博斯腾湖北岸不同地下水埋深对塔干柽柳光合特性的影响
2017-12-29党亚玲韩炜马霄华管文轲费兵强崔凯强张娅
党亚玲, 韩炜, 马霄华, 管文轲, 费兵强, 崔凯强, 张娅
博斯腾湖北岸不同地下水埋深对塔干柽柳光合特性的影响
党亚玲1,2,3, 韩炜1,2,*, 马霄华1,2, 管文轲3, 费兵强1,2, 崔凯强1,2, 张娅1,2
1. 新疆师范大学地理科学与旅游学院, 乌鲁木齐 830054 2. 新疆干旱区湖泊环境与资源重点实验室, 新疆师范大学, 乌鲁木齐 830054 3. 新疆林业科学研究院, 新疆乌鲁木齐 830054
选择博斯腾湖北岸优势种塔干柽柳为研究对象, 对不同地下水埋深处其光合特性进行对比研究, 探究塔干柽柳对不同地下水埋深的响应和适应机制。在垂直湖岸线的方向上, 选取地下水埋深为0 m, 1.25 m和2.25 m的样地, 选择健康的3—5龄塔干柽柳作为测量对象, 用Li-6400光合作用仪测定其光响应过程。结果如下: (1)当>800 μmol·m–2·s–1时, 对应相同的光合有效辐射, 不同地下水埋深处塔干柽柳的净光合速率()的值的表现为:P地下水埋深2.25m>P地下水埋深1.25m>P地下水埋深0m; (2)地下水埋深2.25 m处, 塔干柽柳的净光合速率定积分值最高, 由非直角双曲线拟合得到的饱和净光合速率值最大, 暗呼吸速率最大, 光饱和点最高, 光补偿点最低; (3)当>1000 μmol·m–2·s–1时, 地下水埋深2.25 m处, 塔干柽柳的胞间浓度CO2()和蒸腾速率()随光合有效辐射的增加而增加, 地下水埋深0 m和1.25 m处呈平稳的变化趋势; (4)当<800 μmol·m–2·s–1, 水分利用效率()光响应三条曲线整体值大致表现为: 地下水埋深2.25 m>地下水埋深1.25 m>地下水埋深0 m。得出如下结论: 地下水埋深2.25 m处塔干柽柳的光合特征参数以及水分利用效率等各项生理指标均表现最佳, 博斯腾湖北岸水埋深约2.25 m处适宜塔干柽柳生长。
博斯腾湖; 地下水埋深; 塔干柽柳; 光合特性
1 引言
博斯腾湖湖滨带属于内陆干旱区特殊的生态系统带, 其生态系统的健康发育, 对保持湖滨带及其更大范围的区域生态安全具有重要的保障性意义。塔干柽柳(M. T. Liu)是博斯腾湖滨带的优势种, 其最适宜环境的探寻对其本身的健康发育具有指导性意义。
近年来博斯腾湖地区地下水位波动频繁, 导致其发育受到影响[1]。地下水埋深对植物的光合生理特征有显著的影响[2], 为了量化其影响程度并探明其机制, 许多学者对其做了大量研究。Horton 等人发现地下水埋深的变化对柽柳的生长与分布格局产生较大的影响[3–5]。许皓等人发现缓和的地下水位波动对柽柳现有的碳/水平衡影响不大,地下水位剧烈下降将危及多枝柽柳的生存[6]。陈敏等人发现柽柳叶片各项生理指标与地下水埋深存在着显著相关关系[7]。肖生春等人发现区域降水的差异是造成柽柳最大生态水位差异的主要原因[8]。张佩等人发现不同的地下水埋深, 叶片生理生态指标对不同地下水位呈现出明显差异, 在黑河中游绿洲荒漠过渡带柽柳适宜生存的地下水位在3 m左右[9]。也有研究发现当地下水埋深大于2.5 m时, 柽柳植株顶梢出现了枯死现象[10], 柴宝峰等人发现在受到干旱胁迫时柽柳渗透调节能力随季节而变化, 抗旱性和渗透调节能力随着枝条的木质化和叶子的革质化程度而增强[11]。邓雄等人发现柽柳叶片的气体交换对环境变化的响应能揭示柽柳对生境的内在适应机制, 柽柳属于湿生植物, 适应生境很广[12]。王会提等人的研究表明不同种植模式下柽柳的光合特性表现出不同的响应特征[13], 这些成果为研究内陆干旱区柽柳发挥的生态意义提供了一定的参考价值, 但有关博斯腾湖滨柽柳适宜的地下水埋深探寻的研究尚未见报道。
本文通过测定博斯腾湖北岸不同地下水埋深处柽柳叶片的光合特征参数, 探究博斯腾湖滨塔干柽柳对不同地下水埋深的响应规律, 旨在为这一区域植被恢复及柽柳植物群落的生长和保护提供基础资料, 为保护博斯腾湖湿地生态系统的稳定提供科学依据。
1 材料与方法
1.1 研究区概况
博斯腾湖(86°40′—87°56′E, 41°56′—42°14′N)位于新疆巴音郭楞蒙古自治州境内, 地处塔里木盆地北部, 是我国最大的内陆淡水湖泊[14]。四周高山环抱, 海洋气候微弱, 降水稀少, 蒸发旺盛, 气候干旱, 为典型的温带荒漠气候。湖区多年平均降水量为77.23 mm, 年蒸发量为2241.48 mm, 年均气温7.9 ℃, 7月平均气温22.8 ℃, 1月平均气温–8.1℃。博斯腾湖西岸以农田土和沼泽土为主, 南岸和北岸以沙土为主。分布于湖区西到西北沿岸的芦苇沼泽构成了湖滨湿地, 面积约为280 km2, 湖滨植物主要有芦苇()、塔干柽柳(、盐生草()等。
1.2 数据采集与处理
1.2.1 实验设计
距离湖岸线远近不同, 柽柳生长的地下水位不同, 据此沿博斯腾湖北岸(41°45′—42°15′N, 86°00′—86°26′E)垂直湖岸线方向0 m、100 m、200 m设置3个样带, 并在三个样带内选取长势良好的柽柳植被各设立两个5 m×5 m的植物样方, 并在每个样地布设五个监测断面, 在不同断面又设有多口观测井位观测地下水埋深。在测量期间测得不同样地地下水埋深分别为0 m (0.082±0.029) m、1.25 m (1.233±0.908) m、2.25 m(2.268±0.501) m。三个样带内, 所选择的多年生塔干柽柳的冠幅分别为0 m(P52.35±2.029) cm、1.25 m(P116.32±1.908) cm、2.25 m(P161.36± 2.501) cm。
实验于2016年9月16日—2016年10月15日进行, 这一时期柽柳的生物积累量基本达全年最大。选择典型晴天, 在9:00—12:30(北京时间)外界条件比较稳定时, 选择样方内3株健康生长的柽柳植株, 对向阳无遮光部位的叶片进行标定, 每簇标定的叶片重复测定三次。用便携式光合仪Li-6400- 2B红蓝光源设定模拟光辐射, 光辐射强度(R)的梯度设置为: 1800, 1600, 1400, 1200, 1000, 750, 600, 400, 200, 150, 50, 20, 0 μmol·m–2·s–1。
直接输出参数: 净光合速率(, μmol·m–2·s–1); 胞间CO2浓度(, μmol·mol–1)、气孔导度(, mmol·m–2·s–1)、蒸腾速率(, mmol·m–2·s–1)等光合生理参数。光合有效辐射(, μmol·m–2·s–1)、大气温度(, ℃)、大气CO2浓度(, μmol·mol–1)、大气相对湿度(, %)等环境因子参数在输出光合参数的同时同步输出。
1.2.2 数据处理
用光合助手软件(Photosynthetic Assistant software)对光响应曲线进行非直角双曲线[16,17]拟合(式1), 求算出饱和净光合速率P, 表观量子效率()和暗呼吸效率()等光合生理参数,WUE = P/T, WUE为水分利用效率,为净光合速率,为蒸腾速率。
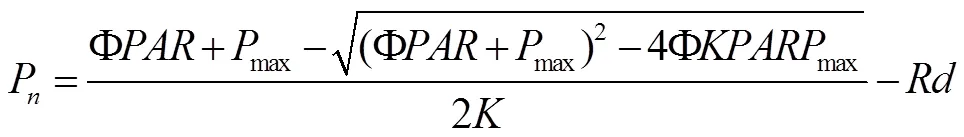
式中,P为净光合速率(μmol·m–2·s–1);为光合有效辐射(μmol·m–·s–1);P为一定CO2浓度下最大净光合速率(μmol·m–2·s–1);为光响应曲线的初始斜率, 即表观量子效率(, μmol·μmol–1);为光响应曲线的曲角;为暗呼吸速率(μmol·m–2·s–1)。
原始实验数据的处理采用Microsoft Excel 2013完成, SPSS17.0进行单因素法方差分析(One-way ANOVA), Origin8.0制图。
2 结果与分析
2.1 环境因子日变化
测量期间各环境因子如图1所示, 光合有效辐射日变化呈单峰曲线, 在14:00达到最大值(1869μmol·m–2·s–1左右), 气温(T)日变化也呈单峰曲线, 在16:00时刻达到最大值, 空气相对湿度在8:00—12:00急剧下降, 可能是因为随着温度升高空气中水蒸气蒸发加剧, 空气中的水汽含量降低, 湿度也随着降低。在12:00—14:00由于地表蒸散作用空气相对湿度又快速上升, 16:00—20:00, 空气相对湿度()表现平稳, 呈现缓慢增加的趋势。大气CO2浓度()波动范围大致在380—390 μmol·mol–1, 8:00—12:00, 大气CO2浓度()逐渐降低, 12:00—18:00, 曲线变化趋势平缓, 傍晚时浓度略有增加。
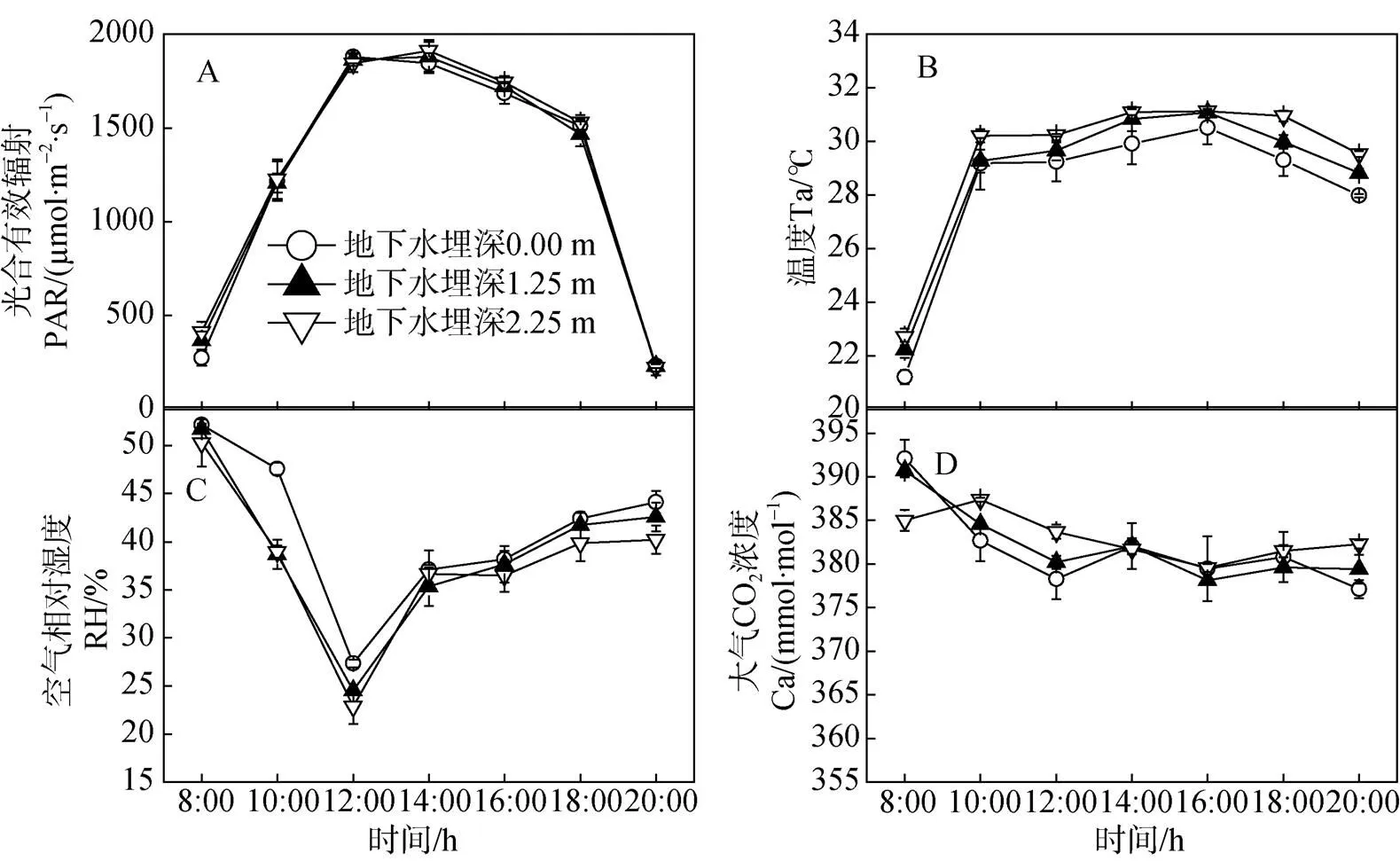
图1 环境因子日变化
2.2 塔干柽柳的净光合速率动态变化
如图2所示, 塔干柽柳光合-光响应曲线拟合的决定系数均在0.96以上, 拟合结果可以较好的反映塔干柽柳叶片对光的响应过程。低光强环境下, 不同地下水埋深处柽柳净光合速率P随光合有效辐射的增大均迅速增加, 达到一定的后,P增幅渐趋平缓呈逐渐下降趋势。方差分析结果显示, 不同地下水埋深处塔干柽柳的光合-光响应(P)有显著差异(<0.05)。当>800 μmol·m–2·s–1时, 不同光照条件下, Pn值均表现为:P地下水埋深2.25m>P地下水埋深1.25m>P地下水埋深0m。
净光合速率在一定的环境条件下反映了植物叶片的最大光合能力。对不同地下水埋深处塔干柽柳的P值进行线性回归(表1), 并对P值在从1至16范围内进行积分运算, 结果表明: IV地下水埋深2.25m>IV地下水埋深1.25m>IV地下水埋深0m(IV为integrated value的缩写)。这说明地下水埋深2.25 m处柽柳的光合作用能力最强。
2.3 柽柳光合-光响应特征参数
如表2所示, 在一定环境条件下, 植物叶片的饱和净光合速率(P)反映了植物利用强光进行光合作用的能力。单因素方差分析结果表明, 不同地下水埋深处塔干柽柳的最大净光合速率组间差异显著(<0.05), 饱和净光合速率的值表现为:P地下水埋深2.25m>P地下水埋深1.25m>P地下水埋深0m。表观量子效率()是反映植物在弱光下吸收、转换和利用光能的指标[18]。由表2可知, 3种地下水埋深处塔干柽柳的值在0.027—0.043之间,地下水埋深2.25 m处其值最大, 且差异达到显著(<0.05)。光补偿点()和光饱和点()是反映植物对光照需求大小的指标[18]。由表2可知, 塔干柽柳具有高光饱和点(963.6—2107.6 μmol·m–2·s–1)和高光补偿点(29.7—74.8 μmol·m–2·s–1), 均为耐阴性很弱的喜光植物。这可能与博斯腾湖流域光源充足的自然条件具有一定的关系。
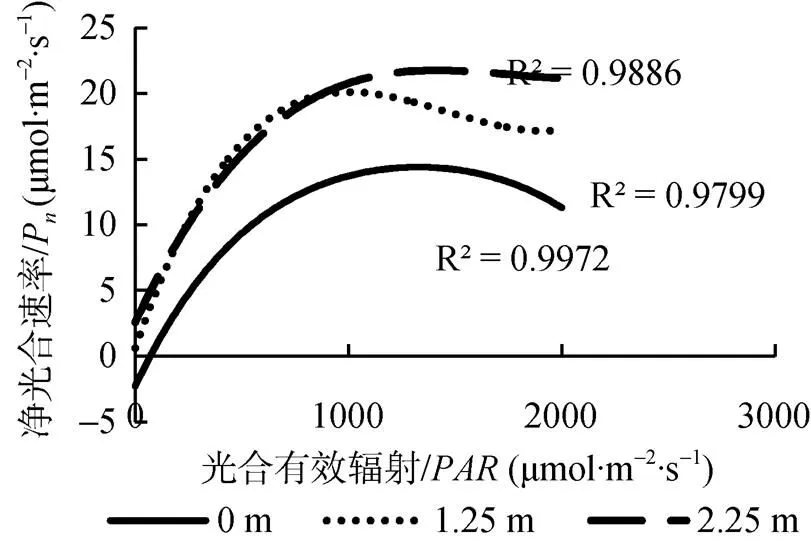
图2 不同地下水埋深处塔干柽柳的光合-光响应曲线
2.4 塔干柽柳蒸腾速率的光响应特征
由图3可知, 在0—800 μmol·m–2·s–1范围内, 塔干柽柳的气孔导度()都随着光合有效辐射的增加而增加, 当>800 μmol·m–2·s–1以后, 随着光强的进一步增加, 地下水埋深0 m和1.25 m处的塔干柽柳气孔导度而呈平稳的变化趋势, 地下水埋深2.25 m处塔干柽柳的气孔则进一步张开, 气孔阻力减小,随着的增强而继续增大。

表1 净光合速率(Pn)的定积分值
注: 变量y代表P, x代表。
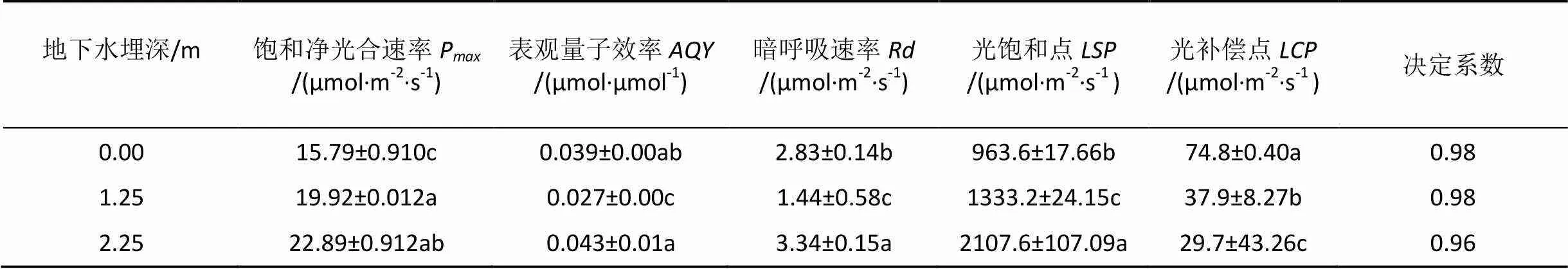
表2 不同地下水埋深处塔干柽柳光响应曲线特征参数
注: 采用Duncan 法(<0.05)检验, 数据中标不同字母表示差异显著不同。
胞间CO2浓度()是外界CO2气体进入叶肉细胞过程所受各种驱动力和阻力以及叶片内部光合作用和呼吸作用的平衡结果。图3反映出在0—1000 μmol·m–2·s–1范围内, 随着光强的增加不同地下水埋深处塔干柽柳的胞间CO2浓度()均呈下降的趋势, 地下水埋深0 m处塔干柽柳的值整体大于地下水埋深1.25 m和2.25 m。当800 μmol·m–2·s–1, 地下水埋深1.25 m和2.25 m处塔干柽柳的值随光强的增加缓慢增加, 地下水埋深0 m处则保持平稳的变化趋势。值由高到低依次为地下水埋深0 m>地下水埋深2.25 m>地下水埋深1.25 m。
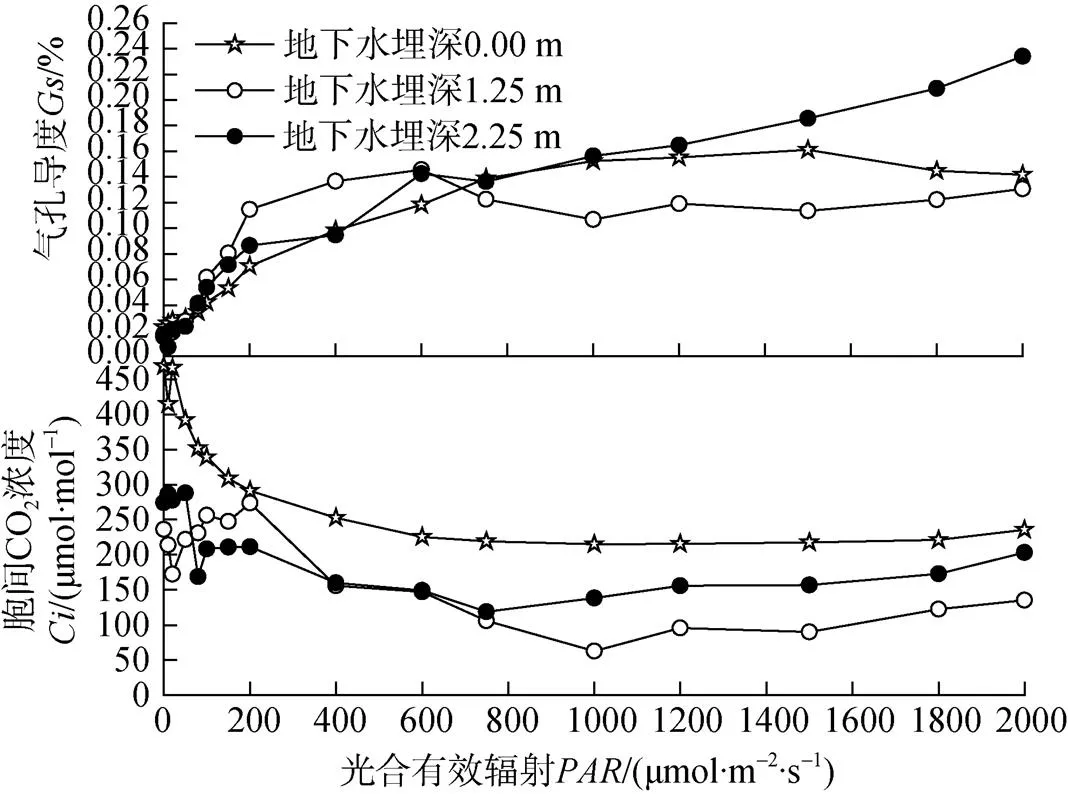
图3 不同地下水埋深处塔干柽柳的气孔导度Gs和胞间CO2浓度(Ci)的光响应
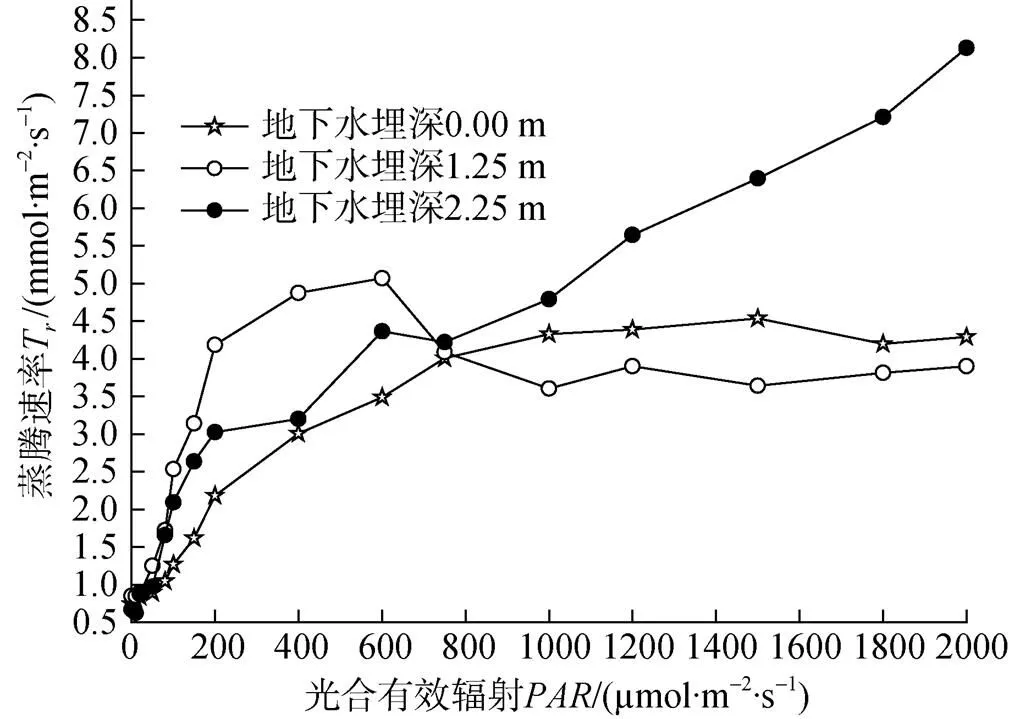
图4 蒸腾速率响应曲线
由图4可知,在0—600 μmol·m–2·s–1范围时, 地下水埋深1.25m处塔干柽柳的蒸腾作用随光合有效辐射的增加而增强, 当在600—1000 μmol·m–2·s–1时, 蒸腾速率随光合有效辐射的增强急剧减弱。当在0—1000 μmol·m–2·s–1范围时, 地下水埋深0 m处和2.25 m处塔干柽柳的T随光强的增加呈增加趋势, 当>1000 μmol·m–2·s–1, 地下水埋深0 m和1.25 m处塔干柽柳的T变化趋势较缓慢平稳, 而地下水埋深2.25 m处随光合有效辐射的增加蒸腾速率迅速增加。
2.5 塔干柽柳水分利用效率的光响应特征
水分利用效率()是植物光合和蒸腾特性的综合反映, 可以反映在干旱缺水的环境条件下植物耐旱生产力的高低[19], 由图5可知, 不同地下水埋深处塔干柽柳水分利用效率的光响应基本都呈单峰曲线, 当>400 μmol·m–2·s–1不同地下水埋深处塔干柽柳均维持较高的水分利用效率。当=1000 μmol·m–2·s–1时, 地下水埋深0 m和1.25 m处塔干柽柳的水分利用效率最大, 当接近800 μmol·m–2·s–1时, 地下水埋深2.25 m处塔干柽柳的水分利用效率最大, 当>800 μmol·m–2·s–1, 水分利用效率大小依次表现为: 地下水埋深1.25 m>地下水埋深2.25 m>地下水埋深0 m。
3 结论与讨论
不同植物的生长对地下水位要求的深度不一样, 随地下水位变化会表现出不同的响应[20]。通过植物光合-光响应曲线(–)得到的光合生理参数可以判断不同尺度植物对环境的适应状况, 并为植物的适应性评价提供理论依据[21–23]。本研究对不同地下水埋深处塔干柽柳的光合-光响应特征进行了研究。由表2可知, 地下水埋深2.25 m处塔干柽柳的饱和净光合速率(P)最大(22.89 μmol·m–2·s–1), 比地下水埋深0m(15.79 μmol·m–2·s–1)高31%, 这说明不同地下水埋深环境下塔干柽柳利用强光进行光合作用的能力有显著差异。不同地下水埋深处柽柳的光饱和点(963.6—2107.6 μmol·m–2·s–1)的变化范围明显大于光补偿点(29.7—74.8 μmol·m–2·s–1)的变化幅度, 这进一步说明不同地下水埋深环境下塔干柽柳利用强光进行光合作用的能力有明显差异, 地下水埋深2.25 m处塔干柽柳的光饱和点最高, 光补偿点最低, 说明此地下水埋深条件对强光环境的适应性最强。有研究表明植物的表观量子效率理论值为0.08—0.125, 但在自然条件下其值为0.04—0.07[16], 本研究柽柳值在0.027—0.043之间, 表现为较低水平但其值在一般植物的范围内, 这说明其喜光但不是典型的阳生植物。暗呼吸速率()可以反应植物叶片的生理活性, 在本研究中, 地下水埋深2.25 m处柽柳的>3 mol·m–2·s–1有较强的呼吸作用, 生理活性最强, 能够较地下水埋深0 m和1.25 m更好的生长。这与周晓冰[24]等人的研究结果相一致。
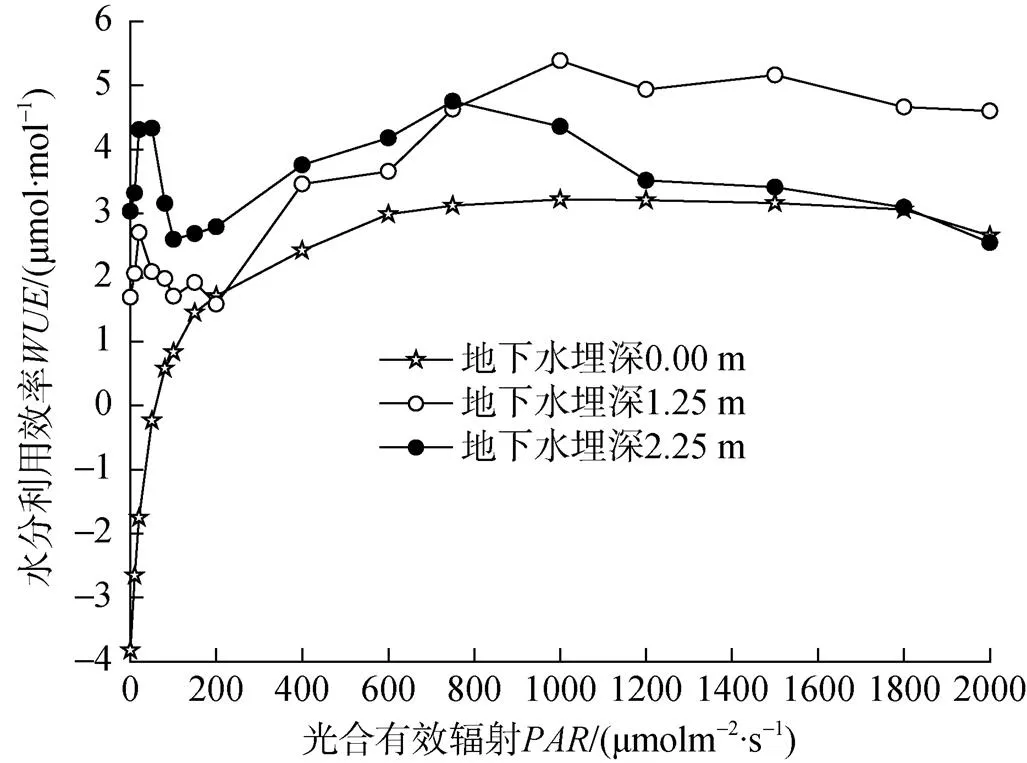
图5 塔干柽柳水分利用效率的光响应
当在0—1000 μmol·m–2·s–1范围内, 胞间CO2浓度随着的增强而减小, 叶室内的CO2浓度由外源钢瓶维持在350 μmol·mol–1的恒定水平,值越低, 就越有利于CO2进入叶片, 光合潜力就越大。当>1500 μmol·m–2·s–1, 地下水埋深1.25 m和2.25m处塔干柽柳的胞间CO2浓度呈现出随光强增加而增加的趋势, 可能是这段光强范围内, 这两处塔干柽柳气孔导度有一个迅速增加, 气孔的开合削弱了CO2进入叶片的阻力, 叶室内的CO2顺着CO2梯度进入叶室, 造成的增加。这与王会提和伍维模等人的研究结果一致[13,26]。
蒸腾作用是植物水分关系中起主导作用最重要的过程之一[13]。不同地下水埋深处塔干柽柳蒸腾速率和气孔导度值的变化基本相似。在一定的光强范围内, 随着气孔导度的降低蒸腾速率随之降低, 随着气孔导度的增加蒸腾速率进一步增大, 这说明气孔导度和蒸腾速率之间呈显著的正相关关系。如图4所示, 当>800 μmol·m–2·–1时, 地下水埋深2.25 m处塔干柽柳蒸腾速率迅速增加, 地下水埋深0 m和1.25 m则变化趋势较为缓慢平稳, 胞间CO2浓度和气孔导度也在同期具有相同的变化趋势, 可能是由于受强光的诱导, 气孔导度骤然增大, 水分通过气孔大量散失, 使得蒸腾速率迅速增大。
水分利用效率在一定程度上能够衡量植物的耗水抗旱性。吴桂林等人[25]发现柽柳在水分利用方面倾向于灵活的适应水分生境波动。本研究中, 当>400 μmol·m–2·s–1, 3处地下水埋深处的塔干柽柳维持较高的水分利用效率, 净光合速率也随着光强的增加逐步增大, 当>800 μmol·m–2·s–1时, 地下水埋深2.25 m处塔干柽柳水分利用效率有明显的下降, 这与同期气孔导度骤增引起蒸腾作用增大有很大关系。
综上所述, 在地下水埋深2.25 m处塔干柽柳的冠幅最大长势最佳, 光合作用能力最强, 具有较高水分利用效率和较强的蒸腾作用, 生理特性表现出最好的适应性, 这种响应特征暗示了在博斯腾湖滨, 塔干柽柳适宜生长地下水位在2.25 m左右。
本文只是在特定时段内对博斯腾湖北岸不同地下水埋深处塔干柽柳光合-光响应特征的研究, 为了更加深入地了解湿地植物对湿地生态系统地下水埋深生境波动的响应规律, 今后还需要做大量细致的研究工作, 以期为该区域博斯腾湖湿地自然植被的恢复和保护提供依据。
[1] 伊丽努尔·阿力甫江, 海米提·依米提, 麦麦提吐尔逊·艾则孜, 等. 1958—2012年博斯腾湖水位变化驱动力[J]. 中国沙漠, 2015, 35(1): 240–247.
[2] 芦晓峰, 苏芳莉, 周林飞, 等. 芦苇湿地生态功能及恢复研究[J]. 西北林学院学报, 2011, 26(4): 53–58.
[3] 尹林克. 中亚荒漠生态系统中的关键种──柽柳(spp)[J]. 干旱区研究, 1995, 12(3): 43–47.
[4] XU H, LI Y, XU G Q, ZOU T. Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation[J]. Plant, Cell and Environment, 2007, 30(4) : 399–409.
[5] HORTON J L, KOLB T E, HART S C. Responses of riparian trees to interannual variation in ground water depth in a semi-arid river basin[J]. Plant, Cell and Environment, 2001, 24(3): 293–304.
[6] 许皓, 李彦, 谢静霞, 等. 光合有效辐射与地下水位变化对柽柳属荒漠灌木群落碳平衡的影响[J]. 植物生态学报, 2010, 34(4): 376–386.
[7] 陈敏, 陈亚宁, 李卫红, 等. 不同地下水埋深柽柳、芦苇的生理响应[J]. 干旱区地理, 2009, 32(01): 72–80.
[8] 肖生春, 肖洪浪. 极端干旱区湖岸柽柳径向生长对水环境演变的响应[J]. 北京林业大学学报, 2006, 28(2): 39–45.
[9] 张佩, 袁国富, 庄伟, 等. 黑河中游荒漠绿洲过渡带多枝柽柳对地下水位变化的生理生态响应与适应[J]. 生态学报, 2011, 31(22): 6677–6687.
[10] HORTON J L, KOLB T E, HART S C. Responses of riparian trees to interannual variation in ground water depth in a semi-arid river basin [J]. Plant, Cell and Environment, 2001, 24(3): 293–304.
[11] 柴宝峰, 李毳. 甘蒙柽柳与沙棘抗旱性研究[J]. 应用与环境生物学报, 1998, 4(1): 24–27.
[12] 邓雄, 李小明, 张希明, 等. 多枝柽柳气体交换特性研究[J]. 生态学报, 2003, 23(1): 180–187.
[13] DENG X, LI X M, ZHANG X M, et al. Studies on gas exchange ofLbd [J]. Acta Ecologica Sinica, 2003, 23(1), 180–187.
[14] 王会提, 曾凡江, 张波, 等. 不同种植方式下柽柳光合生理参数光响应特性研究[J]. 干旱区地理, 2015, 38(4): 753–762.
[15] 刘彬, 王琴, 张海燕. 等. 博斯腾湖芦苇湿地生态环境现状及芦苇生物量影响因素分析[J]. 农业现代化研究, 2014, 35(3): 335–339.
[16] 马蓉, 麦麦提吐尔逊·艾则孜, 海米提·依米提, 等. 新疆博斯腾湖北岸芦苇叶片气孔导度特征及数值模拟[J]. 西北农业学报, 2016, 25(1): 123–128.
[17] FARQUHAR G D. Models of photosynthesis [J]. Plant Physiology, 2001, 125(1): 42–45.
[18] 刘宇锋, 萧浪涛, 童建华, 等. 非直线双曲线模型在光合光响应曲线数据分析中的应用[J]. 中国农学通报, 2005, 21(8): 76–79.
[19] 王艳杰, 薛达元, 彭羽. 盐碱胁迫对两个葡萄品种光合作用-光响应特性的影响[J]. 北方园艺, 2011(16): 30–36.
[20] 赵平, 曾小平, 彭少麟, 等. 海南红豆()夏季叶片气体交换、气孔导度和水分利用效率的日变化[J]. 热带亚热带植物学报, 2000, 8(01): 35–42.
[21] 陈亚宁, 李卫红, 陈亚鹏, 等. 新疆塔里木河下游柽柳、芦苇对生态输水的响应[J]. 冰川冻土, 2004, 26(5): 595–601.
[22] 靳甜甜, 刘国华, 胡婵娟, 等. 黄土高原常见造林树种光合蒸腾特征[J]. 生态学报, 2008, 28(11): 5758–5765.
[23] CLENDENNEN S K, ZIMMENRMAN R C, POWER D A, et al. Photosynthetic response of the giant kelp(phaeophyceae) to ultraviolet Radiation [J]. Journal of Phycology, 1996, 32(4): 614–620.
[24] 张卫强, 曾令海, 王明怀, 等. 东江中上游主要造林树种光合生理特性[J]. 生态环境学报, 2011, 20(1): 51–57.
[25] 周晓兵, 张元明, 王莎莎, 等. 3 种荒漠植物幼苗生长和光合生理对氮增加的响应[J]. 中国沙漠, 2011, 31(1): 82–89.
[26] 吴桂林, 蒋少伟, 周天河, 等. 不同地下水埋深胡杨与柽柳幼苗的水分利用策略比较[J]. 干旱区研究, 2016, 33(6): 1209–1216.
[27] 伍维模, 李志军, 罗青红, 等. 土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特性的影响[J]. 林业科学, 2007, 43(5): 30–35.
党亚玲, 韩炜, 马霄华, 等. 博斯腾湖北岸不同地下水埋深对塔干柽柳光合特性的影响[J]. 生态科学, 2017, 36(6): 188-194.
DANG Yaling, HAN Wei, MA Xiaohua, et al. Effects on photosynthetic characteristics ofto different groundwater depth on the north shore of Boston Lake[J]. Ecological Science, 2017, 36(6): 188-194.
Effects on photosynthetic characteristics ofto different groundwater depth on the north shore of Boston Lake
DANG Yaling1,2,3, HAN Wei1,2,*, MA Xiaohua1,2, GUAN Wenke3, FEI Bingqiang1,2, CUI Kaiqiang1,2, ZHANG Ya1,2
1. Key Laboratory of Arid Area of Lake Environment and Natural Resources of Xinjiang, Urumqi 830054, China 2. The School of Geographic Sciences and Tourism, Xinjiang Normal University, Urumqi 830054, China 3. Xinjiang Academy of Forestry Science, Urumqi 830054, China
On the north shore of Bosten Lake, we took, the dominant species, as research object to analyze the response and adaptation mechanism to different groundwater depth. Along the vertical lake shoreline on the north shore, we chose some same ageat the groundwater depth 0 meter, 1.25 meter and 2.25 meter, and used Li-6400XT, a portable photosynthesis system, to measure the photosynthetic light response.Results were as follows.(1) When the photosynthetic active radiation () was higher than 800 μmol·m–2·s–1, the net photosynthetic rate () values of three plants were ranked as groundwater depth 2.25 meter > groundwater depth 1.25 meter > groundwater depth 0 meter at the same light intensity. (2) When the groundwater depth was 2.25 m, thevalue maintained the highest, so did the dark respiration rate, and the light saturation point, but its light compensation point was the lowest, which indicated that theat this groundwater depth had the best ability to environment. (3) When thewas higher than 1000 μmol·m–2·s–1, the intercellular CO2concentration() and transpiration rate () increased with the enhance of light intensity at the groundwater depth 2.25 m. While the trend was relatively stable at the groundwater depth 0 m and 1.25 m. (4) When thewas less than 400 μmol·m–2·s–1, the overall values of three water use efficiency () response curves were roughly as follows:of groundwater depth 2.25 m > the one of groundwater depth 1.25m > the one of groundwater depth 0 m. The results indicated that various physiological indexes ofshowed the best, such as photosynthetic parameters and water use efficiency, which told that thoseat the groundwater depth 2.25 meter showed the best adaptability to wetland environment on the north shore of Bosten Lake, and 2.25 meter might be the optimal groundwater depth for growth ofin Bosten Lake.
Bosten Lake; groundwater depth;; Photosynthetic characteristics
10.14108/j.cnki.1008-8873.2017.06.026
Q494
A
1008-8873(2017)06-188-07
2017-08-06;
2017-10-9
塔里木河中下游胡杨生态修复研究与示范(HY-2.3); 教育部人文社会科学研究项目青年基金-新疆焉耆盆地生态安全研究(14XJJCZH003); 新疆师范大学博士科研启动基金-克里雅河流域芦苇对水盐运移的生理生态响应(XJNUBS1214); 新疆干旱区湖泊环境与资源重点实验室开放基金-博斯腾湖生态环境特征对区域气候变化的响应机制研究(XJDX0909-2012-07)
党亚玲(1990—), 女, 甘肃定西人, 研究生, 在读硕士, 主要从事绿洲生态建设与规划, E-mail: daqngyaling@yeah.net
韩炜, 副教授, 硕士生导师, 研究方向: 绿洲生态建设与规划, E-mail: hanweiaaa@163.com
