北京地区莴苣中番茄斑萎病毒的鉴定
2018-01-11陈东亮李明远李雪梅王玲玲4黄丛林
陈东亮李明远李雪梅王玲玲,4黄丛林*
(1北京市农林科学院,北京农业生物技术研究中心,北京 100097;2北京市功能花卉工程技术研究中心,北京 100097;3农业基因资源与生物技术北京市重点实验室,北京100097;4西藏农牧学院,西藏林芝860000)
莴苣(Lactuca sativaL.)是一种世界性的蔬菜,也是我国最常见的蔬菜种类之一,在我国南北各地广泛栽植,是一种非常重要的经济作物。病毒病是莴苣生产过程中常见的病害,由于其发病快、防治难,往往造成巨大损失。目前已知有十余种植物病毒可以为害莴苣(Moreno & Fereres,2012;Ciuffo et al.,2016),其中番茄斑萎病毒(Tomato spotted wilt virus,TSWV)由于传播范围广、防治困难而日益成为莴苣生产中危害严重的病害之一。
TSWV为布尼亚病毒科(Bunyaviridae)番茄斑萎病毒属(Tospovirus),已知至少可以感染84个科的1 090种植物,是目前已知的寄主范围最广的植物病毒之一(Parrella et al.,2003)。TSWV危害巨大,对一些农作物甚至会造成毁灭性的损失,是辣椒(Capsicum annuum)、番茄(Lycopersicon esculentum)、 烟 草(Nicotiana tabacum)、 菊 花(Chrysanthemum morifolium)等经济作物重点防疫的病害。自1915年在澳大利亚发现至今,TSWV已传播到了全球大部分国家,成为一种全球性的植物病害。
1986年,首次在我国境内发现TSWV(许泽永 等,1986),随后的十几年其传播范围主要集中于云南、四川及广东部分地区(苏大昆 等,1987;许泽永 等,1989;姚革,1992),并未发展为全国性的病害。与此同时,我国也将TSWV列入了《进境植物检疫性有害生物名录》,加强了入境植物该病毒的检疫防疫工作,有效防止了TSWV进一步的扩散。但近几年,伴随国外引种和国内资源的流通,TSWV扩散速度加快,天津、北京、山东、海南等地均报道发现TSWV(李飞 等,2012;邱树亮 等,2012;高苇 等,2016;车海彦 等,2017;李洁 等,2017)。
2017年3月,笔者在北京市顺义区发现温室内栽培的莴苣疑似发生了严重的病毒病。经RT-PCR检测、序列测定与生物信息学分析,证实其病原物为TSWV。进一步基于TSWV核壳体(nucleocapsid protein,NP)基因序列构建了国内外不同地区TSWV的系统进化树,分析了不同国家、不同地区间TSWV的进化关系,为进一步研究TSWV传播机制、制定防控措施奠定了基础。
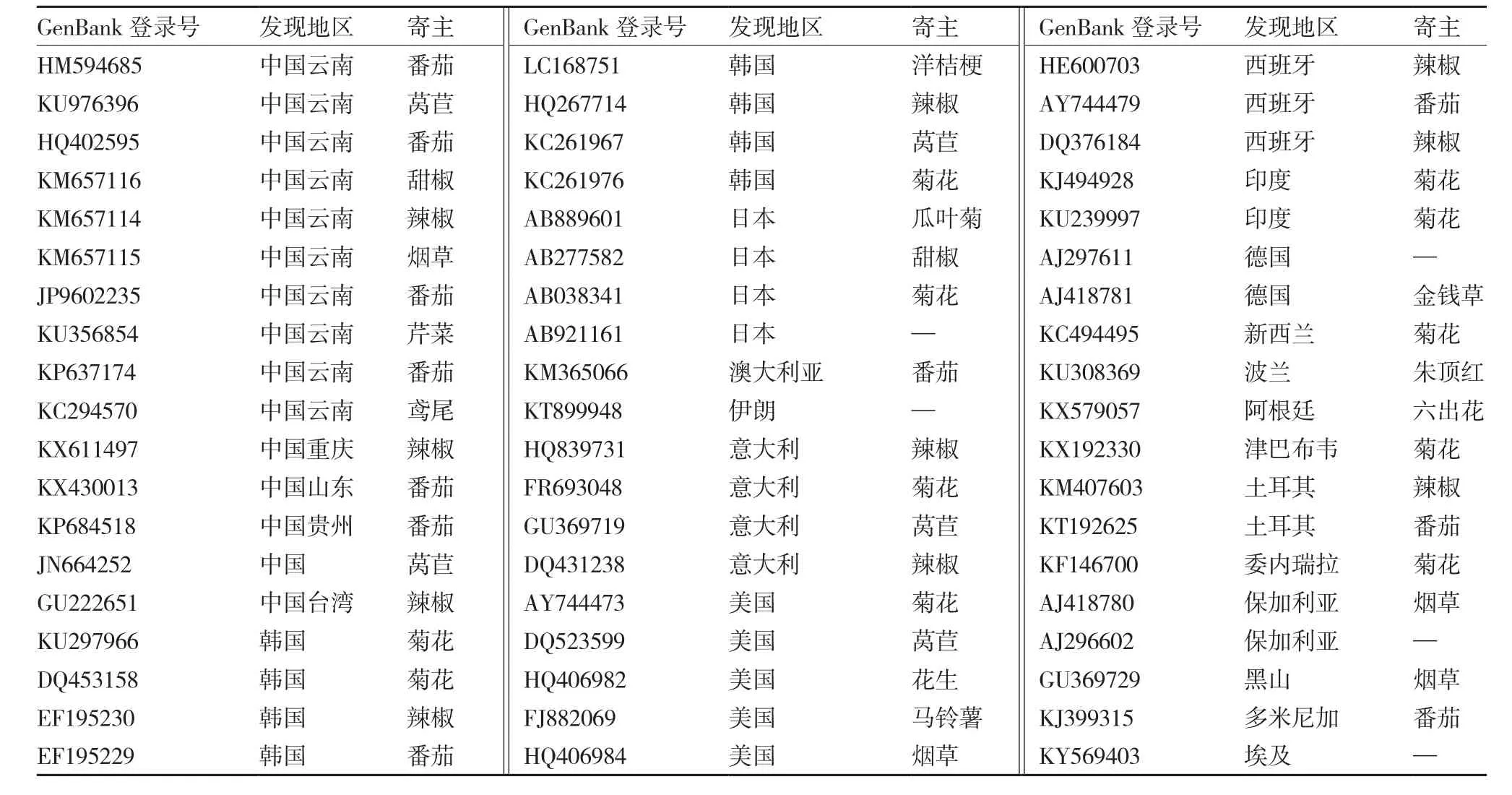
表1 构建系统进化树使用的TSWV NP基因序列信息
1 材料与方法
1.1 样品采集及蓟马的检测
2017年4月,于北京市顺义区北郎中村种植莴苣的温室内采集具有叶片黄化褪绿、枯死、矮化等症状的莴苣叶片,用干净的保鲜袋包裹后放置于冰盒中,带回北京农业生物技术研究中心实验室,于-80 ℃保存备用。同时,收集病害温室带有蓟马的莴苣叶片,装入保鲜袋后带回实验室,于显微镜下观察蓟马虫体特征。
1.2 TSWV特异序列的克隆
病样总RNA的提取使用全式金RNA提取试剂盒。采用Promega反转录试剂盒制备样品cDNA。利用Primer STAR高保真酶(Takara)进行TSWVNP基因特异序列的扩增,引物设计和RT-PCR反应程序参考Chung等(2006)的方法。扩增产物经1%琼脂糖凝胶电泳检测后,纯化回收并连接于克隆载体pGEM-T easy(Promega,USA),热激转化大肠杆菌DH5α后,挑取阳性克隆送生工生物工程(上海)股份有限公司测序。
1.3 序列分析与进化树的构建
利用DNASTAR软件进行序列的拼接和初步处理。利用MEGA 4软件,采用邻接法(neighbour joining)构建不同来源TSWVNP基因的系统进化树。系统进化树构建使用的其他TSWVNP序列均下载自美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI) 数 据 库, 序列GenBank登录号、发生地区、寄主等信息如表1所示。
2 结果与分析
2.1 病害调查
感病莴苣主要表现为:叶片斑驳褪绿,植株矮化,生长缓慢,部分肉质茎部有明显条状黑斑,严重时整株死亡(图1)。这与最近云南地区报道的TSWV引起的莴苣类蔬菜病害特征类似(郑宽瑜等,2015)。经初步统计,温室内具有明显褪绿发黄、矮化等病症的莴苣植株达到25%以上。与此同时,温室内伴有蓟马的大量发生,经显微镜下观察鉴定为西花蓟马(Frankliniella occidentalis)(图2)。西花蓟马为入侵物种,近几年在国内多地大面积暴发,造成严重损失,同时西花蓟马还是TSWV最主要的传播介质。基于病害特征和西花蓟马的发生,初步推测此次发现的莴苣病害是由TSWV引起的病毒病。

图1 感病莴苣病害症状

图2 莴苣上发现的西花蓟马
2.2 RT-PCR检测
利用TSWV特异引物,对发病温室采集的7个表型明显的莴苣样本进行RT-PCR检测。PCR产物经电泳检测(图3),7个样本均在预期位置出现了清晰的条带;以无菌水为模板的阴性对照未出现该条带,表明这7个样品均受到TSWV的侵染。
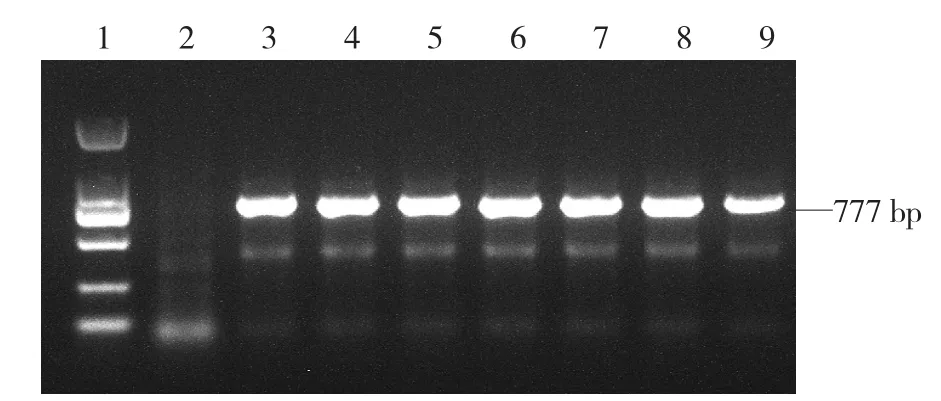
图3 TSWV的RT-PCR检测结果
PCR扩增产物经回收纯化,连接到克隆载体,转化大肠杆菌后,每个样本取3个克隆进行测序。共得到2条大小为777 bp的高度保守序列(BLZ01和BLZ02),2条序列间存在2个碱基的差异。NCBI BLASTN分析显示,这2条序列与已报道的TSWVNP基因序列高度相似,一致性高达98%~99%,其中与西班牙分离的TSWVNP基因(GenBank登录号:FR693152)一致性最高,仅相差2~4个碱基。将这2个基因序列在NCBI进行登录,登录号分别为MF139144和MF139145。RTPCR检测和测序结果进一步证明了本次发现的莴苣病害的病原为TSWV。
2.3 不同来源TSWV的遗传多样性分析
基于NP基因序列,构建本试验分离的莴苣TSWV与国内外其他地区已知TSWV的系统进化树。由图4可知,国内已报道的其他地区分离的TSWV明显聚集在一起,并进一步与日本、韩国的部分TSWV株系聚合在一起,表明中国早期TSWV可能由日本、韩国等地传入,之后在云南、重庆等地逐渐扩散。本试验中分离鉴定的莴苣TSWV与国内已报道株系差异较大,而与西班牙等国外株系聚合较紧密,表明本次发现的莴苣TSWV并非由云南、重庆等国内地区传入,很有可能是由国外直接传入北京的。
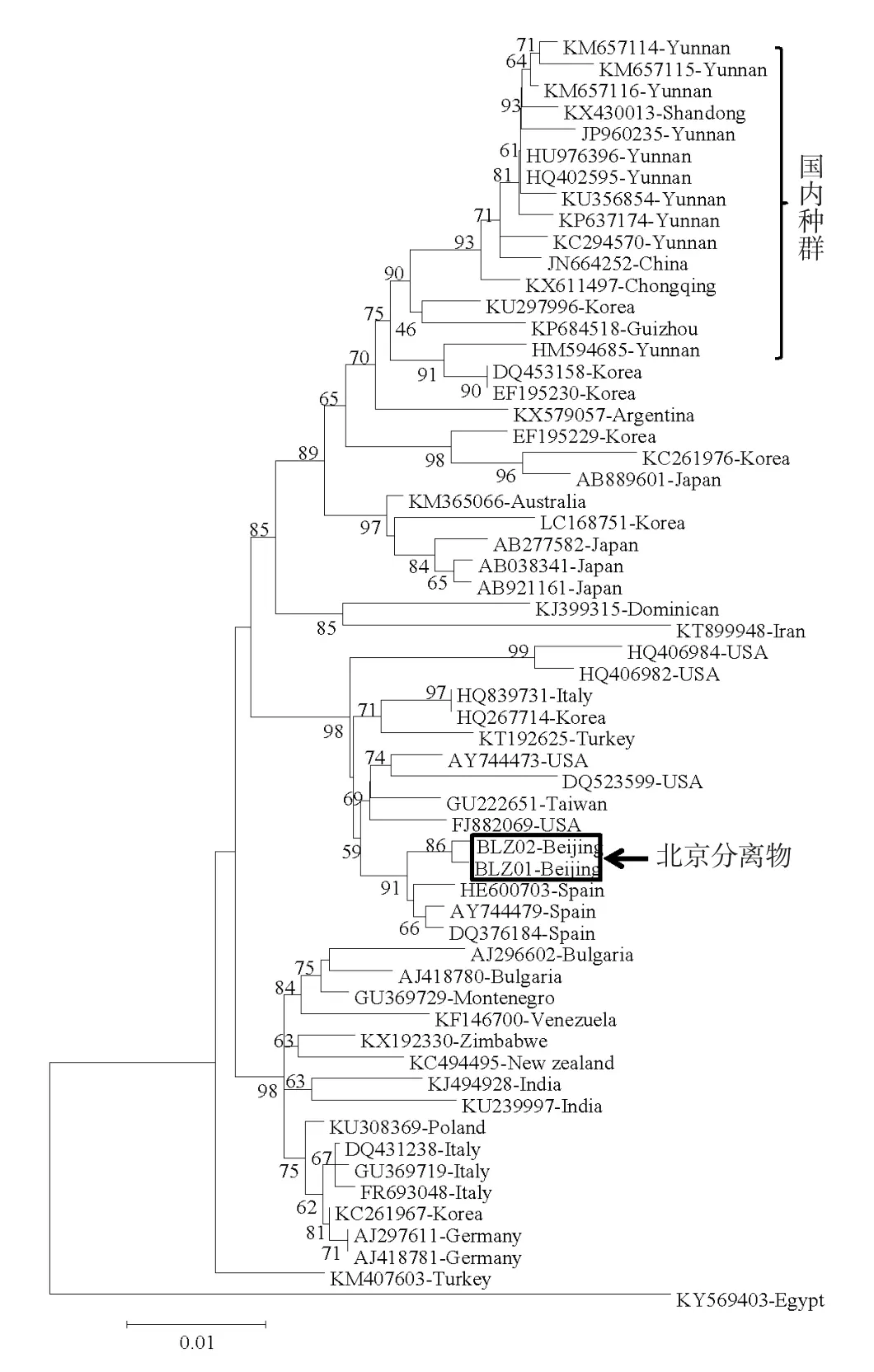
图4 基于NP基因序列的TSWV系统进化树
3 结论与讨论
TSWV寄主范围广泛,甚至能侵染一些常见的田间杂草,从而使得病源清除困难、传播速度快、防治困难。自1915年首次发现,TSWV已扩散到了全球大部分国家和地区,是名副其实的全球性植物病害,被列为世界十大植物病害之一。TSWV主要通过蓟马传播,蓟马生活周期短、繁殖速度快、寄主范围广、适宜性强、极易形成抗药性,防治难度较大。其中,西花蓟马是TSWV最高效的传播媒介。西花蓟马在我国也属于国外入侵的有害生物,同时也是我国入境检疫的有害生物,国内最早关于西花蓟马的报道见诸于2003年(张友军 等,2003)。近几年,西花蓟马在我国大面积暴发,已成为部分地区常见的虫害(张友军 等,2011)。西花蓟马的扩散也加速了TSWV的传播,本试验中发现TSWV的温室及其周边长期存在蓟马为害,经显微镜下观察确定为入侵物种西花蓟马,西花蓟马可能是本次TSWV暴发的主要媒介。
病毒病一旦感染极难防治。为了及时发现、尽早防治,研究人员开发了多种针对TSWV快速检测的方法,比如酶联免疫法(Sherwood et al.,1989)、电镜检测法(张仲凯 等,2004)、LAMP 法(Wu et al.,2016)、RT-PCR 法(Pappu et al.,1996)等。其中,RT-PCR法由于操作简便、灵敏度高、特异性强、检测速度快、成本低等特点而被广泛应用于植物病毒的快速检测。本试验结合病症特征和RTPCR检测,确诊了是由TSWV引起的北京顺义莴苣病害。同时,基于TSWV NP基因序列,对国内外TSWV的遗传多样性进行分析,从系统进化树可以看出,发生在我国云南、广州等地的TSWV亲缘关系都比较近,且与韩国、日本等国的部分株系表现出较近的关系,表明我国南方、西南地区的TSWV可能最早是由韩国、日本等地传入的。这与20世纪80年代、90年代昆明、广州等地大力发展花卉种植产业,自日本、韩国引进优良花卉品种的历史背景相符合。本试验中分离的TSWV与我国已知的TSWV亲缘关系较远,而与西班牙等国外报道的株系在进化关系上较为接近,表明本次发现的TSWV极可能是在引种过程中直接从国外带入的新病毒株系。近几年,国内外品种交流愈加频繁,也增大了有害生物随之带入的风险。本试验在北京地区首次报道TSWV为害重要经济作物莴苣,也为进一步加强TSWV的检疫、防疫工作敲响了警钟。
车海彦,曹学仁,刘培培,罗大全.2017.海南省冬季蔬菜病毒病发生情况调查.热带农业科学,37(1):71-74.
高苇,王勇,张春祥,张安盛,竺晓平.2016.天津地区辣椒病毒病调查及毒源种类初步鉴定.山东农业科学,48(3):91-94.
李飞,吴青君,徐宝云,谢文,王少丽,张友军.2012.北京地区发现番茄斑萎病毒.植物保护,38(6),186-188.
李洁,迟胜起,杨勤民,丁天波,褚栋.2017.山东烟台地区发生番茄斑萎病毒病危害.植物保护,43(1):228-232.
邱树亮,王孝宣,杜永臣,高建昌,国艳梅,朱德蔚,胡鸿,李宝聚,石延霞.2012.番茄斑萎病毒TSWV的鉴定及抗病种质的筛选.园艺学报,39(6):1107-1114.
苏大昆,袁宣泽,谢永红,王淑荣,丁海.1987.成都、渡口两市番茄中检出番茄斑萎病毒.植物病理学报,17(4):255-256.
许泽永,Reddy D V R,Rajeshwari R,张宗义,黄立新.1986.我国南方花生发生一种由番茄斑萎病毒引起的新病害.病毒学报,2(3):80-83.
许泽永,张宗义,陈金香.1989.番茄斑萎病毒(TSWV)广州分离物生物学特性研究.植物病理学报,19(4):198.
姚革.1992.四川晒烟上发现番茄斑萎病毒(TSWV).烟草科技,(6):2-4.
张友军,吴青君,徐宝云,朱国仁.2003.危险性外来入侵生物——西花蓟马在北京发生危害.植物保护,29(4):58-59.
张友军,朱国仁,褚栋,吴青君,王少丽.2011.我国蔬菜作物重大入侵害虫发生、危害与控制.植物保护,37(6):1-6.
张仲凯,杨录明,方琦,彭潞波,汪继玲,普耀芳.2004.番茄斑萎病毒侵染烟草的细胞病理.电子显微学报,23(4):349.
郑宽瑜,吴阔,董家红,方琦,张仲凯.2015.番茄斑萎病毒对云南莴苣类蔬菜的侵染危害.植物保护,41(5):174-178.
Chung B N,Pak H S,Jung J A,Kim J S.2006.Occurrence ofTomato spotted wilt virusin Chrysanthemum(Dendranthema grandiflorum)in Korea.Plant Pathology Journal,22(3):230-234.
Ciuffo M,Mammella M,Vallino M,Caciagli P,Turina M.2016.Molecular identification and biological characterization of a new potyvirus in lettuce.Archives of Virology,161(9):2549-2664.
Moreno A,Fereres A.2012.Virus diseases in lettuce in the Mediterranean basin.Advances in Virus Research,84:247-288.
Pappu H R,Culbreath A K,Bertrand P F,Csinos A S,Niblett C L.1996.Sequence analysis of the nucleocapsid protein gene of aTomato spotted wilt virusisolate from Georgia,USA.Acta Horticulturae,431:237-243.
Parrella G,Gognalons P,Gebre-Selass iè K,Vovlas C,Marchoux G.2003.An update of the host range ofTomato spotted wilt virus.Journal of Plant Pathology,85(4):227-264.
Sherwood J L,Sandbom M R,Keyser G C,Myers L D.1989.Use of monoclonal antibodies in detection ofTomato spotted wilt virus.Phytopathol,79(1):61-64.
Wu X,Chen C,Xiao X,Deng M J.2016.Development of reverse transcription thermostable helicase-dependent DNA amplification for the detection ofTomato spotted wilt virus.Journal of Aoac International,99(6):1596-1599.
