雷帕霉素抑制光老化成纤维细胞周期阻滞的研究
2018-01-15秦登科贾传龙卢勇舟杨清建陈亮吴心愿任润健朱晶晶郭妤杨平周轶群毕波朱宁文刘天一
秦登科,贾传龙,卢勇舟,杨清建,陈亮,吴心愿,任润健,朱晶晶,郭妤,杨平,周轶群,毕波,朱宁文,刘天一
1.复旦大学附属华东医院整形外科,上海200040;2.复旦大学附属华山医院皮肤科,上海200040
皮肤的衰老除了受内源性因素影响外,也受外源性因素影响,包括吸烟、饮酒和光照等[1-2]。中波紫外线(ultravioletB,UVB)照射是导致皮肤光老化的重要原因,表现为皮肤皱纹增多、色素沉着及失去弹性等[3]。此外,UVB照射还会导致皮肤成纤维细胞(fibroblasts,FBs)正常周期改变,p53和p21等周期相关蛋白分泌增加,破坏细胞正常的复制分裂过程,使细胞停滞在S期,影响细胞正常的结构和功能,加剧细胞老化和凋亡[4-5]。近期许多研究发现,哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)通路的抑制剂雷帕霉素(rapamycin,RAPA),作为一种新型大环内酯类免疫抑制剂,具有调节衰老的重要作用[6]。虽然已有关于RAPA对细胞增殖、周期和信号通路的相关报道,但RAPA在光老化周期阻滞方面的研究甚少。本研究旨在探索RAPA是否能够改善UVB照射导致的小鼠FBs周期阻滞。
1 材料与方法
1.1 小鼠真皮FBs获取 取1~3 d SPF级C57BL/6小鼠,乙醚处死后浸泡于75%酒精中约10min,用培养基(dulbecco'smodifiedeaglemedium,DMEM)洗去残留酒精,放入培养皿内,剪刀分离后背皮肤,剪下面积约为1cm×1cm的皮肤组织,立即置于2%中性蛋白酶内,4℃冰箱过夜。第2天取出皮肤,镊子分离表皮真皮,剪碎真皮后置于0.1%胶原酶中,37℃下消化1.5h,至皮肤的组织基本消失。1500r/min离心5min,收集沉淀,弃上清,细胞以 DMEM+10%胎牛血清(fetalbovineserum,FBS)重悬,反复吹打直至均匀,以2×105/cm2的细胞密度接种在培养皿内,于37℃、5%CO2、100%饱和湿度的培养箱内培养,将近80%~90%融合传代,比例约为1∶4。
1.2 药物预处理及光老化模型建立 选用P1代细胞,用含有不同浓度RAPA的DMEM+10%FBS处理48h。CCK-8试剂盒检测细胞活性,选取最适浓度。实验共分4组:Ctrl组(非RAPA处理,非UVB照射)、RAPA组 (RAPA处理,非 UVB照射)、UVB照射组 (非RAPA处理,UVB照射)、UVB+RAPA组 (RAPA处理,UVB照射)。利用UVB照射建立FBs的光老化模型(stress-induced premature senescence,SIPS)[4],吸除培养液,加入少量磷酸盐缓冲液 (phosphate bufferedsaline,PBS)覆盖细胞,移走培养皿盖,放置在UVB灯 (PhilipsTL20w/01RSlamp)正下方,首次照射剂量(使用Lutron UV light meter测量)为120mJ/cm2,距离细胞7.5cm,照射后吸除PBS,加入10mL DMEM+1%FBS继续培养,照射间隔时间为12h,每次照射时间为120s,共4次,末次照射后改为DMEM+10%FBS 培养[4]。
1.3 -半乳糖苷酶染色 吸除6孔板中的细胞培养基,用PBS洗涤2次后每孔加入0.25 mL的4%多聚甲醛固定液,室温下固定15 min。吸除细胞固定液,用PBS洗涤细胞3次,每孔加0.25 mL染色液 (按照试剂盒说明书现用现配),调整 PH为6.0[7],37℃温箱中孵育过夜。普通光学显微镜下观察,胞质呈蓝色者为阳性细胞,表示细胞处于衰老状态。 -半乳糖苷酶染色阳性率的计算方法为:200倍下每孔中随机选取10个视野,计算阳性细胞总数占总细胞数的百分率。
1.4 细胞周期检测 Ctrl组和UVB照射组末次照射48 h后,吸移培养液,收集细胞悬液,1 500 r/min离心5min,弃去上清液,PBS洗2次,预冷的70%乙醇吹打均匀后重悬,4℃下固定过夜后,PBS洗3次,0.5 mL PBS重悬。加入50g/mL碘化丙啶和1mg/mL的RNA酶,避光37℃孵育30min后流式细胞仪检测。细胞周期的分布使用ModiFit LT v2.0软件进行分析。
1.5 周期相关蛋白检测 正常细胞或SIPS细胞最后一次照射48h后,移去培养液,PBS洗涤3次,加入细胞裂解液500L,提取总蛋白。应用二喹啉甲酸法通过微量紫外分光光度计测定蛋白浓度,变性后取40 g,经10%聚丙烯酰胺电泳分离蛋白并将蛋白质转移至聚偏氟乙烯膜上,孵育袋中加入稀释的 p53、p21(艾博抗1∶500)和 -微管蛋白 -tublin(博士德1∶2000),4℃孵育过夜。采用辣根过氧化物酶标记的羊抗兔二抗 (艾博抗1∶2000)室温孵育2h。增强型的化学发光法显影和定影,利用灰度分析法检测蛋白条带的相对蛋白含量。
1.6 统计学分析 采用SPSS13.0统计软件包进行分析,所有数据以均数±标准差(±s)表示,各组间的比较采用方差分析,<0.05为差异有统计学意义。
2 结果
2.1 RAPA对小鼠FBs活性的影响 为选择最适宜的RAPA浓度处理细胞并进行实验,分别使用含有不同浓度RAPA的DMEM+10%FBS处理小鼠FBs。发现48h处理时间内,较低浓度的RAPA(0~5mol/L)并不影响细胞活性,而较高浓度的 RAPA(10mol/L)会产生细胞毒性(<0.05),抑制细胞增殖。因此选择5 mol/L的RAPA作为后续细胞处理药物浓度。见图1。

图1 不同浓度RAPA对小鼠FBs活性的影响
2.2 RAPA降低光老化细胞的衰老染色阳性率 UVB照射组衰老染色明显,主要集中在核周,呈蓝绿色。RAPA预处理后的UVB照射组中阳性染色细胞显著减少,差异有统计学意义(<0.05),正常细胞组和RAPA处理的正常细胞组鲜有染色。见图2。
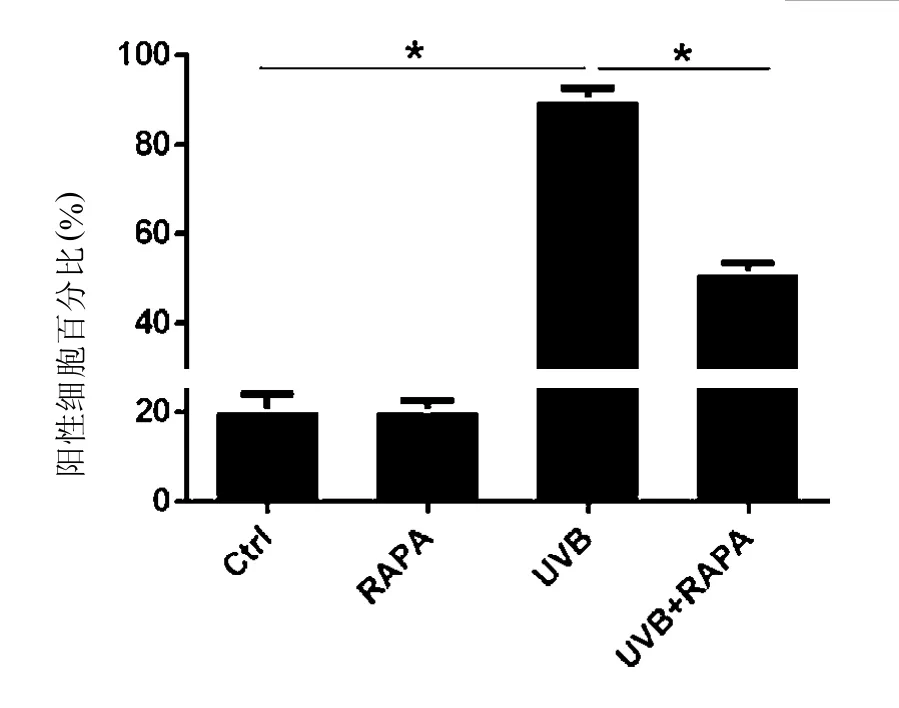
图2 UVB照射后SA--Gal染色阳性率
2.3 各组FBs周期 流式细胞仪检测细胞周期后发现UVB照射组S期的细胞比率是Ctrl组的2.2倍,说明UVB照射后的细胞发生了明显的S期阻滞。而UVB+RAPA组细胞处于S期的比率为26.3%,与UVB照射组相比下降了6.5%,差异有统计学意义(<0.05),可见RAPA预处理可以缓解UVB照射导致的S期阻滞,而RAPA预处理后对Ctrl组细胞的S期没有影响。见图3。2.4RAPA可以降低光老化FBs周期蛋白p53和p21的生成 p53和p21是细胞周期相关蛋白,也是作为细胞衰老的常用指标。Western Blot结果显示与正常细胞组相比,UVB照射组p53和p21的表达量分别是Ctrl组的1.9倍和1.8倍,UVB+RAPA组p53和p21的表达量是Ctrl组的1.4倍和1.3倍,说明RAPA预处理后减少了UVB照射组p53和p21的表达,差异有统计学意义(<0.05),而RAPA预处理组p53和p21的表达量与Ctrl组比,差异无统计学意义。见图4。
3 讨论
UVB照射可以破坏皮肤的正常结构和功能,并能引起皮肤提前衰老,造成皮肤的粗糙、干燥、皱纹增多以及色素沉着等[8]。真皮的FBs是皮肤的主体细胞,除了衍生真皮外,还可以形成细胞外基质,对维持皮肤的结构、功能以及伤口的修复和愈合起着至关重要的作用[9]。长期暴露在UVB照射下,不仅会导致皮肤老化甚至会诱发皮肤癌[10]。UVB照射可以直接损伤细胞DNA,诱导产生细胞毒性物质(卟啉、胆红素、黑色素、氧自由基和活性氧簇等),造成细胞周期阻滞,干扰细胞的增殖进程,影响的正常细胞功能[11]。
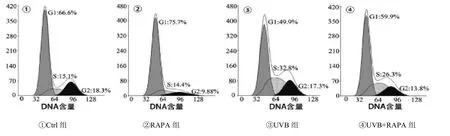
图3 流式检测各组FBs周期
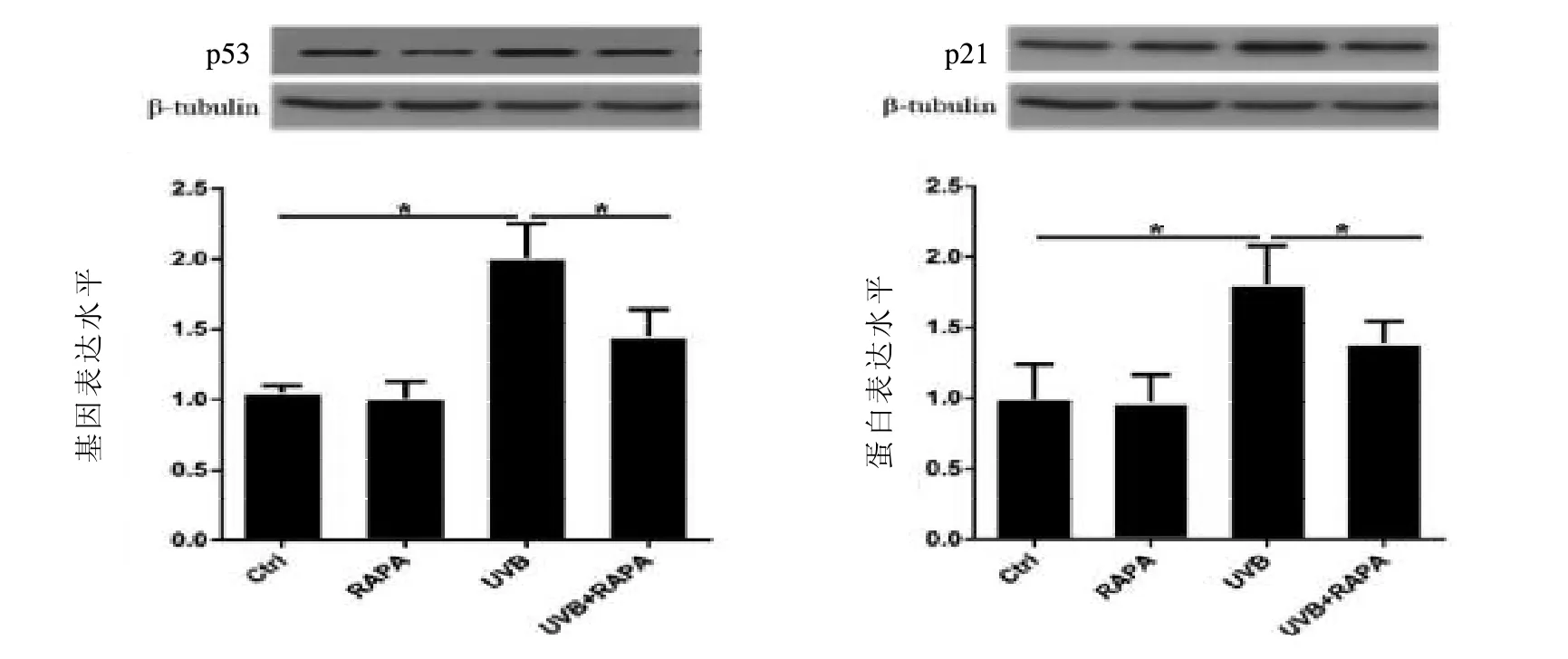
图4 RAPA对UVB诱导的FBs周期相关蛋白表达的影响
细胞内DNA可以直接吸收UVB中的光量子,造成DNA突变和核苷酸结构重排,导致缺陷DNA链的产生,影响DNA的正常复制过程,干扰细胞增殖的正常进程[12],细胞内的这些改变可以引起包括细胞增殖、分化、老化、周期阻滞及DNA损伤修复在内的多条信号通路激活[13]。而Bhatia-Dey等[14]的研究发现,细胞可以通过周期阻滞来进行DNA修复,是细胞对DNA损伤的一种反馈表现,通过阻滞细胞周期防止细胞DNA的进一步损害。p53和p21等抑癌基因可以调控细胞周期,是激活衰老信号通路的中枢信号[15]。UVB导致的DNA损伤可以激活p53和p21基因,导致视网膜母细胞瘤蛋白的去磷酸化,抑制细胞周期蛋白和细胞周期蛋白依赖激酶的合成,使细胞进入一个稳定的周期阻滞期,加剧老化的进程[12,14]。UVB照射可以导致线粒体内产生大量的活性氧簇(reactiveoxygenspecies,ROS),影响细胞内脂质、蛋白质和DNA的合成,干扰细胞间正常的信号通路[16]。p53和p21表达增多可以激活丝裂原活化蛋白激酶和转化生长因子-通路,进一步加速 ROS生成。ROS一方面直接破坏DNA,另一方面可以通过激活核因子appaB和激活蛋白-1等信号通路促进基质金属蛋白酶(matrix metalloproteinases,MMPs)的分泌,MMPs可以降解细胞外基质,破坏细胞的正常结构和功能[17]。在本实验中,UVB照射组发生了明显的 S期阻滞,并且p53和p21蛋白的分泌显著增加也符合细胞衰老的生物学特征,进一步验证了 SIPS模型的合理性。
RAPA是一种大环内酯类化合物,主要用来抑制器官移植后产生的免疫排斥反应。近期研究发现RAPA在改善氧化应激反应和延缓衰老过程等方面发挥着重要作用[18]。RAPA可以通过结合 mTOR受体诱导自噬,吞噬细胞内衰老的蛋白、功能受损的细胞器、代谢废物及过氧化物酶体等来维持细胞的正常功能[18]。已经有实验证明,RAPA可以明显延长哺乳动物的生存期,调控正常的生理衰老进程,机制可能与其诱导的自噬有关[19]。同时Finkel等[20]的研究证明,RAPA预处理后可以减少 ROS和总体炎症因子产生,降低DNA破坏水平,从而减弱辐射对上皮干细胞造成的损伤。而Sonis等[21]发现,RAPA可以通过减少细胞内ROS的生成来降低DNA破坏水平,抑制衰老相关蛋白p53和p21产生,维持细胞周期的正常进行。
[1]Kohl E,SteinbauerJ,Landthaler M, .Skinageing[J].JEur Acad Dermatol Venereol,2011,25(8):873-884.
[2] 许阳,骆丹.皮肤衰老的研究新进展 [J].老年医学与保健,2016,22(6):334-338.
[3] 陈亮,毕波,曾继平,等.罗格列酮对UVB诱导的小鼠光老化皮肤成纤维细胞MMPs表达和细胞外基质蛋白合成的影响 [J].中国美容整形外科杂志,2015,26(8):500-503.
[4]ZengJP,BiB,ChenL, .Repeatedexposureofmousedermal fibroblasts at a sub-cytotoxic dose of UVB leads to premature senescence:arobust model of cellular photoaging[J].JDermatol Sci,2014,73(1):49-56.
[5] ChenL,BiB,ZengJ, .Rosiglitazoneamelioratessenescencelike phenotypes in a cellular photoaging model[J].J Dermatol Sci,2015,77(3):173-181.
[6] Demidenko ZN,Zubova SG,Bukreeva EI, .Rapamycin decelerates cellular senescence[J].Cell Cycle,2009,8(12):1888-1895.
[7] 陈亮,毕波,曾继平,等.pH对衰老相关 -半乳糖苷酶染色的影响[J].中国美容整形外科杂志,2014,25(12):751-754.
[8] Wang XF,Huang YF,Wang L, .Photo-protective activity of pogostone against UV-induced skin premature aging in mice[J].Exp Gerontol,2016,77:76-86.
[9] 卢勇舟,陈亮,毕波,等.胶原蛋白碎片对光老化真皮细胞外基质的影响 [J].中国美容整形外科杂志,2015,26(4):213-215.
[10] Tobin DJ.Introduction to skin aging[J].J Tissue Viability,2017,26(1):37-46.
[11]Schuch AP,Moreno NC,Schuch NJ, .Sunlight damage to cellular DNA:Focus on oxidatively generated lesions[J].Free Radic Biol Med,2017,107(17):110-124.
[12]GreussingR,Hackl M,Charoentong P, .Identificationof microRNA-mRNA functional interactions in UVB-induced senescenceof humandiploidfibroblasts[J].BMCGenomics,2013,14:224.
[13]Bosch R,Philips N,Suárez-Pérez JA, .Mechanisms of photoaging and cutaneous photocarcinogenesis,and photoprotectivestrategieswithphytochemicals[J].Antioxidants(Basel),2015,4(2):248-268.
[14]Bhatia-DeyN,KanherkarRR,StairSE, .Cellular senescence as the causal nexus of aging[J].Front Genet,2016,7:13.
[15]陈亮,毕波,曾继平,等.罗格列酮调控光老化成纤维细胞周期阻滞的研究[J].中国美容整形外科杂志,2015,26(7):436-440.
[16]Karthikeyan R,Kanimozhi G,Prasad NR, .7-Hydroxycoumarin prevents UVB-induced activation of NF-B and subsequent overexpression of matrix metalloproteinases and inflammatorymarkersinhumandermal fibroblast cells[J].JPhotochem Photobiol B,2016,161:170-176.
[17]Lu J,Guo JH,Tu XL, .Tiron inhibits UVB-Induced AP-1 binding sites transcriptional activation on MMP-1 and MMP-3 promoters by MAPK signaling pathway in human dermal fibroblasts[J].PLoS One,2016,11(8):e0159998.
[18]Rubinsztein DC,Marino G,Kroemer G.Autophagy and aging[J].Cell,2011,146(5):682-695.
[19]Laplante M,Sabatini DM.mTOR signaling in growth control and disease[J].Cell,2012,149(2):274-293.
[20] Finkel T.Relief with rapamycin:mTOR inhibition protects against radiation-induced mucositis[J].Cell Stem Cell,2012,11(3):287-288.
[21] Sonis ST.Oral mucositis[J].Anticancer Drugs,2011,22(7):607-612.
