3个南亚热带季风常绿阔叶林群落主要性状特征比较
2017-12-29易慧琳郭颖涛刘玲刘晓丹黄柳菁刘世忠张倩媚刘楠韩瑞宏任海
易慧琳, 郭颖涛, 刘玲, 刘晓丹, 黄柳菁, 刘世忠, 张倩媚, 刘楠, 韩瑞宏, 任海,*
3个南亚热带季风常绿阔叶林群落主要性状特征比较
易慧琳1,2, 郭颖涛2, 刘玲2, 刘晓丹2, 黄柳菁1, 刘世忠1, 张倩媚1, 刘楠1, 韩瑞宏3, 任海1,*
1. 中国科学院华南植物园, 广州 510650 2. 广州普邦园林股份有限公司,广州 510600 3. 仲恺农业工程学院, 广州 510225
以南亚热带地区保存相对完好的鼎湖山、大岭山、白云山的季风常绿阔叶林为研究对象, 分析比较了各群落的生活型谱、Raukiaer 叶特征、群落果实类型和植物观赏特性等主要性状特征。结果表明, 这3个群落以高位芽植物占绝对优势, 单叶、革质、全缘、中叶比例较高; 3个群落由于演替时间的不同, 生活型谱有所差异, 鼎湖山云南银柴—香楠群落内中高位芽植物数量最多, 大岭山华润楠—慧竹群落以中高位芽和小高位芽植物为优势, 而白云山荷木—九节群落中小高位芽植物为优势; 不同叶特征植物比例也有所不同, 鼎湖山云南银柴—香楠群落中具中型叶、纸质叶等中生植物特征比例高于其他两个群落; 3个群落中肉质果植物占优势, 果实以被动扩散为主; 3个群落中在花、果、叶和其它部位(根、茎及树型姿态)具有一定的观赏价值植物分别有69、56、41和34种。在这些结果的基础上可以对南亚热带地带性城市森林群落建设提出适当建议。
南亚热带; 季风常绿阔叶林; 生活型; Raukiaer叶特征; 果实类型; 观赏特性
1 前言
季风常绿阔叶林是中国南亚热带季风气候区的地带性植被, 其群落结构复杂、生物多样性丰富, 是亚热带陆地生态系统的重要组成部分[1–2]。它是区域内人类生产活动所必需的多种药材、木材、食物及工业原料等的主要来源, 同时还具有涵养水源、保持水土、调节气候等生态系统服务功能, 对保持区域生态系统的平衡和稳定有着不可替代的作用[3]。对南亚热带季风常绿阔叶林的研究经历了从面上调查到长期定位研究, 目前已在广东省鼎湖山和黑石顶、云南哀牢山、福建武夷山建设了长期定位研究站[3–7]。这些研究也经历了从定性到定量的过程, 研究内容也从本底调查向群落结构、动态、生物量和生产力、生态系统结构与功能关系, 生态系统关键过程及其耦合对全球变化的响应与适应性等深化[3,8–9]。目前对南亚热带季风常绿阔叶林的自然植被分布、物种组成、生物量与生产力、林冠层CO2通量、群落动态、物种多样性和群落结构等的研究较为深入[3,10–11]。未见有从城市森林建设角度对南亚热带季风常绿阔叶林群落特征进行多样地的综合比较研究。
随着城市化进程的加快, 中国城市生态环境的建设明显滞后于社会经济的发展, 严重的生态环境问题给城市的可持续发展带来了不利的影响, 威胁到城市居民的生活及生存[12–13]。在生态、经济、社会和文化驱动下, 城市森林成为解决城市生态环境问题的有效途径, 森林城市成为城市生态建设的目标[14]。城市森林具有调节碳氧平衡[15–16]、改善城市小气候[17–18]、涵养水源、保持水土、防风固沙、净化空气[19]及降低噪音等生态功能, 还有游憩[20]、保健[21]、文化教育、拉动产业发展、促进科学技术进步等社会服务功能[22], 是城市文化、社会、自然的有机交会。城市森林建设应强调区域地带性, 模拟地带性植物群落、以近自然方法建设城市森林可最大限度提高群落生态效益[3]。对地带性植物群落进行研究, 有助于把握地带性植物群落的主要特征, 在园林景观设计的原则、方法的基础上, 更好地营造地带性城市森林景观。本研究以南亚热带地区保存相对完好的鼎湖山、大岭山、白云山的季风常绿阔叶林为研究对象, 围绕“南亚热带地带性群落有何主要性状特征”这一科学问题, 对其生活型谱、Raukiaer 叶特征、群落果实类型、群落植物观赏特性等群落特征进行比较分析, 为南亚热带群落研究积累研究数据的同时, 为南亚热带地带性城市森林群落建设提供参考。
2 自然概况
本研究所选择的三个研究地点均位于我国南亚热带(图1), 鼎湖山自然保护区(DHS)(112°30′—112° 33′E, 23°09′—23°11′N), 白云山风景区(BYS) (113° 16′—113°19'E, 23°09′—23°13′N) , 大岭山(DLS) (113°46′—113°49′E, 23°37′—23°39′N)分别位于广东省肇庆市, 广州市中心以及广东省广州市东北部。
三个样地均属于南亚热带季风气候, 其水热条件及土壤条件相似(表1), 年平均气温在20.9—21.7 ℃之间, 有干湿季之分, 年平均降水量1727—1927 mm, 鼎湖山和白云山位于南亚热带中部, 降水集中在4—9月, 年平均湿度分别为81.5%和79.1%[23], 大岭山位于南亚热带偏北部, 降水主要集中在5—8月份, 相对湿度多在85%以上[24]。三个样地地带性土壤均以赤红壤为主[23,25–26]。鼎湖山森林群落复杂多样, 地带性植被为位于保护区中心区域的南亚热带季风常绿阔叶林。另外还存在大部分针阔混交林、针叶林以及少量的南亚热带沟谷雨林和山地常绿灌草丛。而白云山的现状植被是人工植被和天然植被的混合体[27], 白云山的地带性植被是次生常绿阔叶林、针阔混交林、马尾松针叶林。大岭山到了冬季出现低温现象, 温度较周边偏低, 所以除了常绿阔叶林、山顶矮林及亚热带针叶林、针阔混交林等[28], 还出现了许多落叶阔叶林或常绿与落叶阔叶混交林。

注: DHS:鼎湖山自然保护区; BYS: 白云山风景区; DLS: 大岭山; GZ: 广州

表1 3个研究样地基本信息
本研究分别选取了3地保存相对完整的鼎湖山云南银柴—香楠群落(community)、大岭山华润楠—慧竹群落(community)、白云山荷木—九节群落(community)为研究对象。鼎湖山云南银柴—香楠群落位于鼎湖山庆云寺后山, 群落郁闭度高, 以中生植物云南银柴为优势乔木种, 群落有较明显的亚层分化, 第一亚层为高大阳生植物, 以第三亚层为优势乔木亚层。灌木层高度多集中在0.8—1.5m之间, 以香楠()、柏拉木()、罗伞树()、黄果厚壳桂()、褐叶柄果木()等为优势灌木(含乔木层苗木); 受群落郁闭度影响, 林下草本层的种类不多, 以乔灌木幼苗及蕨类植物、禾本科(Gramineae)、姜科(Zingiberaceae)植物为主; 层间植物种类不多, 但多为大型木质藤本, 如杖藤()、宽药青藤()、丁公藤()、扁担藤()等。大岭山华润楠—慧竹群落位于大岭山石灶工区, 除明显的乔灌草三个基本层次外, 群落乔木层分为明显的三个亚层, 且高大乔木丰富, 以华润楠()为优势种; 灌木层和草本层物种组成丰富, 以耐阴植物及乔木幼苗为主; 群落中常见藤本植物有肖菝葜()、甜果藤()、络石()、菝葜()、白花油麻藤()等南亚热带地带性藤本植物。白云山荷木—九节群落位于白云山山庄旅社附件, 分为乔灌草三个基本层次, 乔木层以荷木()、降真香()为优势种, 并未见明显的亚层分化; 灌木植物以耐阴植物为主, 优势种为九节()、变叶榕(varolosa)、狗骨柴()等; 草本层以乔木、灌木的幼苗为主, 草本植物较少, 主要为蕨类植物、姜科、禾本科等为主; 群落层间有藤本植物分布。
3 研究方法
白云山、大岭山群落调查于2010年3月至10月间完成, 利用典型群落样方法对两个样地的季风常绿阔叶林进行植被调查的数据。在大岭山石灶工区设置28个100 m2(10×10 m2)样方, 样地面积共2800 m2; 在白云山山庄旅社设置12个100 m2(10× 10 m2)样方, 样地面积共1200 m2。调查乔木层(DBH≥2 cm)每木种类、高度、胸径、基面积和冠幅。再在每个10×10 m2样方中随机选取1个5 m×5 m的灌木样方, 在灌木层样方内随机套设1个1 m× 1 m的草本层调查样方,分别进行灌木层和草本层的调查。
鼎湖山数据采用鼎湖山森林生态系统定位研究站2010年调查的庆云寺后山南亚热带季风常绿阔叶林10000 m2(100×100 m2)大样方数据。2010年样地调查时记录了所有10×10 m2样方内DBH≥1cm的乔木层植物种名、胸径和坐标值, 每个10×10 m2样方分为4个5 m×5 m样方, 并从中随机选取1个5 m× 5 m作为灌木样方, 在灌木层样方内随机套设1个2 m×2 m的草本层调查样方, 分别进行灌木层和草本层的调查。为了与白云山、大岭山群落进行比较, 在大样方中随机抽取25个10×10 m2样方, 样地面积共2500 m2, 作为本次分析的数据, 并将DBH≥2 cm木本植物作为群落乔木层。
整理调查数据, 进行群落生活型谱、Raukiaer 叶特征、群落果实类型以及群落植物观赏特性分析。
4 结果与分析
4.1 南亚热带季风常绿阔叶林生活型特征
生活型是植物对外界环境适应的外部表现形式, 是植物对综合环境条件的长期适应, 在外貌上反映出来的植物类型。鼎湖山、白云山、大岭山三地的南亚热带季风常绿阔叶林群落中, 均是高位芽植物占绝对优势。由图2可知, 3个群落高位芽植物分布存在一定的差异。鼎湖山云南银柴—香楠群落内中高位芽植物最多, 有26种, 占23.21%, 其次为小高位芽和矮高位芽, 分别占18.75%和12.50%, 藤本高位芽有13种, 占11.61%。大岭山华润楠—慧竹群落中小高位芽和中高位芽植物比例相差不大分别为18.50%和18.00%, 为群落高位芽中最多的部分, 矮高位芽和藤本高位芽分别占8.00%和6.00%; 白云山荷木—九节群落中小高位芽最多, 有24种, 占22.02%, 其次为矮高位芽和中高位芽, 分别占14.68%和12.84, 藤本高位芽占12.84%。
鼎湖山、白云山、大岭山三个样地的季风常绿阔叶林群落中以高位芽植物占优势的生态型谱特征与南亚热带水热充沛的自然生态条件相适应。三个群落相比, 鼎湖山云南银柴—香楠群落内中高位芽植物数量最多, 大岭山华润楠—慧竹群落以中高位芽和小高位芽为优势, 而白云山荷木—九节群落中小高位芽数量最多。这种变化趋势很大程度上和3个群落分别所处的演替阶段有关。彭少麟将南亚热带森林群落演替的主要模式分别为针叶林—以针叶树种为主的针阔混交林—以阳生阔叶树种为主的针阔混交林—以阳生树种为主的常绿阔叶林—以中生植物为主的常绿阔叶林—中生群落[3]。鼎湖山云南银柴—香楠群落处于向中生群落发展的阶段[29-30], 群落中属于中高位芽植物的中生性植物类群发展壮大; 大岭山和白云山群落仍处于以阳生树种为主的常绿阔叶林阶段, 群落内中高位芽植物生长受到顶层高大阳生植物的影响较大; 且白云山荷木—九节群落为位于市中心的次生常绿阔叶林, 很大程度上受人为干扰, 在一定程度上影响了群落生活型谱分布。
在营造南亚热带地带性生态植物景观的过程中, 以中高位芽和小高位芽植物为主, 搭配部分高大大高位芽植物, 有助于快速建立相对稳定群落结构, 形成与地带性南亚热带季风常绿阔叶林外貌相似的生态群落景观。

图2 南亚热带季风常绿阔叶林生活型谱
4.2 南亚热带季风常绿阔叶林植物叶特征
植物群落叶型、叶质在一定程度上反映了组成群落植物的生态型和环境适应特点。表2从叶级、叶型、叶质、叶缘和叶尖等5个方面对南亚热带季风常绿阔叶林植物叶的特征进行分析。
3个不同群落叶级大小比例区别不大, 均以中型叶占优势, 其次为小型叶和大型叶, 小型叶数量均较大型叶多, 细型叶数量接近与0, 微型叶数量为0。3个群落中, 中型叶、小型叶、大型叶占总种数的比例之和, 均超过95%。
3个群落叶缘为全缘的植物占多数, 分别为83.93%、67.34%、71.30%。3个群落叶型均以单叶占绝对优势, 鼎湖山、大岭山、白云山三地群落中单叶植物比例分别为83.04%、81.41%和79.63%, 复叶植物以蕨类植物为主, 其中大岭山、白云山蕨类植物占复叶植物的一半左右。
3个群落叶质组成比例相似, 为革质叶>纸质叶>草质叶>膜质叶>肉质叶, 革质叶植物和纸质叶植物在南亚热带季风常绿阔叶林群落中占绝对优势, 革质叶比例分别为42.86%、52.26%和48.15%, 纸质叶分别为43.75%、29.63%和25.13%, 其中鼎湖山云南银柴—香楠群落纸质叶比例显著高于另外两个群落。
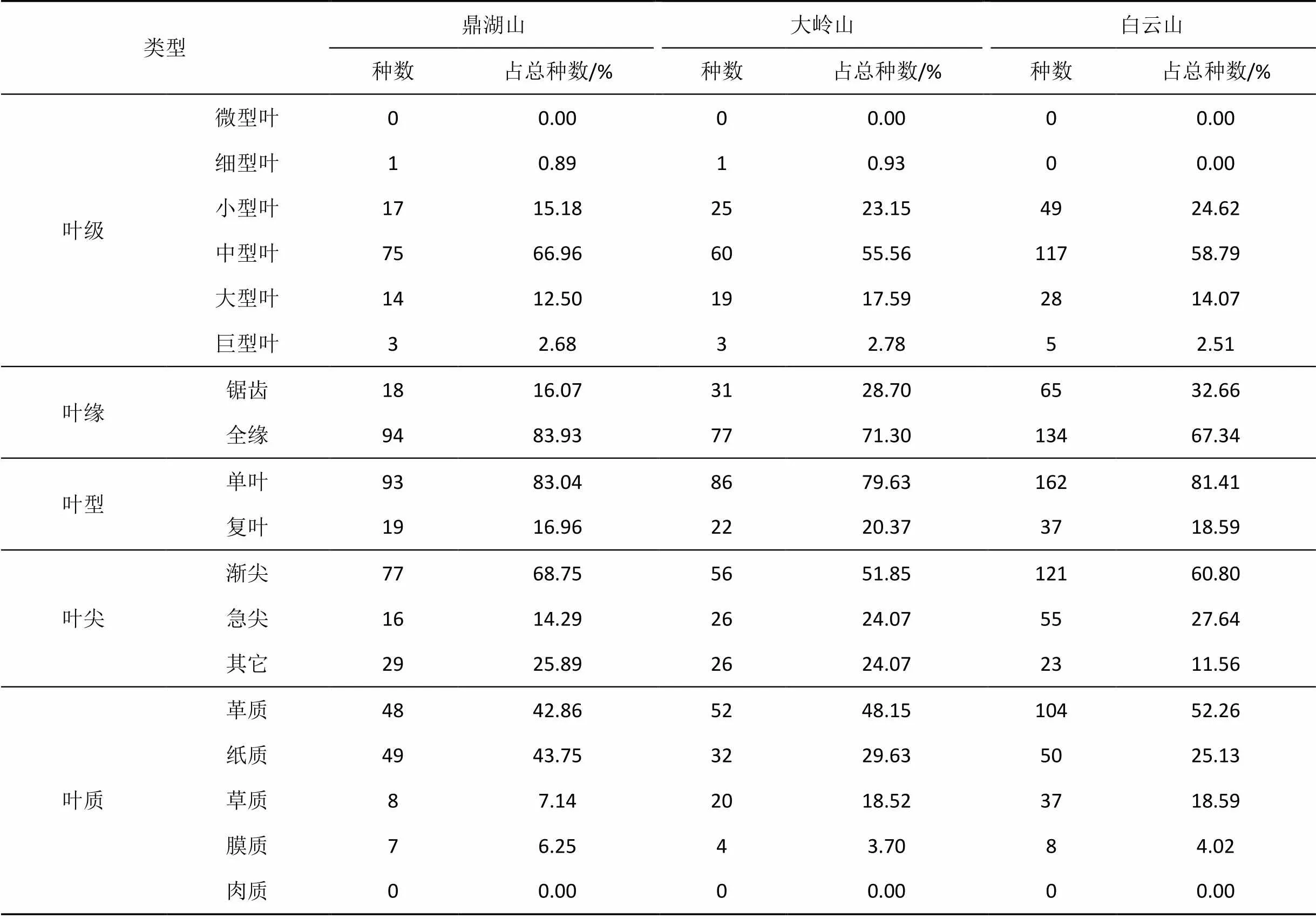
表2 南亚热带季风常绿阔叶林植物Raunkiaer叶特征
植物叶子的形状大小和颜色, 对构成群落外貌特征有重要意义, 而且与群落的生产率有关。大的叶片经常出现于热带温带而潮湿的气候中, 而小的叶片经常地出现于十分干燥和寒冷地区植物的特征[31], 3个群落叶级均以中型叶为主, 其中鼎湖山鼎湖山云南银柴—香楠群落的中型叶所占比例最大, 一定程度上说明以中型叶为主的叶级谱适应南亚热带温暖湿润的环境特征, 而鼎湖山鼎湖山云南银柴—香楠群落的适应性强于其他两个群落。挑选对区域气候环境适应性更强的中型叶植物, 有助于建设适宜南亚热带高温多湿的气候环境的人工地带性生态植物群落。
叶型以单叶为主, 所占比例达与常绿阔叶林群落单叶比率超过80%的特征基本一致[32]。群落中全缘叶所占比例高于非全缘叶, 且以革质叶所占比例最大, 符合亚热带常绿阔叶林的基本特征。一般来说革质叶植物适应强光照, 一般分布在群落上层, 而纸质叶和草质叶植物分布于群落中下层吸收弱光。各群落中的纸质叶也占很大比例, 分别为43.75%、29.63%和25.13%, 鼎湖山云南银柴—香楠群落纸质叶比例显著高于另外两个群落, 而大岭山、白云山群落中草质叶植物比例远大于鼎湖山群落。说明鼎湖山群落中吸收弱光的灌木小乔木数量远大于其他两个群落, 而大岭山、白云山群落中草质地被植物相对丰富。这与鼎湖山云南银柴—香楠群落处于中生顶级群落演替阶段, 群落以中生植物为主要优势种; 而大岭山、白云山群落尚处于以阳生树种为主的常绿阔叶林阶段, 群落以阳生大乔木为优势种, 地被植物相对丰富的群落特征相符合。
在建设南亚热带城市地带性园林植物景观时, 尽量模拟南亚热带季风常绿阔叶林群落植物叶特征, 选择全缘、革质、单叶植物为群落优势种类如华润楠、白颜树()、降真香等为优势乔木层物种, 群落下层搭配部分吸收弱光的纸质叶植物如香楠、罗伞、九节等, 以草质叶植物为主要地被植物如楼梯草()、三菅兰()、山姜()等, 形成科学合理的基本群落分层, 充分利用群落中的光照资源。
4.3 南亚热带季风常绿阔叶林植物果实类型
图3是3个南亚热带季风常绿阔叶林群落果实类型的分布比例分析, 群落中聚合果、聚花果比例非常低, 以单果为主。从图3中可以看出, 3个群落均以核果、浆果、蒴果占优势, 鼎湖山云南银柴—香楠群落核果、浆果、蒴果比例分别为30.77%、31.73%、和18.27%, 大岭山华润楠—慧竹群落核果、浆果、蒴果比例分别为18.54%、29.14%和19.21%, 白云山荷木—九节群落核果、浆果、蒴果比例分别为27.62%、21.90%和27.62%。鼎湖山、白云山和大岭山3个群落中肉质果(核果、浆果)占优势, 比例分别为62.50%、49.52%和47.68%。
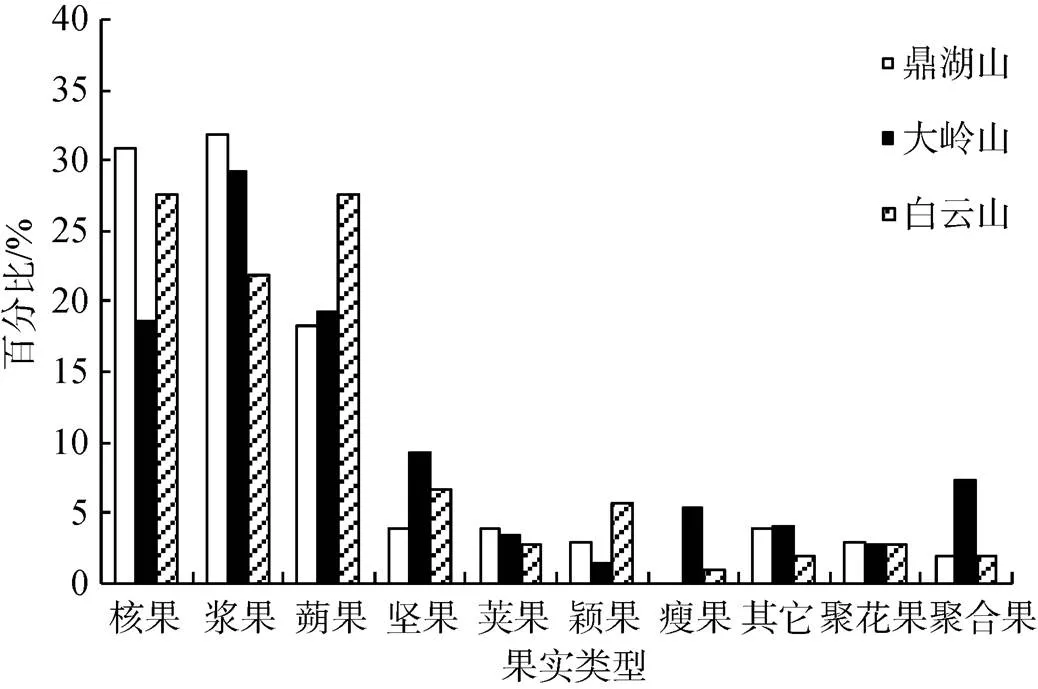
图3 南亚热带季风常绿阔叶林果实类型比例
果实类型的产生既是植物在地质历史过程中长期演化的产物, 也是植物群落果实类型的构成式样可能就是植物对生态环境的直接反映之一[33]。3个群落中以肉质果植物为主, 符合于顺利等随着纬度和海拔的升高, 肉果比例下降的研究结果[34]。在植物种子传播过程中动物、昆虫、鸟类等起到重要作用, 形成了群落以被动扩散为主的群落扩散机制, 促进植物群落的演变。在进行南亚热带地带性园林植物景观营造时, 选择肉质果植物作为群落优势种如杜英科(Elaeocarpaceae)、紫金牛科(Myrsinaceae)、樟科(Lauraceae)、桑科榕属(Ficus microcarpa)等植物, 有助于群落自我更新的同时还可以吸引鸟类、动物、昆虫等, 增加群落的活力, 实现人与自然的和谐共处。
4.4 南亚热带季风常绿阔叶林植物观赏特性
植物的形体和各个部位给人以美感是植物观赏特性的具体体现, 包括植物花、果、叶的形状、大小、颜色等和其它部位(根、茎及树型姿态)等带来的观赏效果。表3中列出了, 具有各观赏特性的植物种类。3个南亚热带季风常绿阔叶林群落中花的形状、色彩、大小、气味等具有明显特征, 优美可观、心旷神怡特点的植物种类共69种。在果实形状、大小、颜色等具有观赏性的植物共有56种。在叶颜色、大小、形状等具有观赏性的植物共有41种, 蕨类植物具有形状各异的叶型, 具有很高的观赏价值。其中共有45个物种同时具有两个或两个以上的观赏特性。

表3 南亚热带季风常绿阔叶林植物观赏特性
注: 上述植物拉丁文学名见附录。
为增加城市森林的居民参与性, 在在进行城市森林建设时不仅要考虑群落的生态性的同时还要关注群落的观赏性。表3中列出本文研究的3个南亚热带季风常绿阔叶林群落中具有良好观赏特性的植物, 可作为城市森林群落建设物种选择的参考资料。
5 结论
本研究中的3个南亚热带季风常绿阔叶林群落的生活型谱、Raukiaer 叶特征和群落果实类型特征进行比较, 并分析3个群落的植物观赏特性, 为南亚热带地带性城市森林景观营造, 生态园林建设提供参考。主要结论有:
5.1 3个群落以高位芽植物为主, 由于所处的演替阶段不同, 生态型谱有所差异, 鼎湖山云南银柴—香楠群落内中高位芽植物数量最多, 大岭山华润楠—慧竹群落以中高位芽和小高位芽植物为优势, 而白云山荷木—九节群落中小高位芽植物为优势; 建议城市地带性森林群落建设以中高位芽和小高位芽植物为主, 群落上层搭配高大大高位芽植物。
5.2 3个群落以单叶、革质、全缘、中叶植物为主 , 不同叶特征植物比例也有所不同, 鼎湖山云南银柴—香楠群落中具中型叶、纸质叶等中生特征的植物比例高于其他两个群落。建议南亚热带城市森林群落景观全缘、革质、单叶植物为群落优势乔木层, 中下层以纸质、草质叶植物为主。
5.3 3个群落中肉质果植物占优势, 群落种群扩散机制以被动扩散为主。为增加城市森林群落活力, 促进群落自主演替, 城市森林群落建设以肉质果植物为主。
5.4 统计了3个群落中在花、果、叶和其它部位(根、茎及树型姿态)具有一定的观赏价值植物分别有69、56、41和34种, 为建设具有强观赏性的城市森林群落, 提供了植物物种选择依据。
[1] 丁圣彦, 宋永昌. 常绿阔叶林植被动态研究进展[J]. 生态学报, 2004,24(8): 1769–1779.
[2] 宋永昌, 陈小勇, 王希华. 中国常绿阔叶林研究的回顾与展望[J]. 华东师范大学学报(自然科学版), 2005(1): 1–8.
[3] 彭少麟. 南亚热带森林群落动态学[M]. 北京: 科学出版社, 1996.8.
[4] 张宏达, 王伯荪, 张超常, 等. 广东高要鼎湖山植物群落之研究[J]. 中山大学学报, 1955, (3):161–229.
[5] 张祝平, 丁明懋. 鼎湖山亚热带季风常绿阔叶林的生物量和光能利用效率[J]. 生态学报, 1996, 10, 16(5): 525–534.
[6] 尹爱国, 苏志尧, 陈北光, 等. 广州白云山常绿阔叶林区系成分分析[J]. 华南农业大学学报(自然科学版), 2002, 4: 45–48.
[7] 史军辉, 黄忠良, 周小勇, 等. 鼎湖山森林群落多样性垂直分布格局的研究[J]. 生态学杂志, 2005, 24(10): 1143– 1146.
[8] 任海, 彭少麟. 鼎湖山森林生态系统演替过程中的能量生态特征[J]. 生态学报, 1999, 11(6): 817–822.
[9] ZHOU G Y, PENG C H, LI Y L, et al. A climate change– induced threat to the ecological resilience of a subtropical monsoon evergreen broadleaved forest in south China [J]. Global Change Biology, 2013, 4(19): 1197–210.
[10] 王伯荪, 彭少麟. 鼎湖山森林群落分析Ⅳ. 相似性与聚类分析[J]. 中山大学学报(自然科学版), 1985(1): 31–38.
[11] 王伯荪, 植物群落学[M]. 北京: 高等教育出版社, 1987.
[12] 章滨森. 城市森林建设的驱动模式与规划研究——以杭州半山国家森林公园为例[D]. 北京: 中国林业科学研究院, 2012.
[13] 朱俊. 中国城市森林学理论与实证研究[D]. 上海: 复旦大学, 2004.
[14] 聂法良. 我国城市森林多主体协同治理问题研究[D]. 哈尔滨: 东北林业大学, 2015.
[15] NOWAK D J, GREENFIELD E J, HOEHN R E, et al. Carbon storage and sequestration by trees in urban and community areas of the United States [J]. Environmental Pollution, 2013(178): 229–236.
[16] 吴耀星, 康文星, 郭清和, 等. 广州市城市森林对大气污染物吸收净化的功能价值[J]. 林业科学, 2009,3(5): 42–48.
[17] 黄少卫, 康文星, 吴耀星, 等. 城市森林对小气候的调节及其功能价值评估[J]. 中南林业科技大学学报, 2010,01 (1): 90–94.
[18] 李振军. 广州市萝岗区森林空间布局规划研究[D]. 华南理工大学, 2010.
[19] NOWAK D J, HIRABAYASHI S, BODINE A, et al. Tree and forest effects on air quality and human health in the United States [J]. Environmental Pollution, 2014(193): 119–129.
[20] 沈芝琴, 陈秋华, 程贵松, 等. 城市森林游憩功能评价研究——以福州国家森林公园和金牛山公园为例[J]. 林业经济问题, 2011,6(3): 228–233.
[21] 韩明臣. 城市森林保健功能指数评价研究——以北宫国家森林公园为例[D]. 北京: 中国林业科学研究院, 2011.
[22] 姚先铭, 康文星. 城市森林社会服务功能价值评价指标与方法探讨[J]. 世界林业研究, 2007, 8(4): 67–71.
[23] 张金泉. 广州白云山风景名胜区的植被特点及其林分改造[J]. 生态科学, 1995(1): 31–39.
[24] 李胜强, 崔杰. 广州石门国家森林公园维管植物资源调查[J]. 广东林业科技2013, 29(3): 37–42.
[25] 黄忠良, 孔国辉, 何道泉. 鼎湖山植物群落多样性的研究[J]. 生态学报, 2000, 3, 20(2): 193–198.
[26] 易绮斐, 成夏岚, 曾庆文, 等. 广州石门国家森林公园彩叶植物调查研究[J]. 福建林业科技, 2008, 35(1): 112– 116.
[27] 苏志尧, 古炎坤, 陈北光, 等. 广州白云山风景区的植被和主要植物群落类型[J]. 华南农业大学学报, 1999, 18(2): 23–29.
[28] 姜艳.石门国家森林公园植物群落特征及物种多样性研究[D].北京: 中国林业科学研究院, 2009.
[29] 彭少麟, 方炜. 鼎湖山植被演替过程优势种群动态研究Ⅲ. 黄果厚壳桂和厚壳桂种群[J]. 热带亚热带植物学报, 1994, 2(4): 79–87.
[30] 彭少麟, 方炜, 任海, 等. 鼎湖山厚壳桂群落演替过程的组成和结构动态[J]. 植物生态学, 1998, 22(3): 245–249.
[31] CAIN S A, DE OLIWEIRA CASTRO G M, PIRES J M, et al. Application of Some Phytosociological Techniques to Brazilian Rain Forest[J]. American Journal of Botany, 1956, 43: 911-941.
[32] GRUBB P J, LOYD J R, PENNINGTON T D, et al. A Comparison of Montane and Lowland Rain Forest in Ecuador I. The Forest Structure, Physiognomy, and Floristics[J]. Journal of Ecology, 1963, 51(3): 567–599.
[33] 陈学林, 田芳, 戚鹏程. 白水江自然保护区植物果实类型组成及垂直分异[J]. 林业科学, 2007, 43(60): 61–66.
[34] 于顺利, 王宗帅, 泽仁旺姆, 等. 种子地理学: 概念及其基本科学问题[J]. 应用生态学报, 2010, 21(1): 239–246.
易慧琳, 郭颖涛, 刘玲, 等. 3个南亚热带季风常绿阔叶林群落主要性状特征比较[J]. 生态科学, 2017, 36(6): 137-146.
YI Huilin, GUO Yingtao, LIU Ling, et al. A Comparative study on the community main morphological characteristics of three lower subtropical monsoon evergreen broad-leaved forests[J]. Ecological Science, 2017, 36(6): 137-146.
附录: 文中相关植物拉丁文学名
紫玉盘Roxb. var. microcarpa (Champ.) Finet et Gagnep.
棕叶芦
硃砂根Sims
栀子Ellis
越南紫金牛 AdisiaWall. var. cochinchinensis Pitard
玉叶金花Ait. f.
野牡丹D. Don
野含笑Dunn
艳山姜(Pers.) Burtt. et Smith
香花崖豆藤Harms
香花枇杷Champ.
腺叶桂樱
鼠刺Hook. et Arn.
十大功劳(Lindl.) Fedde
深山含笑Dunn
珊瑚树Ker-Gawl.
山蒲桃Merr. et Perry.
山姜(Thunb.) Miq.
山矾Buch.-Ham. ex D. Don
山杜英(Lour.) Poir
日本五月茶japoicum Sieb. et Zucc.
青皮木
青风藤Osbeck
楠藤Champ.
南五味子
南川茶H. T. Chang et Xiong
木莲
荷木Gardn. et Champ.
毛桃木莲
龙船花IxoraLam
阔叶十大功劳(Fort.) Carr.
九里香L.
金银花Thunb.
脚骨脆Gagn
尖叶杜英Masters
假鹰爪Lour.
假苹婆Cav.
鸡血藤
火棘(Maxim.) Li
黄杞Wall.
黄牛木(Lour.) Bl
黄花大苞姜(Hance) K. Larsen
华山姜(Retz.) Rosc.
中华杜英(Gardn)
花叶山姜Hook. f.
厚叶素馨Hand.-Mazz.
厚叶木莲
猴欢喜(Hance) Hemsl.
红枝蒲桃Merr. et Perry
红淡比Thunb
海芋
广东毛蕊茶Hand.-Mazz.
广东金叶子(Dop) Judd var. kwangtungense
观光木
瓜馥木(Hemsl.) Merr.
狗骨柴(Lindl.) Masam.
多花野牡丹D. Don
短尾叶柃Dunn
短柄紫珠(Benth.) Hance
杜英Hemsl.
大叶杜英A. DC
常绿荚蒾K. Koch
糙果茶(Merr.) Coh. St.
宝铎草D. Don
白花鬼灯笼
小叶买麻藤(Warb.) C. Y. Cheng ex Chun
野木瓜DC.
锈毛莓Ker
小叶红叶藤(Hook. et Arn.) Planch.
小盘木Planch.
肖蒲桃(Blume) Merr. et Perry
藤槐
算盘子L
树参(Harms) Merr
三花冬青Bl.
榕叶冬青Hemsl
毛菍Sims
毛果算盘子Champ. ex Benth
毛冬青Hook. et Arn.
蔓九节L.
岭南柿Dunn
岭南山竹子Champ. ex Benth
岭南槭Duthie
亮叶猴耳环Benth
虎舌红Hance
猴耳环(Jack) Benth.
广东蛇葡萄(Hook. et Arn.) Planch.
光叶红豆Y. C. Wu
光山香圆
橄榄(Lour.) Raeusch
倒卵叶野木瓜Hemsl.
赤楠Hook. et Arn.
草珊瑚(Thunb.) Nakai
滨海槭sino-oblongum Metc.
薜荔Linn.
白皮黄杞Merr.
白果香楠(Champ. ex Benth.) Tirveng.
白桂木Hance
窄叶半枫荷Roxb.
蜘蛛抱蛋(L.) DC.
鱼尾葵Hance
鹅掌柴(Lour.) Harms
肖菝葜
显脉杜英A. DC.
锡叶藤(Lour.) Hoogland
铁线蕨-L. f. capillus-veneris
山乌桕(Champ. ex Benth.) Muell. Arg.
山麦冬(Thunb.) Lour.
山菅兰(L.) DC
三叉蕨(Hook. et Arn.) Cop.
琴叶榕Hance
密脉鹅掌柴(Lour.) Harms
毛蕊茶Hand.-Mazz.
芒萁(Thunb.) Berhn
瘤足蕨(Bl.) Bedd.
楝叶吴茱萸(Champ. ex Benth.) Huang
厚壳桂(Hance) Hemsl.
分叉露兜Roxb
变叶树参(Champ.) Benth.
白楸(Lam.) Muell. Arg
白背瓜馥木(Hance) Merr.
菝葜
棕竹Burret
锥(Skan) Rehd. et Wils.
浙江润楠
穗花杉(Hance) Pilger
水同木Reinw. ex Bl.
三桠苦
罗浮柿Hance
金毛狗(L.) J. Sm.
黑桫椤Hook.
短叶黍L.
大叶苦柯Chun et Tsiang
扁担藤(Hook.) Gagnep.
百日青D. Don
A comparative study on the community main morphological characteristics of three lower subtropical monsoon evergreen broad-leaved forests
YI Huilin1,2, GUO Yingtao2, LIU Ling2, LIU Xiaodan2, HUANG Liujing1,LIU Shizhong1, ZHANG Qianmei1, LIU Nan1, HAN Ruihong3, REN Hai1,*
1. South China Botanical Garden, Chinese Academy of Sciences, Guangzhou510650, China2.Pubang Landscape Architecture Company Limited, Guangzhou 510600, China3. Zhongkai College of Agricultural Technology, Guangzhou 510225, China
In the paper, we analyzed the community main morphological characteristics such as community life-form spectrum, Raukiaer leaf characteristics, fruit types and the plants ornamental characteristics of the lower subtropical monsoon evergreen broad-leaved forests in Dinghu mountain, Daling mountain and Baiyun mountain, south China. The results indicated that the plants dominated by phanerophytes as well as by leathery and entire mesophylls with higher percentage of simple leaves in these three communities. Due to the different succession time, there were differences in the life-form spectrum; the percentage of Mesophanerophytes was the highest in thecommunity of Dinghu mountain. The Mesophanerophytes and Microphanerophytes were the dominant life-form in thecommunity of Daling mountain, and the percentage of Microphanerophytes was the highest in thecommunityin Baiyun mountain. The ratios of the plants with different leaf characteristics in the three communities were different; the ratio of mesophytes characteristics such as mesophylls, papery leaves at Dinghushan community was higher than the other two communities. Fleshy fruit plants dominated in the three communities, and passive dispersal was the main way. This study can be used for regional urban forest construction
lower subtropical; monsoon evergreen broad-leaved forest;life-form; Raukiaer leaf characteristics;fruit types; ornamental characteristics
10.14108/j.cnki.1008-8873.2017.06.019
Q948.15+8
A
1008-8873(2017)06-137-10
2016-09-16;
2016-10-12
国家基金面上项目“植物功能性状驱动南亚热带季风常绿阔叶林演替的机理研究”(编号31570422)
易慧琳(1991—), 女, 四川资阳人, 研究生,E-mail: YHL0314@yeah.net
任海, 男, 博士, 研究员, 主要从事植被生态恢复研究, E-mail: renhai@scib.ac.cn
