白菜型冬油菜抗寒性研究进展
2017-12-20袁金海顾学平李自峰贾晓莉鲁丽莉马丽娟
袁金海 顾学平 李 涛 李自峰 贾晓莉 鲁丽莉 马丽娟
(甘肃省临泽县种子管理局,临泽 734200)
白菜型冬油菜抗寒性研究进展
袁金海 顾学平 李 涛 李自峰 贾晓莉 鲁丽莉 马丽娟
(甘肃省临泽县种子管理局,临泽 734200)
抗寒性强是白菜型冬油菜能够在-32℃极端低温下安全越冬的一个关键因素。由于北方寒旱区冬寒春旱,积温不够,一般农作物只能一年一熟,土地利用率低。此外,北方地区冬末春初的沙尘暴和扬沙天气使得地区生态环境极其脆弱。因此,白菜型冬油菜抗寒品种的成功选育及推广对提高北方地区生态效益和经济效益具有十分重要的意义。文章综述了白菜型冬油菜抗寒性的最新研究进展,主要包括抗寒性机制研究、提高抗寒性方法及抗寒性育种等情况,以期为从事冬油菜育种工作者提供借鉴。
白菜型冬油菜;抗寒性;抗寒机理
植物抗寒性的形成源于植物长期对寒冷生境条件的适应,是在基因与环境互作的双重条件下引起植物体内多个信号系统参与抗寒应激的综合反应过程。寒冷干燥是我国北方地区冬季典型的气候特征,每年冬末春初的沙尘暴和扬沙天气在北方已司空见惯。随着北方寒旱区冬油菜成功北移[1],陇油系列冬油菜品种为西北这片广袤土地生态环境的改善、轮作制度的革新、经济效益的增加带来了新的希望。十几年来,伴随着分子生物学和生物技术的发展,冬油菜抗寒性机理研究也不断深入,已经在生理生化[2-15]、相关基因克隆[16-24]、相关激素应答[25-30]、蛋白质组学[31-37]等方面取得了一定成果,并在此基础上提出了一系列作为鉴定其抗寒性的综合指标。但是,冬油菜抗寒机理形成是一个复杂的过程,对于明确其抗寒性的响应模式及关键链式反应还需进一步深入研究。随着转录组学的出现和发展,想必会为相关研究提供一个新的思路。本文就冬油菜北移工作开展以来所取得的研究成果作以总结,以期为冬油菜研究工作者提供参考和借鉴。
1 冬油菜抗寒性机制研究
1.1 细胞膜系统、组织含水量及花芽分化与抗寒性 研究发现:在植物逆境胁迫中,细胞膜结构性质的变化先于细胞其他结构活性的变化,主要表现为细胞膜通透性的改变。盖玥等[2]研究结果表明冬油菜随着温度的降低,叶片细胞膜透性显著增加,且强抗寒品种膜透性增幅较小。同时,为了适应低温胁迫,叶片组织含水量也明显下降。张朋飞等[10]也指出,低温胁迫下,冬油菜根部含水量呈“厂”字形变化;而花芽作为植株由营养生长向生殖生长过渡转变的标志,其分化程度可以反映出品种的抗寒性强弱,北方寒旱区冬油菜花芽分化过程研究结果表明,与弱抗寒品种相比较,强抗寒品种花芽表现出分化早、分化速度慢的特点。
1.2 植株生育期及细胞酶系统与抗寒性 孙万仓等[4]认为可以从植物学形态和生长发育进程两方面来判断冬油菜的抗寒性。强抗寒品种植株在越冬前生长缓慢,作为地上部分的叶片进入枯叶期早,根系发达,能够为越冬期贮备大量的有机物,翌年返青时间晚,生长发育周期长。冬油菜北移(如由天水北移到张掖)后,整个生育期呈“短—长—短”的特点,且产量发生较大的变化,抗寒性弱的早熟品种产量下降,而抗寒性强的晚熟品种增产显著[8]。低温条件下,白菜型冬油菜抗氧化酶活性变化与细胞冰冻状态之间的相关性表明[11],强抗寒品种在冷冻后叶片不产生水渍或水渍较少,其根在冻后受冷害程度较轻,弱抗寒品种则相反。蒲媛媛等[9]、刘自刚等[12]对低温胁迫下白菜型冬油菜不同抗寒品种的生理特性研究发现,低温胁迫下,强抗寒和弱抗寒品种油菜叶片SOD和CAT酶活性、可溶性蛋白含量、MDA含量随温度的降低均呈现先升高后下降的变化规律,且抗寒性强的品种能保持较高低温SOD、CAT活性、可溶性蛋白含量和较低的MDA含量。武军艳等[13]研究了冬油菜唯一能够越冬的根部器官,发现根部相关的酶活性及渗透调节物质与抗寒性存在相关性,降温后,各测量指标与越冬率相关性显著或极显著,而降温前,相关性则不显著。此外,史鹏辉等[14]研究发现,可以利用同工酶谱带对冬油菜抗寒性进行鉴定分类,谱带数越多且谱带越宽的品种抗寒性越强。
1.3 光合特性与抗寒性 光合速率作为衡量植物生长发育状况的重要指标,能够直接反映植物受逆境胁迫的程度。植物光合器官对低温反应灵敏,刘自刚等[12]研究夜间低温对白菜型冬油菜光合机构的影响结果表明,夜间温度降至5℃处理7d后,不同抗寒品种气孔导度、胞间CO2浓度、净光合速率(Pn)均下降,且弱抗寒品种叶片气孔大多数处于半开半闭的状态,而强抗寒品种气孔仍然保持着完全开放的状态,其光合速率、根冠比、根部干物质积累均显著高于弱抗寒品种。研究发现,叶片栅栏组织与海绵组织比例,海绵组织厚度,叶片厚度,叶片上、下表皮厚度,叶片气孔的密度及大小与植物抗寒性密切相关。许耀照等[15]对冬油菜叶片解剖结构研究指出白菜型冬油菜在冬前低温条件下无光合午休现象,且Pn变化表现为“单峰”曲线,同时强抗寒品种叶片组织结构(气孔密度、面积、周长等)变幅较小,光抑制程度较强。低温条件下,气孔对大气湿度和光照反应不敏感,导致植物气孔密度变小,长度变短,有利于植物在低温胁迫下维持较高的组织含水量,提高水分利用率。
1.4 植物激素与抗寒性 生产上常采用控制植物生长调节剂(植物激素)来提高作物的抗寒性,通过借鉴前人的研究经验,利用植物激素进行抗寒性的研究在冬油菜生产上得到了进一步应用。方彦等[25]冬油菜浸种试验表明,30mg/L浓度的ABA是冬油菜抗寒性最佳诱导剂。张腾国等[38]认为MAPKK抑制剂能够通过控制冬油菜光合作用,改变相关酶的表达活性提高植株抗寒能力。刘海卿等[28]通过冬油菜不同叶龄期喷施一定浓度的外源ABA研究发现,一定浓度的ABA能够明显提高冬油菜的越冬率,且在6叶期喷施条件下抗寒性提高最为显著,进一步丰富了武军艳等[26]的前期研究结果。随着研究的深入,最终确定了冬油菜叶片最佳的外源ABA喷施浓度为20mg/L[27]。陈奇等[30]在自然降温过程中测定了冬油菜6个温度(25℃、12℃、8℃、3℃、-1℃、-4℃)梯度下内源激素ABA、GA3、IAA、ZR含量及其比值的变化,试验结果表明:较高的ABA含量和较低的GA3、IAA、ZR含量能够促进冬油菜抗寒性的形成,进而提高其抗寒能力。
1.5 抗寒相关基因与抗寒性 曾秀存等[16-18]研究发现,白菜型冬油菜的抗寒性的增强,主要受APX基因、Fe-SOD基因和Cu/Zn-SOD基因的调控表达。半定量RT-PCR及酶活性分析表明,APX基因、Fe-SOD基因和Cu/Zn-SOD基因的表达量伴随着低温程度的逐级加剧,均呈“上升—下降—上升”的趋势,与实际生产中田间冬油菜的生长习性相吻合,说明APX基因、Fe-SOD基因和Cu/Zn-SOD基因属于低温诱导表达的抗逆基因。张腾国等[19-22]利用RACE技术从陇油6号中分别克隆了BnICE1、BnMKK4、CBF、COR、BnMKK2、BnMPK6 和MKK2基因,实时荧光定量表明BnICE1的转录受到低温胁迫诱导,在油菜抵御寒冷等环境胁迫的过程中发挥作用。马骊等[23]研究发现白菜型冬油菜BnCOR25、BnICE1和Cu/Zn-SOD基因在低温下被诱导表达,其相对表达量随温度的降低均上升,且均在-4℃时到达峰值。后来本课题组利用双向电泳对分离出的类甜蛋白进行了基因克隆和表达分析[24],其结果与曾秀存等[16-18]抗寒性分析试验结果相同。
1.6 抗寒相关蛋白与抗寒性 当植物遭受不良环境的胁迫时,作为植物体的第一道天然屏障——质外体,为了响应逆境胁迫,从而激发质外体信号系统的表达,以此来抵御相关的外来胁迫。袁金海等[33]对冬油菜质外体蛋白的提取方法进行了探究,在保证高提取率和低污染率的情况下明确了适合冬油菜质外体蛋白的提取方法,为进一步研究抗寒相关蛋白奠定了基础。
抗冻蛋白(AFPs)是一类具有抑制胞内冰晶形成且对已经形成的冰晶具有抑制其体积继续增大的抗冻活性物质。1992年,M.Griffith等[39]第一次从经低温锻炼的冬黑麦叶片质外体中得到并部分纯化了AFPs,开启了植物抗冻蛋白的研究。受前人研究的启发,史鹏辉[31]从冬油菜陇油6号的叶片及根部提取了质外体蛋白,电泳结果表明在33~45kD之间出现了一条特异性条带,经质谱分析鉴定为β-1,3-葡聚糖酶,该酶在冬黑麦中已被证明为抗冻蛋白,具有冰晶修饰活性。杨刚等[32]对经过冷驯化的陇油6号叶片和根部质外体蛋白提取液进行活性分析发现,冷驯化后其提取液具有极强的冰晶抑制活性。刘自刚等[34]比较了-20℃条件贮存的经过低温处理的陇油7号叶片质外体蛋白提取液冰晶形态,再次证明和丰富了课题组先前的研究结果,向更加精准地明确冬油菜的抗寒机理迈进了可喜的一步。
目前,双向电泳技术作为蛋白质组学研究的首选技术,在研究作物抗逆性差异蛋白质的分析及鉴定中越来越受到研究者的青睐。陈奇等[35]在蒲媛媛[36]的研究基础上首次对适合于冬油菜双向电泳技术进行了优化,并且分离出了白菜型冬油菜陇油7号响应低温胁迫的关键蛋白——类甜蛋白。袁金海等[37]模拟我国北方初春显著的倒春寒现象,利用双向电泳技术成功分离出了双半胱氨酸型氧化还原蛋白酶过氧化物酶(2-Cysperoxiredoxin),为进一步明确冬油菜响应反复低温胁迫与恢复的气候条件机理提供了理论基础。
通过以上研究结果可以发现,白菜型冬油菜响应抗寒机制模式如下(图1)。
2 冬油菜抗寒性提高方法与措施
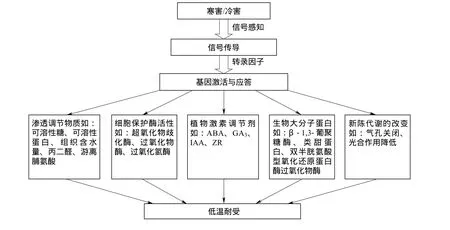
图1 白菜型冬油菜响应抗寒机制模式
目前,随着一系列超强抗寒品种的成功选育,以陇油6号、陇油7号为典型代表的品种得到大范围的应用和推广,研究表明,可以通过采用覆盖处理、改变施肥方式、调整播期、控制群体密度、激素浸种、化学诱变剂处理、叶面喷施激素等提高冬油菜的越冬率和抗寒性。孙万仓等[3]研究发现,甘肃中部、北部露地栽培的冬油菜越冬率为77%,通过不同覆盖方式处理,其越冬率较露地提高1.3%~19.0%,且覆盖效果表现为地膜覆盖>秸秆覆盖>农家肥+土覆盖>农家肥覆盖,说明覆盖处理对冬油菜具有良好的保墒防冻作用。曾军等[5]研究结果表明,科学合理的施肥方式能够最大限度的保证冬油菜安全越冬,冬前施种肥、冬前追肥以及冬前施种肥和冬前追肥的施肥方式对冬油菜的生长发育有着显著的影响,通过不同的施肥方式处理,冬油菜的越冬率可提高2~3个百分点。春化对冬油菜花芽分化有着显著的影响,这主要表现在适宜播期的确定,燕妮等[6]认为,适期早播可提高冬油菜的越冬率,主要原因为不同播期导致冬油菜的苗龄不同,进而影响其渗透调节物质和细胞保护性酶活性等生理生化特性的变化程度不同。叶剑等[7]指出,合理的种植密度能够提高冬油菜的壮苗率,有利于其安全越冬,在种植密度为60万株/hm2的情况下越冬率为89%,当种植密度扩大1倍(120万株/hm2)时,其越冬率反而降为80%,由此可见,合理的种植密度也是提高冬油菜越冬率的关键技术之一。方彦等[25]研究了不同浓度ABA浸种对冬油菜抗寒性的诱导效应,试验结果表明冬油菜抗寒性的提高主要借助于ABA能诱导细胞保护酶活性的变化,进而通过酶系统的连锁反应来减轻低温对细胞的伤害程度。化学诱变剂在提高作物抗逆性方面已得到了大量的应用,课题组发现,一定体积分数的EMS诱变剂能提高细胞膜保护酶活性,酶活性越高消除氧自由基的能力越强,从而提高冬油菜对低温的耐受能力[40]。武军艳等[26-27]、刘海卿等[28-29]对利用叶面喷施激素提高冬油菜抗寒性技术进行了多年研究,表明外源ABA能够显著提高细胞保护酶活性和渗透调节物质等生理生化物质含量,缓解零下低温对冬油菜的冻害;郭仁迪等[41]在此基础上,研制出了适宜于提高冬油菜抗寒性的新型抗寒复合剂(20mg/L ABA+300mg/L PEG-6000+50mg/L PP333),其对冬油菜越冬率较对照提高18.87个百分点,产量和经济系数增加显著。
3 抗寒性育种成就
选择更寒冷的生态环境和抗寒性强的基因型进行轮回选择是改良北方冬油菜抗寒性的有效途径[42],目前,生产上应用和推广的抗寒丰产良种陇油系列品种都是通过轮回选择育成的。植物的抗寒性是由多基因控制下的数量性状不断累积并逐渐提高,孙万仓科研团队成功培育的陇油6号、陇油7号等陇油系列超强抗寒品种可耐-32℃,其抗寒性已达到国际领先水平。陇油6号等抗寒品种的选育,把我国冬油菜种植范围由35°N向北推至40°N~48°N,向北推移了5~13个纬度,打破了业内人士关于我国原有冬春油菜产区呈似“厂”型的分界线,对我国油菜产区进行了重新区划。杨建胜等[40]研究了经EMS处理的陇油7号种子,其生育期各项生理指标表明EMS能够提高冬油菜的抗寒性。抗寒早熟品种陇油9号的选育更大程度地满足了我国北方一年多熟的耕作制度。今天,在国家“一带一路”伟大战略背景下,在以“生态文明”为时代主题的号召下,冬油菜种植和推广将会带来更加显著的生态效益和经济效益。
[1] 孙万仓,马卫国,雷建明,等.冬油菜在西北旱寒区的适应性和北移的可行性研究 [J].中国农业科学,2007,40(12):2716-2726
[2] 盖玥,牛俊义,孙万仓,等.降温处理对白菜型油菜品种抗寒生理指标的影响 [J].甘肃农业大学学报,2005,40(2):182-185
[3] 孙万仓,牛俊义,滕文惠,等.覆盖处理对旱寒区冬油菜越冬率和产量的影响 [J].中国油料作物学报,2006,28(3):315-318
[4] 孙万仓,武军艳,曾军,等.8个白菜型冬油菜品种抗寒性的初步评价 [J].湖南农业大学学报,2007,33(43):151-155
[5] 曾军,孙万仓,张亚宏,等.不同施氮方式对冬油菜生理生化指标及生长发育和产量的影响[J].西北农业学报,2008,17(3):176-181
[6] 燕妮,孙万仓,朱惠霞,等.播期对冬油菜越冬率及生理生化特性的影响 [J].中国农学通报,2007,23(7):250-253
[7] 叶剑,孙万仓,武军艳,等.群体密度对冬油菜产量和经济性状的影响 [J].西北农业学报,2008,17(3):171-175
[8] 孙万仓,武军艳,方彦,等.北方旱寒区北移冬油菜生长发育特性[J].作物学报,2010,36(12):2124-2134
[9] 蒲媛媛,孙万仓.白菜型冬油菜抗寒性与生理生化特性关系[J].分子植物育种,2010,8(2):335-339
[10] 张朋飞,孙万仓,武军艳,等.北方寒旱区冬油菜花芽分化过程观察 [J].西北农业学报,2011,20(11):60-65
[11] 史鹏辉,孙万仓,赵彩霞,等.低温下抗氧化酶活性与冬油菜根细胞结冰关系的初步研究[J].西北植物学报,2013,33(2):329-335
[12] 刘自刚,孙万仓,杨宁宁,等.冬前低温胁迫下白菜型冬油菜抗寒性的形态及生理特征 [J].中国农业科学,2013,46(22):4679-4687
[13] 武军艳,方彦,张鹏飞,等.北方旱寒区冬油菜根系抗寒指标分析[J].干旱地区农业研究,2014,6(32):250-255
[14] 史鹏辉,刘自刚,张亚宏,等.23份油菜的抗氧化酶特性及过氧化物同工酶分析 [J].西北农业学报,2014,23(1):113-119
[15] 许耀照,曾秀存,张芬琴,等.白菜型冬油菜叶片结构和光合特性对冬前低温的响应 [J].作物学报,2017,43(3):432-441
[16] 曾秀存,孙万仓,方彦,等.白菜型冬油菜抗坏血酸过氧化物酶(APX)基因的克隆、表达及其活性分析[J].作物学报,2013,39(8):1400-1408
[17] 曾秀存,刘自刚,史鹏辉,等.白菜型冬油菜铜锌超氧化物歧化酶(Cu/Zn-SOD)基因的克隆及其在低温条件下的表达[J].作物学报,2014,40(4):636-643
[18] 曾秀存,孙万仓,孙佳,等.白菜型冬油菜铁超氧化物歧化酶(Fe-SOD)基因的克隆及表达分析 [J].中国农业科学,2013,46(21):4603-4611
[19] 张腾国,常燕,王娟,等.油菜BnICE1的克隆及表达分析[J].中国农业科学,2013,46(1):205-214
[20] 张腾国,聂婷婷,陈琼琼,等.白菜型油菜CBF基因克隆及功能分析 [J].西北植物学报,2015,35(10):1964-1971
[21] 张腾国,郭艳峰,陈琼琼,等.油菜COR基因的克隆及原核表达[J].甘肃农业科技,2015(4):1-4
[22] 张腾国,郭艳峰,王圆圆,等.油菜MKK2基因原核表达及特性分析 [J].兰州大学学报,2015,51(3):405-410
[23] 马骊,孙万仓,刘自刚,等.低温胁迫下白菜型与甘蓝型冬油菜抗寒基因表达差异[J].中国油料作物学报,2016,38(3):135-141
[24] 马骊,袁金海,孙万仓,等.白菜型冬油菜类甜蛋白的筛选、克隆及其在低温胁迫下的表达[J].作物学报,2017,43(4):623-631
[25] 方彦,武军艳,孙万仓,等.外源ABA浸种对冬油菜种子萌发及幼苗抗寒性的诱导效应 [J].干旱地区农业研究,2014,32(6):70-74
[26] 武军艳,刘海卿,方彦,等.外源ABA对白菜型冬油菜‘陇油8号’抗寒性的影响 [J].核农学报,2015,29(9):1828-1832
[27] 武军艳,刘海卿,孙万仓,等.苗期喷施外源脱落酸对北方白菜型冬油菜越冬生理的影响[J].中国油料作物学报,2015,37(3):310-315
[28] 刘海卿,武军艳,孙万仓,等.不同叶龄叶面喷施ABA对北方白菜型冬油菜抗寒性的影响[J].草业学报,2015,24(9):173-180
[29] 刘海卿,方园,武军艳,等.低温胁迫下内源ABA、GA及比值对白菜型和甘蓝型冬油菜抗寒性的响应[J].中国生态农业学报,2016,24(11):1529-1538
[30] 陈奇,袁金海,孙万仓,等.低温胁迫下白菜型冬油菜与春油菜叶片光合特性及内源激素变化比较[J].中国油料作物学报,2017,39(1):37-46
[31] 史鹏辉.北方强冬性白菜型冬油菜抗寒机制研究[D].兰州:甘肃农业大学,2012
[32] 杨刚,史鹏辉,孙万仓,等.白菜型冬油菜质外体抗冻蛋白研究[J].中国生态农业学报,2016,24(2):210-217
[33] 袁金海,刘自刚,孙万仓,等.白菜型冬油菜叶片质外体蛋白提取及响应低温胁迫的蛋白表达分析[J].华北农学报,2016,31(4):44-50
[34] 刘自刚,袁金海,孙万仓,等.低温胁迫下白菜型冬油菜差异蛋白质组学及光合特性分析[J].作物学报,2016,42(10):1541-1550
[35] 陈奇,袁金海,刘自刚,等.白菜型冬油菜‘陇油7号’叶片响应低温胁迫的差异蛋白鉴定与分析[J].中国生态农业学报,2017,25(3):381-390
[36] 蒲媛媛.低温胁迫下白菜型冬油菜抗寒性的生理特性与蛋白质组的双向电泳分析[D].兰州:甘肃农业大学,2010
[37] 袁金海,刘自刚,孙万仓,等.白菜型冬油菜反复低温与恢复过程中叶片的差异蛋白分析 [J].分子植物育种,2017,15(1):307-314
[38] 张腾国,张艳,夏小慧,等.MAPKK抑制剂对低温胁迫下油菜幼苗光合作用和抗氧化酶活性的影响[J].兰州大学学报,2013,49(1):92-98
[39] Griffith M,Ala P,Yang D S,et al.Antifreeze protein produced endogenously in winter rye leaves[J].Plant Physiol,1992,100(2):593-596
[40] 杨建胜,孙万仓,刘自刚,等.化学诱变剂EMS对白菜型冬油菜‘陇油7号’形态及生理生化的影响[J].西北农业学报,2015,24(3):75-81
[41] 郭仁迪,刘海卿,武军艳,等.抗寒复合剂对白菜型冬油菜生长发育及产量性状的影响[J].干旱地区农业研究,2017,35(4):263-269
[42] 孙万仓,曾秀存,刘自刚,等.轮回选择对白菜型冬油菜抗寒性及经济性状的影响[J].中国油料作物学报,2015,37(4):443-452
张掖国家级玉米种子生产基地(临泽)项目
2017-09-13)
