外源水杨酸对干旱胁迫下马铃薯生长及抗性生理指标的影响
2018-01-12姚岭柏韩海霞
姚岭柏, 任 琴, 韩海霞
(集宁师范学院生物系,内蒙古乌兰察布 012000)
马铃薯(SolanumtuberosumL.)为茄科茄属的草本植物,具有适应性强、生育期短的特点,在全世界栽培广泛,是重要的粮食、蔬菜和经济作物。马铃薯是典型温带植物,对水分和高温敏感[1-2]。中国马铃薯栽培面积达400万hm2,其中内蒙古是全国主要的马铃薯产区,内蒙古乌兰察布市有“中国马铃薯之都”的称号,该地区气候冷凉、光照充足、昼夜温差大、夏季与雨季重合,恰好适合马铃薯的生长发育[3]。所以,乌兰察布马铃薯产量高、品质好。但是,本地区的春季雨量少,春季内蒙古中西部降水多为10 mm以下,且近30年间的降水量呈波动下降趋势,易造成马铃薯幼苗期干旱,从而影响马铃薯植株叶面积增大,进而影响马铃薯产量与品质[3-6]。
水杨酸(salicylic acid,SA)是植物内源信号分子之一,能诱导植物对干旱、低温、病害、盐害胁迫的防御反应[7]。关于外源水杨酸能否缓解逆境植物伤害,已在干旱胁迫小麦、干旱胁迫黄瓜、盐胁迫菊芋等植物的逆境胁迫中发现,外源水杨酸起到一定的调控作用[8-11],另外外源水杨酸能增强马铃薯叶绿素合成,降低丙二醛的含量,提高抗氧化酶活性,从而显著降低马铃薯对疮痂病的感病率[12]。外源水杨酸还可在一定程度上使连作马铃薯的生长得到促进,叶片内渗透调节物质含量和抗氧化酶活性提高[13],但针对外源水杨酸对干旱马铃薯影响的相关研究未见报道。本研究在室内模拟干旱胁迫下对马铃薯叶片喷施水杨酸,探讨外源水杨酸对干旱胁迫下马铃薯抗性诱导的影响机制,以期为干旱地区马铃薯的生产应用提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 品种 马铃薯后旗红品种由乌兰察布市种子公司提供,为“薯都”乌兰察布市主栽品种。
1.1.2 试剂 水杨酸(SA),由天津市致远化学试剂有限公司提供。
1.2 试验方法
1.2.1 幼苗准备 试验于2016年3月在集宁师范学院植物园日光温室内进行。3月18日将后旗红马铃薯种薯(已催芽)切成50 g左右的小块,每块具1~2个芽,铺平晾干。采用16 cm×16 cm塑料盆进行盆栽试验。盆土由园土混合15%有机肥,再添加少量马铃薯专用复合肥混匀消毒装盆,每盆装等体积的土,播种深度5 cm,压实土,等量1次浇透水;设空白盆作为对照,共播种210盆。已测定盆土田间最大持水量(24.44%),在幼苗培养期间采用正常土壤含水量,盆土含水量控制在70%~80%,每天称量空白盆,确定浇水量。出苗后每盆留1株幼苗,当幼苗长到15 cm时进行处理。
1.2.2 干旱胁迫及处理 待马铃薯植株长到15 cm时,将马铃薯分为A、B等2组,A组(干旱前喷施)喷施不同浓度的水杨酸,之后进行干旱胁迫;B组(干旱后喷施)直接进行干旱胁迫,当马铃薯植株表现出萎蔫、老叶片黄化等干旱症状时,再进行水杨酸处理。干旱胁迫期间,均控制盆土含水量在35%~45%,以空白盆为参考确定浇水量。设SA浓度分别为0(CK)、6.9、13.8、27.6 mg/L,对照以清水喷施,各处理分别隔离喷施,直到叶片开始滴水为止。A、B组共8个处理,每次处理重复3次,每重复8盆。从B组喷施水杨酸开始取样,喷洒后0(喷后2 h)、10、20 d测定生长指标,并取同一部位真叶,测定生理指标。
1.2.3 指标测定 生长指标测定马铃薯最大叶长、叶面积、茎高,测定后计算10、20 d与0 d的指标变化值。取同部位功能叶片,测定叶片质膜透性。丙二醛MDA含量、游离脯氨酸含量、过氧化物酶POD活性、过氧化氢酶CAT活性、超氧化物歧化酶SOD活性、抗坏血酸过氧化物酶APX活性。参照张志良等的方法[14]进行叶片质膜透性,MDA和游离脯氨酸含量,POD、CAT、SOD活性的测定,参照高俊凤的方法[15]测定APX活性。其中,POD活性计算时以1 minD470 nm值变化0.01作为1个酶活性单位(U),SOD活性以抑制NBT光化还原的50%为1个酶活性单位(U),CAT以1 minD240 nm值下降0.1作为1个酶活性单位(U)。
1.3 数据处理
试验数据以Excel 2003处理图表,以SPSS 19.0处理系统对结果进行方差分析。
2 结果与分析
2.1 外源SA对干旱马铃薯生长指标的影响
由表1可见,与对照相比,A组方法(在干旱前施用外源SA)各处理浓度对干旱马铃薯的最大叶长、单株叶面积和茎高3个指标有不同趋势的影响。用13.8 mg/L SA进行A组处理后,除了10 d时的单株叶面积变化率(比对照低了0.3百分点,13.8 mg/L处理单株叶面积在10 d变化值÷13.8 mg/L 单株叶面积在0 d观测值×100%-CK单株叶面积10 d变化值÷CK在0 d观测值×100%)以外,对干旱马铃薯的最大叶长、20 d单株叶面积和茎高有显著(P<0.05)的改善作用,20 d时变化率分别较对照高29.0、13.4、13.2百分点。6.9 mg/L处理对最大叶长和 20 d 单株叶面积缩小有显著减缓作用外,对其余生长指标无显著影响。27.6 mg/L处理显著促进干旱马铃薯最大叶长,而对单株叶面积和茎高则较对照有显著抑制作用。
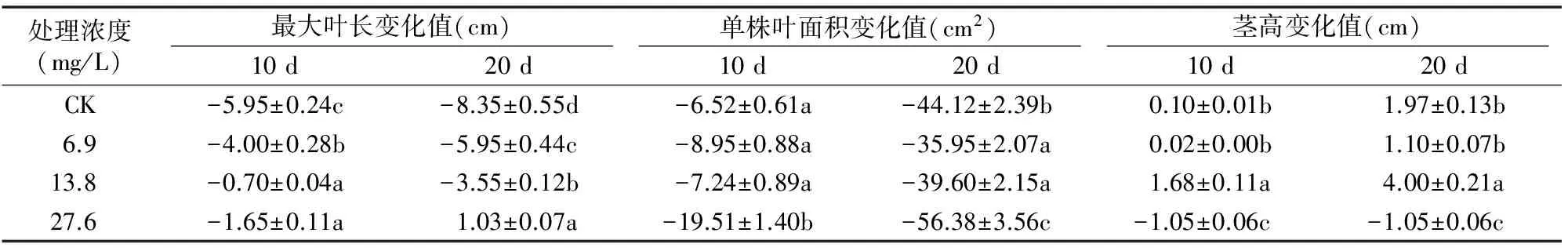
表1 外源SA处理下持续干旱马铃薯生长指标的变化(A组)
注:表中数据为对应时间数值与0 d数值之差±标准差。同列数据后不同小写字母代表在0.05水平上差异显著,下同。
由表2可见,与对照相比,B组的各个处理(干旱后施用外源SA)对干旱马铃薯的最大叶长均无显著影响(P>0.05),但对单株叶面积和茎高有不同程度的影响。B组SA处理浓度为13.8 mg/L时可显著减缓干旱马铃薯单株叶面积的缩小,在干旱10、20 d时单株叶面积变化率分别较对照多保持了17.78、23.21百分点;13.8 mg/L SA还促进了干旱马铃薯的茎高的增长,其中20 d时变化率与对照的差异达显著水平,提高20.9百分点。6.9~27.6 mg/L处理对单株叶面积的缩小有一定程度减缓作用,其中在20 d时与对照差异达显著水平,对茎高则无明显作用。27.6 mg/L处理对以上指标无明显改善作用。
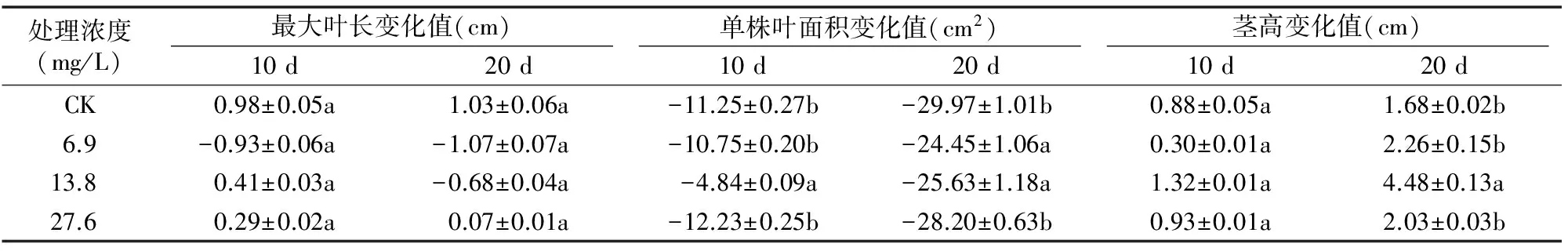
表2 外源SA处理下持续干旱马铃薯生长指标的变化(B组)
可见,A(干旱前处理)、B(干旱后处理)组随着处理浓度由6.9~27.6 mg/L不断增大,外源SA对干旱马铃薯的生长指标具有一定改善作用,其作用大小呈小、大、微小(B组)或加剧伤害(A组)的趋势,13.8 mg/L处理均表现最佳。但A、B组间存在一定差异,干旱胁迫相同时间情况下,A组较B组在最大叶长方面的改善作用更优,而在叶面积和茎高方面的改善作用则较差。另外,A、B组对比结果显示,处理浓度为 13.8 mg/L 时,B组单株叶面积变化率在10、20 d较对照分别提高了17.8、23.2百分点,而A组为-0.3、13.4百分点,说明外源SA浓度为13.8 mg/L时能够有效减缓马铃薯干旱胁迫的伤害,B组较A组方法能更大程度地保持干旱马铃薯单株叶面积,有利于增加光能利用率,从而有利于光合同化产物的积累。
2.2 外源SA对干旱马铃薯质膜透性变化、游离脯氨酸和MDA含量变化的影响
由图1可知,A、B组在随着干旱时间(10~20 d)的延长,各处理的膜质透性变化值均表现为上升趋势。与对照相比,A组干旱10 d各处理的膜质透性变化率下降了15.4~34.9百分点,20 d降低了33.9~39.1百分点,与对照差异均达到显著水平,A组各处理的膜质透性均受到抑制。B组各处理在10、20 d的膜质透性变化率均较对照有显著下降,分别下降了8.6~24.9、9.3~11.2百分点。
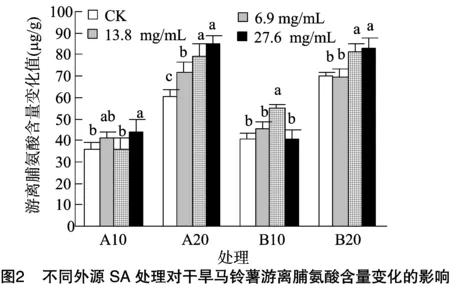
由图2可知,A、B组随着干旱时间的延长,各处理的游离脯氨酸含量呈上升趋势。A组处理10 d时,各处理游离脯氨酸含量变化值与对照差异不大,仅27.6 mg/L处理的增加量与对照差异达显著水平; 20 d时各处理变化值随浓度增加而依次增大,且均显著高于对照。B组处理10 d和20 d的浓度分别为13.8 mg/L和13.8、27.6 mg/L时,其脯氨酸含量变化值显著高于对照值,其余处理与对照无显著差异。说明无论A组还是B组方法,外源SA都可提高干旱马铃薯的游离脯氨酸含量的增幅,且20 d时的增幅大于10 d的。

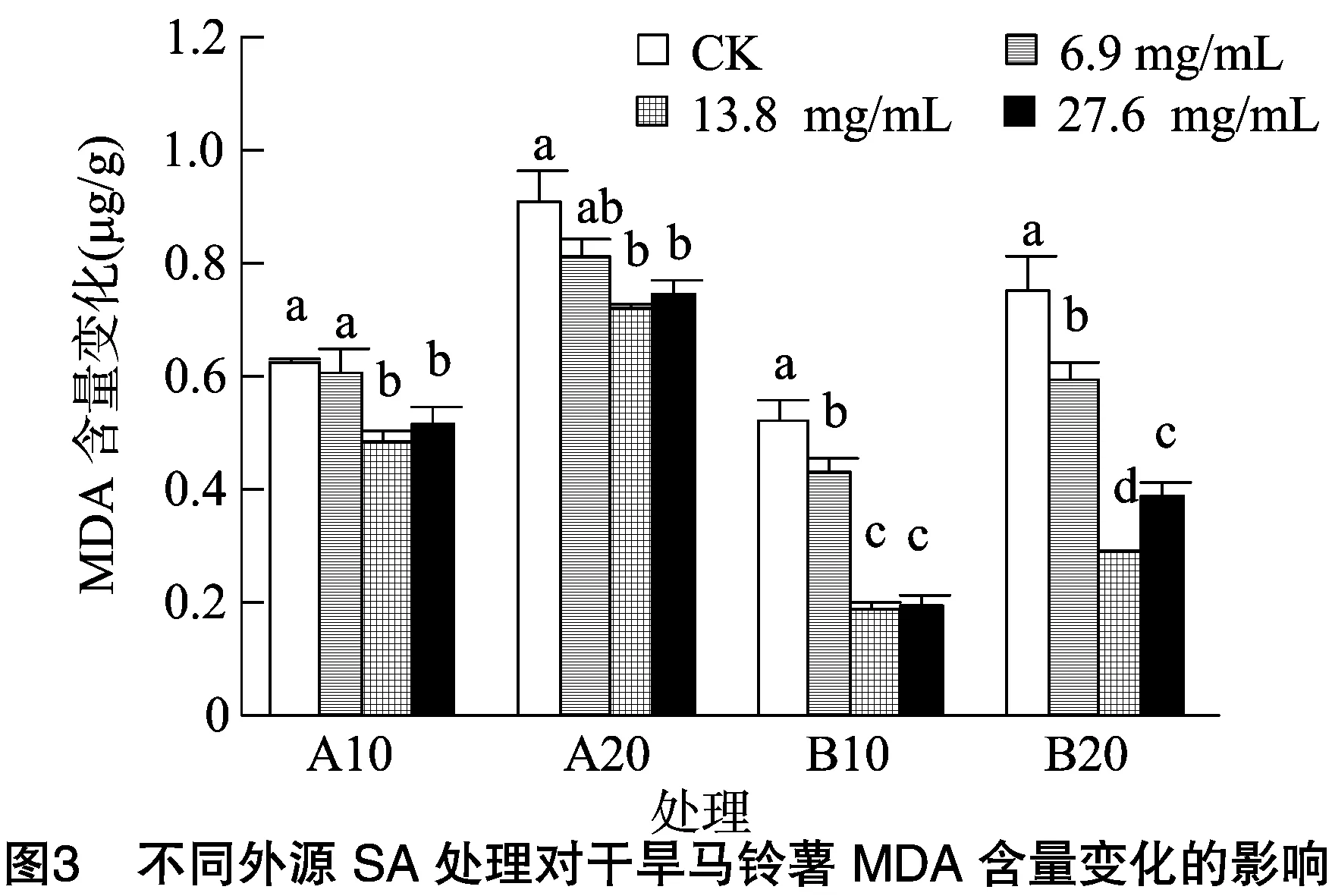
由图3可知,随着处理时间的延长,A、B组各处理MDA增加量呈上升趋势,与对照相比,各处理增加幅度均有不同程度的下降。A组中13.8、27.6 mg/L处理与对照的差异达显著水平(P<0.05),B组中6.9~27.6 mg/L处理与对照的差异达显著水平,且A、B组13.8 mg/L处理的增幅为最小。13.8 mg/L处理下,A组在10、20 d的MDA增量变化率分别较对照降低19.7、26.8百分点,B组降低了33.3、47.2百分点。说明外源SA可有效抑制干旱马铃薯MDA的积累,减轻了马铃薯细胞膜质过氧化程度,与A组相比,B组的改善效果更好。
2.3 外源SA对干旱马铃薯4种抗氧化酶活性的影响
由表3可知,A组干旱马铃薯外施SA后,4种抗氧化酶活性呈规律性变化,其在10、20 d的变化值与对照具有一定差异。POD是清除氧自由基的重要抗氧化酶之一,持续干旱后,POD活性变化值基本为负值,即活性下降。10 d时POD活性呈较大幅度下降,20 d时有小幅度的回升。其中, 27.6 mg/L 处理的变化值与对照差异达显著水平,有效抑制了POD活性的下降,POD活性变化率分别比对照下降了15.3、12.4百分点。
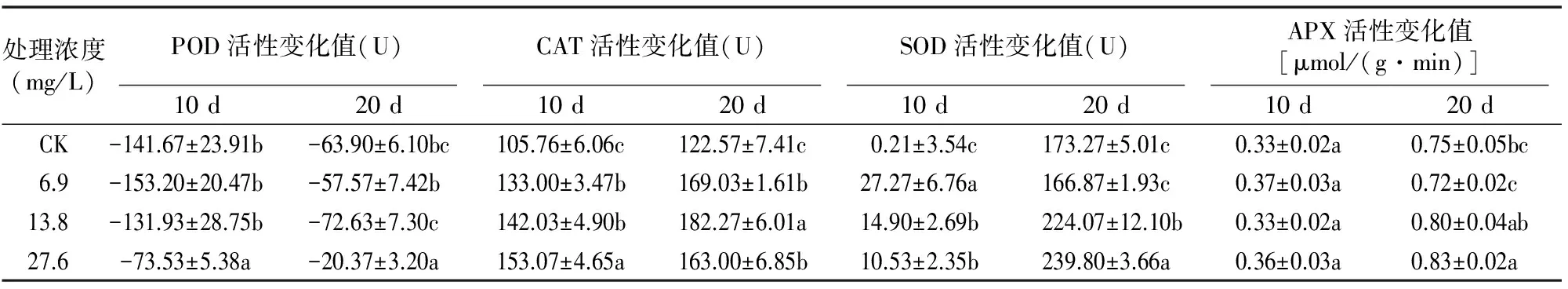
表3 外源SA处理下持续干旱马铃薯抗氧化酶活性的变化(A组)
CAT活性在持续干旱过程中变化值为正值,即活性上升,且10、20 d各个处理均与对照差异达显著水平,CAT活性变化率分别较对照提高了21.6~29.2、27.3~37.0百分点。说明外源SA能够使干旱马铃薯的CAT活性随着时间的延长稳定上升。
SOD活性的变化值均为正值,即持续干旱10~20 d SOD活性呈上升趋势。10 d时,6.9~27.6 mg/L SA均可提高干旱马铃薯的SOD活性的增量,与对照的差异达显著水平(P<0.05);20 d时,13.8~27.6 mg/L SA与对照的差异达显著水平。2次测得的最大变化率较对照分别提高109.5、398.1百分点。说明A组中外源SA能够大幅度提高干旱马铃薯的SOD活性。
APX活性变化值均为正值,即干旱期间APX活性上升,10 d时各处理变化率与对照相近,增加了4.0~5.6百分点;20 d时增幅升高,其中27.6 mg/L较对照变化率上升了10.2百分点,与对照差异达显著水平。
由表4可见,B组干旱马铃薯外施SA后,POD活性变化趋势与A组的变化趋势相似,在干旱胁迫后10、20 d先大幅度下降后小幅回升。同时,POD活性变化幅度(变化值的绝对值)随处理浓度升高而减小,且10 d时13.8~27.6 mg/L、20 d时6.9~27.6 mg/L与对照差异达显著水平,说明此浓度范围内的外源SA能有效减缓干旱马铃薯POD活性的下降。
CAT活性变化值为正,即活性上升,各处理增加幅度均大于对照,且10 d时6.9 mg/L、20 d时6.9~13.8 mg/L与对照差异达显著水平。
SOD活性变化量均为正值,即干旱期间马铃薯中SOD活性呈上升趋势变化,在10 d时6.9~27.6 mg/L处理马铃薯中的SOD活性增加量较对照显著下降;20 d时6.9、27.6 mg/L处理SOD活性增加量与对照无显著差异, 13.8 mg/L 增加量较对照显著上升。说明外源SA在10 d时显著抑制了SOD活性的上升,到20 d时此抑制作用消失,其中13.8 mg/L SA可显著提高SOD的增加幅度。

表4 外源SA处理下持续干旱马铃薯抗氧化酶活性的变化(B组)
APX活性也呈上升变化,10 d时各处理的APX活性增加率高于对照24.1~51.9百分点,均达到显著水平;20 d时,增加率较对照上升至26.1~54.8百分点,达到显著水平。说明干旱后(B组)外施SA能够显著提高马铃薯APX活性。
综上所述,持续干旱10、20 d马铃薯体内POD活性下降,CAT、SOD和APX等3种酶活性上升,干旱前(A组)和干旱后(B组)施加外源SA对POD和CAT活性的影响相近,外源SA均能够在一定程度上促进酶活性升高,但对另2种酶则有不同影响:A组可大幅度促进马铃薯的SOD活性,10 d最高增幅比对照提高了18.86倍,而B组处理在10 d时抑制了SOD活性的升高,到20 d时抑制作用消失;A组处理对干旱马铃薯APX活性的影响不大,B组处理有效促进了APX活性的升高,10 d时比对照的增加率高了24.1~51.9百分点。表明外施SA后能够减缓POD活性的下降趋势,有效促进CAT活性的增加量,干旱前外施SA有利于SOD活性的提高,干旱后外施SA有利于APX活性的提高,且干旱后施用酶活性提高幅度较大。
说明干旱前、后SA处理对马铃薯防御干旱的4种抗氧化酶活性有不同的影响。干旱前施用SA,主要激活了SOD的活性,其次是CAT,马铃薯体内按抗氧化酶活性增加量由大到小依次为SOD、CAT、APX、POD;干旱后施用SA,主要激活了APX和CAT,马铃薯体内按抗氧化酶活性增加量由大到小依次为APX、CAT、SOD、POD。这几种抗氧化酶一起清除氧自由基,减轻干旱胁迫对马铃薯的伤害,且干旱后的施用效果较好。
3 结论与讨论
外源SA的浓度在6.9~13.8 mg/L时能够不同程度缓解干旱对马铃薯生长的胁迫伤害,以13.8 mg/L为最佳处理浓度,而27.6 mg/L则只有微弱的缓解作用(B组)或者加剧胁迫伤害(A组)。结果表明,外源SA处理干旱马铃薯浓度不宜超过13.8 mg/L,且干旱后处理较干旱前处理效果更好。在干旱前后进行SA处理,均能够显著抑制干旱胁迫下马铃薯叶片膜质透性的增大,促进游离脯氨酸含量的增加,抑制MDA含量的增加,其中干旱后处理(B组)对MDA的抑制效果更好,且以13.8 mg/L处理最佳。马铃薯叶片中抗氧化酶活性与其抗旱有关,POD活性在干旱后呈下降趋势变化,干旱后以外源SA处理能够在中高浓度下有效抑制这种下降,在外源SA诱导下SOD、APX活性出现大幅度上升,而CAT活性则随胁迫时间平稳上升,与SA处理浓度无明显相关性。
逆境胁迫导致植物生长受抑制、体内大量积累超氧阴离子和H2O2[1-2,6],植物会启动调控反应,如调节细胞渗透势、增强抗氧化酶活性等。其中,游离脯氨酸是植物内调节细胞渗透势的主要物质之一,SOD能够清除氧自由基,POD和CAT能够清除过量H2O2,APX可以特异性地催化抗坏血酸与H2O2反应,起到清除H2O2的目的。本试验中外源SA对干旱胁迫下马铃薯的生长和生理指标均有一定的调控作用。
关于外源SA喷施浓度的研究结果与王磊等关于水杨酸在菊芋上的研究结果[10]一致,同时与杨伟等关于青花菜的研究结果[16]相近,但比回振龙等在连作马铃薯上喷施SA最佳浓度(20~30 μmol/L)大一些[13],可能是本试验只处理了1次而回振龙试验中处理3次的原因。本试验关于外源SA对提高植物生理方面抗性能力的研究结果,与前人的研究结果[7-17]一致,表明外源水杨酸在干旱胁迫下小麦、黄瓜、青花菜、高羊茅等植物的生长和生理指标方面,均具有不同程度的调控作用[7-9,16-17],SA具有增强植物抗旱性方面的积极作用。还有研究表明,外源SA在增强马铃薯抵抗疮痂病能力和克服马铃薯连作障碍方面也具有一定的作用[12-13],也有研究显示外源SA可在增强菊芋抗盐性、增强萝卜克服高温、铅胁迫能力方面具有一定的作用[10-11,18]。所以,外源水杨酸能够有效增强后旗红马铃薯的抗旱能力。
[1]Belanger G,Walsh J R,Richards J E,et al. Tuber growth and biomass partitioning of two potato cultivars grown under different N fertilization rates with and without irrigation[J]. Amer J of Potato Res,2001,78(2):109-117.
[2]抗艳红,龚学臣,赵海超,等. 不同生育时期干旱胁迫对马铃薯生理生化指标的影响[J]. 中国农学通报,2011,27(15):97-101.
[3]苏立娟. 内蒙古地区云、降水和可降水量的时空分布、变化趋势及其与气候因子的关系[D]. 兰州:兰州大学,2005:41-43.
[4]孙雪平,苏 筠. 降水约束条件下的粮食生产适应度评价——以内蒙古乌兰察布市为例[J]. 干旱地区农业研究,2016,34(3):234-239.
[5]海梅荣,陈 勇,周 平,等. 干旱胁迫对马铃薯品种生理特性的影响[J]. 中国马铃薯,2014,28(4):199-204.
[6]张丽莉,王庆祥,石 瑛,等. 干旱胁迫对马铃薯叶肉和茎部细胞超微结构的影响[J]. 沈阳农业大学学报,2015,46(1):91-95.
[7]孟雪娇,邸 昆,丁国华. 水杨酸在植物体内的生理作用研究进展[J]. 中国农学通报,2010,26(15):207-214.
[8]单长卷,赵新亮,汤菊香. 水杨酸对干旱胁迫下小麦幼苗抗氧化特性的影响[J]. 麦类作物学报,2014,34(1):91-95.
[9]郝敬虹,易 旸,尚庆茂,等. 水杨酸处理对干旱胁迫下黄瓜幼苗氮素同化及其关键酶活性的影响[J]. 园艺学报,2012,39(1):81-90.
[10]王 磊,隆小华,孟宪法,等. 水杨酸对NaCl胁迫下菊芋幼苗光合作用及离子吸收的影响[J]. 生态学杂志,2011,30(9):1901-1907.
[11]初 敏. 水杨酸和茉莉酸对高温胁迫下萝卜幼苗的缓解效应[D]. 泰安:山东农业大学,2012:19-23.
[12]汤晓莉,薛红芬,邓国宾,等. 水杨酸诱导马铃薯疮痂病抗性的生理机制研究[J]. 西南农业学报,2010,23(6):1851-1854.
[13]回振龙,王 蒂,李宗国,等. 外源水杨酸对连作马铃薯生长发育及抗性生理的影响[J]. 干旱地区农业研究,2014,32(4):1-8.
[14]张志良,瞿伟菁,李小方. 植物生理学实验指导[M]. 4版. 北京:高等教育出版社,2009.
[15]高俊凤. 植物生理学实验技术[M]. 北京:高等教育出版社,2006:211-217.
[16]杨 伟,张国斌,肖雪梅,等. 水杨酸对不同灌水下限青菜花根系生长、根冠比及叶片抗氧化酶活性的影响[J]. 干旱地区农业研究,2015,33(3):148-152.
[17]赵春旭,刘芳芳,赵 伟,等. 水杨酸浸种对高羊茅在干旱胁迫下萌发的影响[J]. 草业科学,2011,28(11):1945-1949.
[18]杜连彩. 水杨酸对铅胁迫下潍县萝卜抗氧化酶活性的影响[J]. 安徽农业科学,2012,40(2):828-829,877.
