位基因均来自东乡野生稻。由上述分析可知,除了第1染色体上的RM10176-RM10300区间和第4染色体上的RM273-RM5709区间的QTL增效等位基因方向一致外,其余3个QTL簇中的QTL增效等位基因方向不同,表明对某些性状有利的野生稻等位基因可能对另一些性状的影响不利。在5个QTL簇中,QTL簇均为主效QTL,且其有利等位基因来自东乡野生稻,将成为进一步精细定位和克隆的候选对象;相应地,从群体中挑选到含该QTL的株系108,其糙米Ca和Zn含量分别为142.80 mg/kg和32.06 mg/kg,该株系可应用于稻米Zn和Ca含量改良育种。由于初定位区间较大,一般难以判断QTL簇的成因。因此,需将QTL簇中的不同QTL分解到更小的区间内,以探究QTL的一因多效机理以及不同性状QTL簇的存在形式,可为多性状的同步改良或标记辅助选择提供有力的手段,如同时开展稻米中多个矿质元素的聚合改良。
谢辞: 感谢中国水稻研究所庄杰云研究员对本研究的悉心指导!
[1] Stein A J. Global impacts of human mineral malnutrition., 2010, 335(1/2): 133-154.
[2] White P J, Brown P H. Plant nutrition for sustainable development and global health., 2010, 105(7): 1073-1080.
[3] Rawat N, Neelam K, Tiwari V K, Dhaliwal H S. Biofortification of cereals to overcome hidden hunger., 2013, 132: 437-455.
[4] Bashir K, Takahashi R, Nakanishi H, Nishizawa N K. The road to micronutrient biofortification of rice: Progress and prospects., 2013, 4: 15.
[5] Bhullar N K, Gruissem W. Nutritional enhancement of rice for human health: The contribution of biotechnology., 2013, 31(1): 50-57.
[6] Gregorio G B, Senadhira D, Htut T. Breeding for trace mineral density in rice., 2000, 21(4): 382-386.
[7] Srinivasa J, Arun B, Mishra V K, Singh G P, Velu G, Babu R, Vasistha N K, Joshi A K. Zinc and iron concentration QTL mapped in a×.cross., 2014, 127(7): 1643-1651.
[8] Šimić D, Mladenovic D S, Zdunic Z, Jambrović A, Ledencan T, Brkić J, Brkić A, Brkić I. Quantitative trait loci for biofortification traits in maize grain., 2012, 103: 47-54.
[9] Zhang M, Pinson S R M, Tarpley L, Huang X Y, Lahner B, Yakubova E, Baxter I O, Guerinot M L, Salt D E. Mapping and validation of quantitative trait loci associated with contents of 16 elements in unmilled rice grain., 2014 127: 137-165.
[10] Du J, Zeng D L, Wang B, Qian Q, Zheng S, Ling H Q. Environmental effects on mineral accumulation in rice grains and identification of ecological specific QTLs., 2013, 35(2): 161-170.
[11] Zhang X Q, Zhang G P, Guo L B, Wang H, Zeng D L. Identification of quantitative trait loci for Cd and Zn concentrations of brown rice grown in Cd-polluted soils., 2011, 180(2): 173-179.
[12] Ishikawa S, Abe T, Kuramata M, Yamaguchi M, Ando T, Yamamoto T, Yano M. A major quantitative trait locus for increasing cadmium-specific concentration in rice grain is located on the short arm of chromosome 7., 2010, 61: 923-934.
[13] Xu Q, Zheng T Q, Hu X, Cheng L R, Xu J L, Shi Y M, Li Z K. Examining two sets of introgression lines in rice (L.) reveals favorable alleles that improve grain Zn and Fe concentrations., 2015, 10(7): e0131846. doi:10.1371/journal.pone.0131846.
[14] Norton G J, Deacon C M, Xiong L Z, Huang S Y, Meharg A A. Genetic mapping of the rice ionome in leaves and grain: Identification of QTLs for 17 elements including arsenic, cadmium, iron and selenium., 2010, 329(1/2): 139-153.
[15] Norton G J, Duan G L, Lei M, Zhu Y G, Meharg A A, Price A H. Identification of quantitative trait loci for rice grain element composition on an arsenic impacted soil: Influence of flowering time on genetic loci., 2012, 161(1): 46-56.
[16] Kumar K, Jian S, Jian R K. Linkage mapping for grain iron and zinc content in F2population derived from the cross between PAU201 and Palman 579 in rice (L.)., 2014, 42(3): 389-400.
[17] Lu K Y, Li L Z, Zheng X F, Zhang Z, Mou T, Hu Z. Quantitative trait loci controlling Cu, Ca, Zn, Mn and Fe content in rice grains., 2008, 87(3): 305-310.
[18] Yu Y H, Shao Y F, Liu J, Fan Y Y, Sun X C, Cao Z Y, Zhuang J Y. Mapping of quantitative trait loci for contents of macro- and microelements in milled rice (L.)., 2015, 63, 7813-7818.
[19] Anuradha K, Agarwal S, Rao K V, Rao K V, Viraktamath B C, Sarla N. Mapping QTLs and candidate genes for iron and zinc concentrations in unpolished rice of Adhukar´Swarna RILs., 2012, 508(2): 233-240.
[20] Garcia-Oliveira A L, Tan L B, Fu Y C, Sun C Q. Genetic identification of quantitative trait loci for contents of mineral nutrients in rice grain., 2009, 51: 84-92.
[21] 黄莹莹, 邹德堂, 王敬国, 刘化龙, 兴旺, 马婧, 武琦. 水稻子粒锰、铁、锌、铜含量的QTL定位分析. 作物杂志, 2012(6): 77-81.
Huang Y Y, Zou D T, Wang J G, Liu H L, Xing W, Ma J, Wu Q. QTL mapping for Mn, Fe, Zn and Cu contents in rice grains., 2012(6): 77-81. (in Chinese with English abstract)
[22] Sands D C, Morris C E, Dratz E A, Pilgeram A. Elevating optimal human nutrition to a central goal of plant breeding and production of plant-based foods., 2009, 177(5): 377-389.
[23] Anandan A, Rajiv G, Eswaran R, Prakash M. Genotypic variation and relationships between quality traits and trace elements in traditional and improved rice (L.) genotypes., 2011, 76(4): 122-130.
[24] Jiang S L, Shi C H, Wu J G. Studies on mineral nutrition and safety of wild rice (L.)., 2009, 60(s1): 139-147.
[25] Tanksley S D, McCouch S R. Seed banks and molecular maps: Unlocking genetic potential from the wild., 1997, 277: 1063-1066.
[26] 黄得润, 陈洁, 侯丽娟, 樊叶杨, 庄杰云. 协青早B//协青早B/东乡野生稻BC1F5群体产量性状QTL分析. 农业生物技术学报, 2008, 16(6): 977-982.
Huang D R, Chen J, Hou L J, Fan Y Y, Zhuang J Y. Identification of QTLs for yield traits in the BC1F5population of Xieqingzao B//Xieqingzao B/Dongxiang wild rice., 2008, 16(6): 977-982. (in Chinese with English abstract)
[27] Zheng K L, Huang N, Bennet J. PCR-based marker-assisted selection in rice breeding//IRRI Discussion Paper Series No. 12. Manila: IRRI, 1995: 1-24.
[28] Wang S C, Basten C J, Zeng Z B. Windows QTL cartographer 2.5. Raleigh, NC: Department of Statistics, North Carolina State University, 2012. http://statgen. ncsu.edu/qtlcart/WQTLCart.htm.
[29] McCouch S R, CGSNL. Gene nomenclature system for rice., 2008, 1: 72-84.
[30] Jiang S L, Wu J G, Thang N B, Feng Y, Yang X E, Shi C H. Genotypic variation of mineral elements contents in rice (L.)., 2008, 228: 115-122.
[31] Jiang S L, Wu J G, Feng Y, Yang X E, Shi C H. Correlation analysis of mineral element contents and quality traits in milled rice (L.)., 2007, 55(23): 9608-9613.
[32] 孙明茂, 杨仁昌, 李点浩, 崔仁守, 曹桂兰, 李圭星, 金弘烈, 于元杰, 李英泰, 韩龙植. 粳稻“龙锦1号/香软米1578”F3家系群糙米矿质元素含量变异及相关性分析. 中国水稻科学, 2008, 22(3): 290-296.
Sun M M, Yang R C, Li D H, Choi I S, Cao G L, Lee K S, Kim H Y, Yu Y J, Lee Y T, Han L Z. Variation and correlation analysis of mineral element contents in brown rice of F3lines from japonica combination “Longjin 1/Xiangruanmi 1578”., 2008, 22(3): 290-296. (in Chinese with English abstract)
[33] 曾亚文, 申时全, 汪禄祥, 刘家富, 普晓英, 杜娟. 云南稻种矿质元素含量与形态及品质性状的关系. 中国水稻科学, 2005, 19(2): 127-131
Zeng Y W, Shen S Q, Wang L X, Liu J F, Pu X Y, Du J. Relationship between morphological and quality traits and mineral element content in Yunnan rice., 2005, 19(2): 127-131. (in Chinese with English abstract)
[34] Pinson S R M, Tarpley L, Yan W G, Yeateref K, Lahnerg B, Yakubovah E, Huangi X Y, Zhangj M, Mary G M L, Salti D E. Worldwide genetic diversity for mineral element concentrations in rice grain., 2015, 55(1): 294-311.
[35] Huang Y, Tong C, Xu F F, Chen Y L, Zhang C Y, Bao J S. Variation in mineral elements in grains of 20 brown rice accessions in two environments., 2016, 192: 873-878.
QTL Analysis for Mineral Contents in Brown Rice Using a BC2F4:5Population Derived from Dongxiang Wild Rice (Griff.)
HU Biaolin1, 2, HUANGDerun1, XIAO Yeqing2, HE Qiangsheng3, WAN Yong2,*, FAN Yeyang1,*
(1,,,;2,,,;Jiangxi Xingan Seed Industry Co. Ltd, Shangrao 334300, China;,:)
Biofortifying food crops with essential minerals would alleviate mineral deficiencies in humans.One plant A58 was selected from Xieqingzao B// Xieqingzao B /Dongxiang wild rice BC1F5population, and was backcrossed with Xieqingzao B to develop a BC2F4:5population. The contents of Mg, Ca, Zn, Fe, Mn and Cu in brown rice of 132 BC2F4:5lines were measured with an inductively coupled plasma atomic emission spectrometer (ICP-AES). Detection of quantitative trait loci (QTLs) for mineral contents in brown rice was conducted using Windows QTL Cartographer 2.5.A total of 17 QTLs for mineral contents in brown rice were detected on chromosomes 1, 4, 6, 8, 9 and 11, respectively, including one for Mg content, four for Ca content, four for Zn content, two for Fe content, two for Mn content, and four for Cu content. The explained phenotypic variations ranged from 5.0% to 47.2%, and eight QTLs of them had the enhancing alleles derived from.Twelve QTLs were clustered in five chromosomal regions, indicating that common genetic-physiological mechanisms were involved for different mineral nutrients, and the beneficial alleles could be utilized to improve grain nutritional quality by marker-assisted selection.
Dongxiang wild rice; brown rice; mineral content; QTL mapping
10.16819/j.1001-7216.2018.7011
Q343.1+5; S511.03
A
1001-7216(2018)01-0043-08
2017-01-19;
2017-04-02。
国家863计划资助项目(2014AA10A604);江西省重点研发计划重大专项(20161ACF60022);江西省自然科学基金资助项目(20171ACB21071);江西省农业科学院创新基金资助项目(20161CBS002)。
